1. Introduction
Recent developments in computational modeling of the heart’s bio-electrical activity have established the most highly detailed example of a virtual organ [
1,
2,
3]. These advances are the result of the significant progress in cardiac cell modeling, corroborated with experimentation and clinical practice. Moreover, continuous increases in computational power have led to whole-heart electrical models that have shown promising translational outcomes, improving the general understanding of heart function, its pathologies, and therapies [
4,
5,
6,
7,
8,
9]. Still, critical modeling challenges arise when local heterogeneities at different spatio-temporal scales are taken into account [
10,
11,
12,
13,
14,
15,
16].
Various approaches have been proposed to simulate complex electrical behavior of the heart, e.g., ventricular arrhythmias. First, detailed cell models, with highly accurate and validated biophysical relationships representing the ground truth, have been incorporated to improve the physiological relevance of in silico cardiac predictions. Nonetheless, these approaches may result computationally demanding, further requiring advanced optimization tools [
17,
18,
19,
20,
21,
22,
23]. Alternatively, reduced phenomenological models were developed, preserving the main features of physiological approaches but still allowing for a robust in silico investigation. Depending on the application at hand, one of the two options is favorable, identifying the specific characters that minimize alterations of the final results [
24]. This is especially true in the study of abnormal electrical waves, e.g., ventricular tachycardia (VT) and ventricular fibrillation (VF), in healthy and diseased conditions [
25,
26].
The computational cardiology literature contains a plethora of biophysical models that describe the electrical behavior of the human myocardium [
27]. A detailed description of ion channels, pumps, and exchangers is usually included to explain experimental observations [
28]. However, many parameters are highly difficult or nearly impossible to measure in vivo, and different formulations have been shown to replicate similar dynamics. For example, the O’hara Rudy dynamic (Ord) model (41 variables, 16 parameters) and the Ten Tusscher-Panfilov 2006 (TP06) model (19 variables, 48 parameters) produce similar electrical behaviors [
29,
30,
31]. In addition, the multiple time scales involved in the formulation, from milliseconds to seconds, make the resulting dynamic system very stiff, which poses challenges in the numerical solution [
32,
33,
34]. Alternatively, phenomenological models are derived from the overall description of cumulative inward and outward currents across the cell membrane and rely on a smaller set of variables. For instance, the Mitchell-Schaeffer model (2 variables, 9 parameters) [
35] and its generalizations [
36] or the Fenton-Karma (FK) model (3 variables, 14 parameters) and its later extensions (minimal model—4 variables, 28 parameters) [
37,
38] are well-known instances of simplified phenomenological descriptions able to reproduce several dynamic properties of the cardiac electrical activity: threshold of excitation, maximum upstroke velocity, action potential (AP) shape and morphology, restitution properties, action potential duration (APD) and alternans dynamics [
39,
40,
41]. Increased numerical efficiency is enforced, lacking detailed ionic descriptions, although well-suited for large-scale whole-heart numerical studies.
Cardiac arrhythmias, such as VT and VF, have been studied in detail using physiologically-based computational approaches. Various cardiac diseases, such as long-QT syndrome or Brugada Syndrome, have been elucidated using advanced computational techniques [
42,
43,
44]. Computational cardiology modeling has also been used to explain the relation between AP shape changes with the likelihood of a reentrant arrhythmia (see e.g., [
24] and references therein). Estimates of membrane potential and ionic currents distributions in the core region of a sinoatrial node reentry was obtained through detailed computer simulations [
45]. Subject-specific whole-heart computational models explored arrhythmia risk-stratification in patients with myocardial infarction (MI) [
9,
46], and complex VF dynamics have also been characterized in terms of scroll wave filaments through detailed ventricular models [
42]. Likewise, the study of AP vortex dynamics has been conducted via phenomenological descriptions in both simplified [
47,
48,
49] and whole-heart simulations [
50,
51,
52,
53]. These studies show that the in silico study of arrhythmias is currently being done either using phenomenological ionic models or biophysical ionic models, but how the choice of ionic model affects the arrhythmic signatures of VF in normal and failing hearts remains understudied. Furthermore, heart simulations based on phenomenological ionic models are expected to take less wall-clock time due to the simplified mathematical representation of simplified ionic models. This has been explored in previous works using the Eikonal equation to simulate action potential propagation [
40]. However, using the complete monodomain or bidomain approach, a study of the differences in computing time for biophysical versus phenomenological ionic models in realistic heart geometries has not been fully elucidated.
The scientific question that guides the current study is the following: How does the choice of ionic model affect the dispersion of repolarization, arrrhythmic signatures of VF, and the overall computing time? To answer this question, we constructed subject-specific computational models of normal and failing hearts using two models of ionic transmembrane current, namely the FK and the TP06 models. To analyze the differences between these two modeling choices, we assessed the resulting dispersion of repolarization, VF signatures, and wall-clock time.
4. Discussion
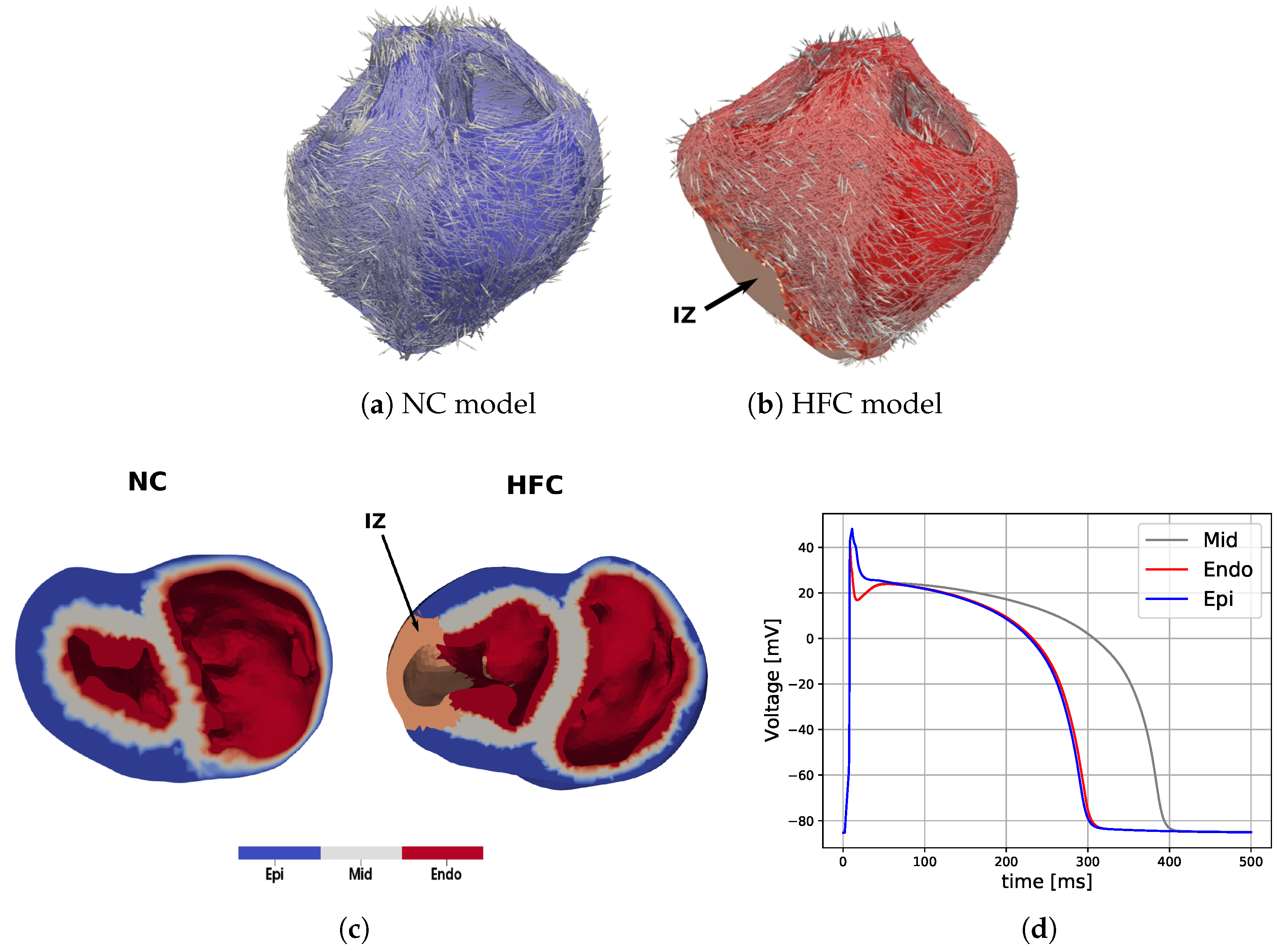
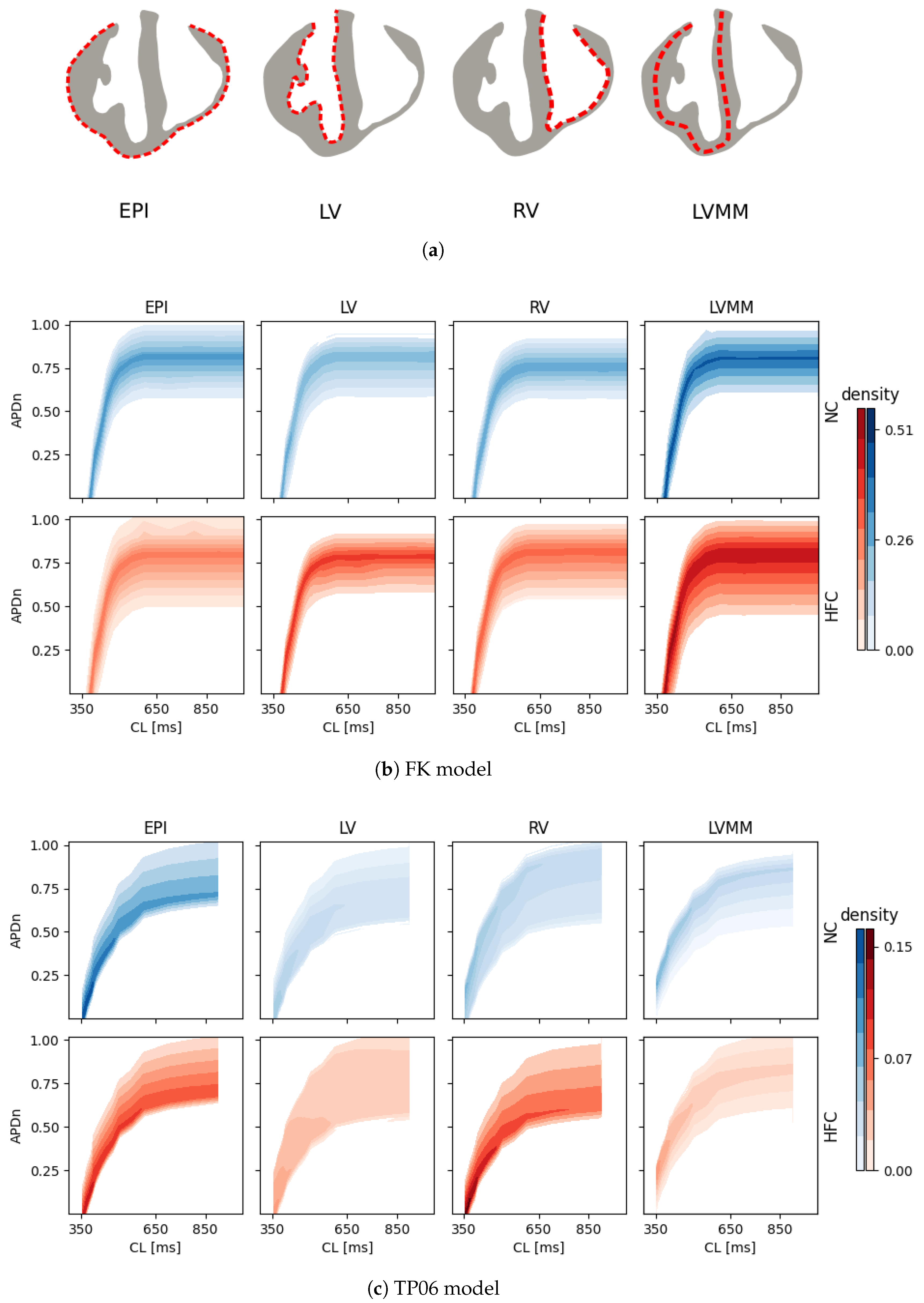
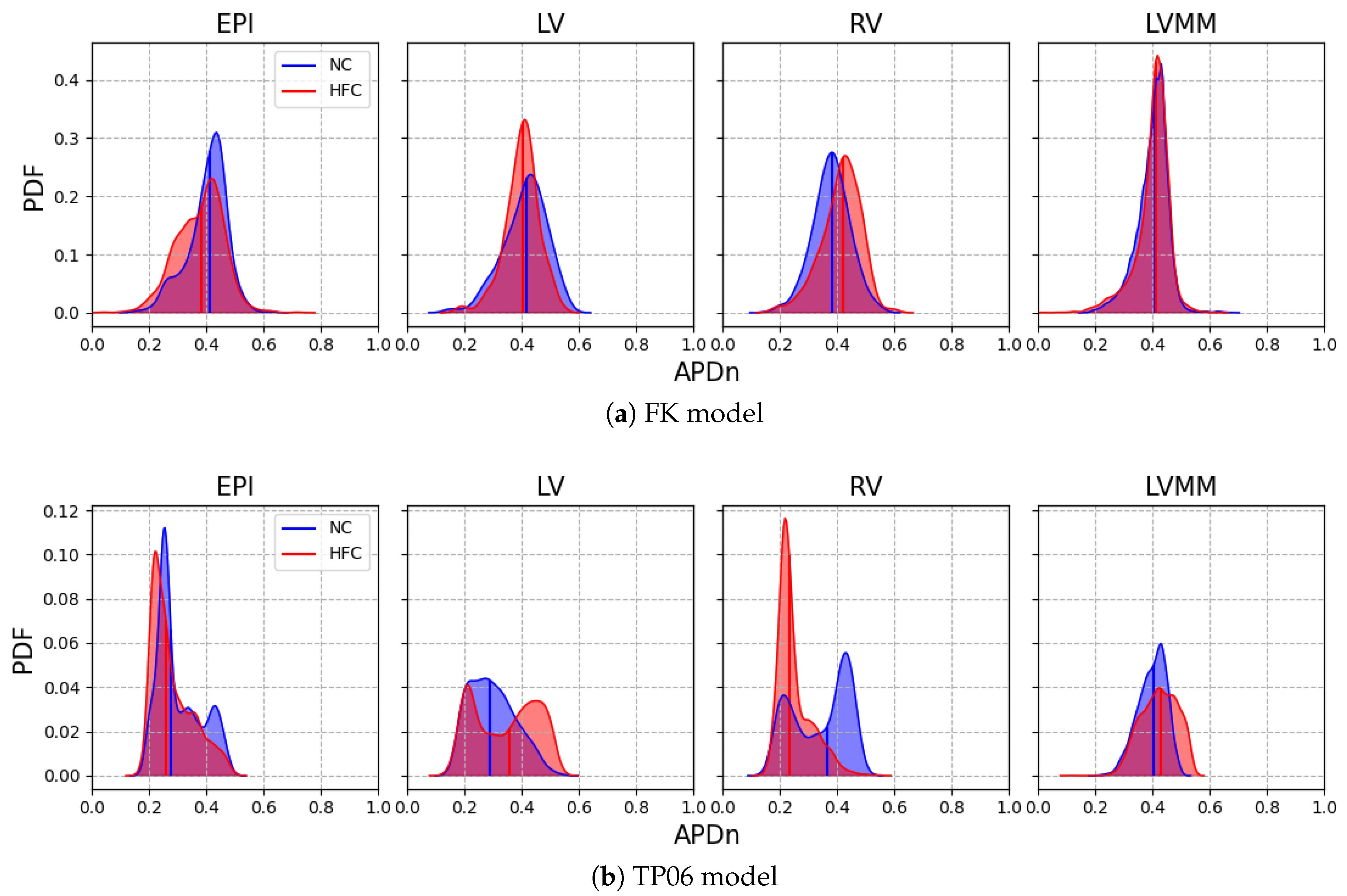
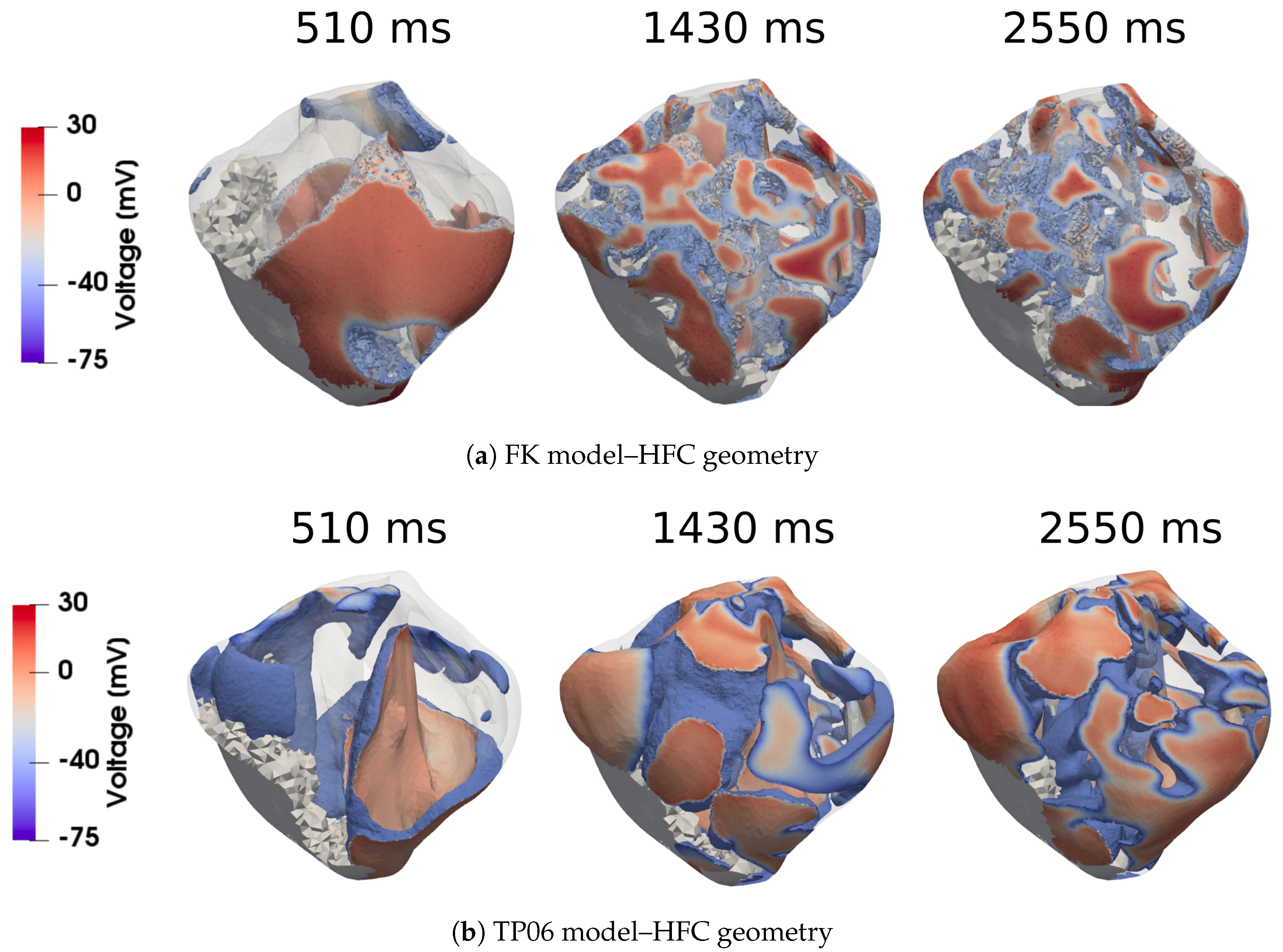
In this work, we investigated how the choice of ionic models affects the repolarization properties and VF signatures of computational models of normal and failing hearts. Numerical analyses allowed us to build up probability distribution functions of APD restitution curves for four selected anatomical surfaces. Accordingly, we identified left and right ventricular endocardium as the anatomical regions characterized by a higher degree of dispersion of repolarization (multimodal PDFs) for the myocardial infarction case. On the other hand, strong electrotonic effects within the mid-myocardial layer homogenize the resulting electrical behavior, thus providing coherent PDFs among the two ionic models for healthy and diseased cases, although non-negligible differences appear for epicardium, left and right endocardium.
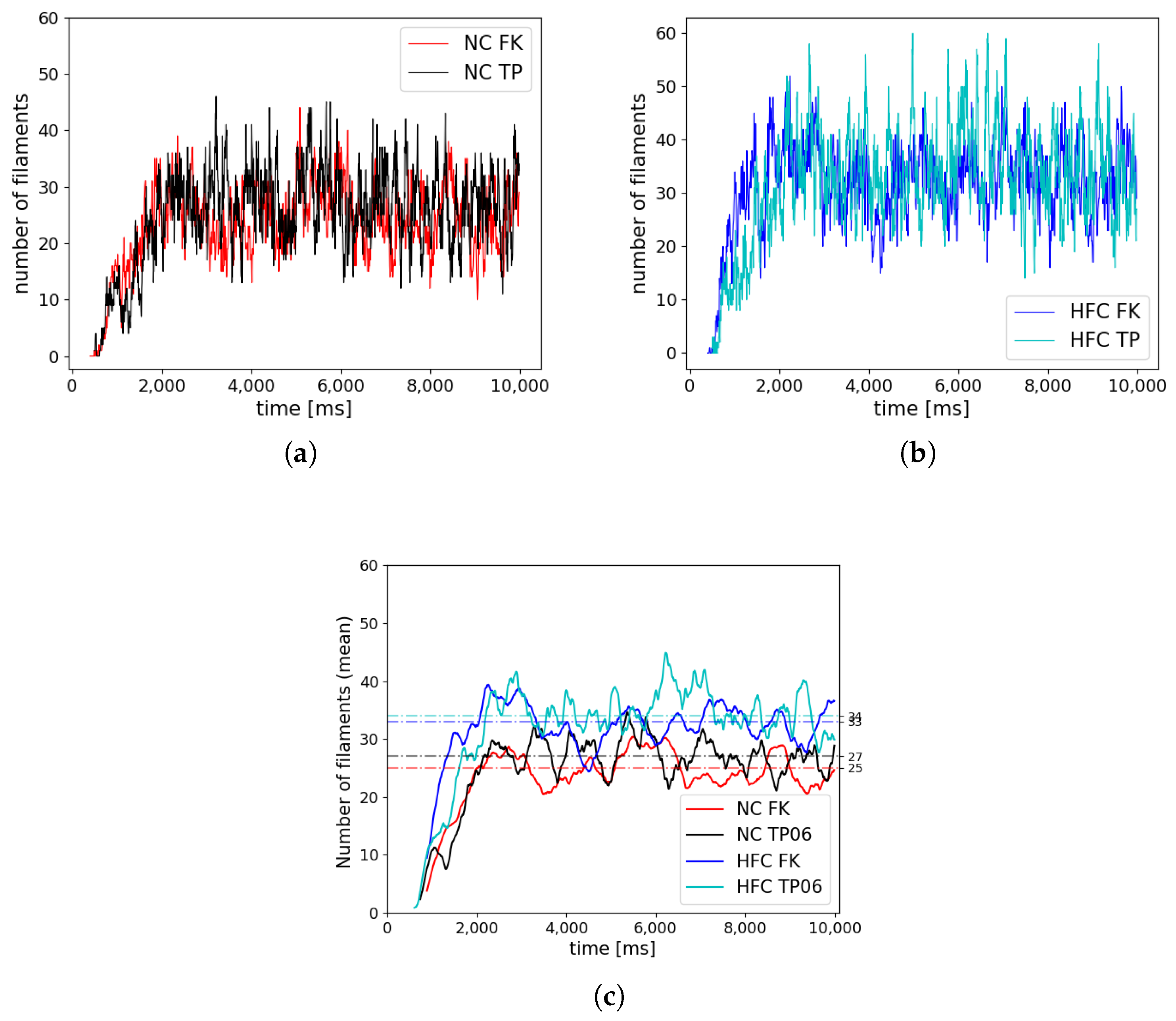
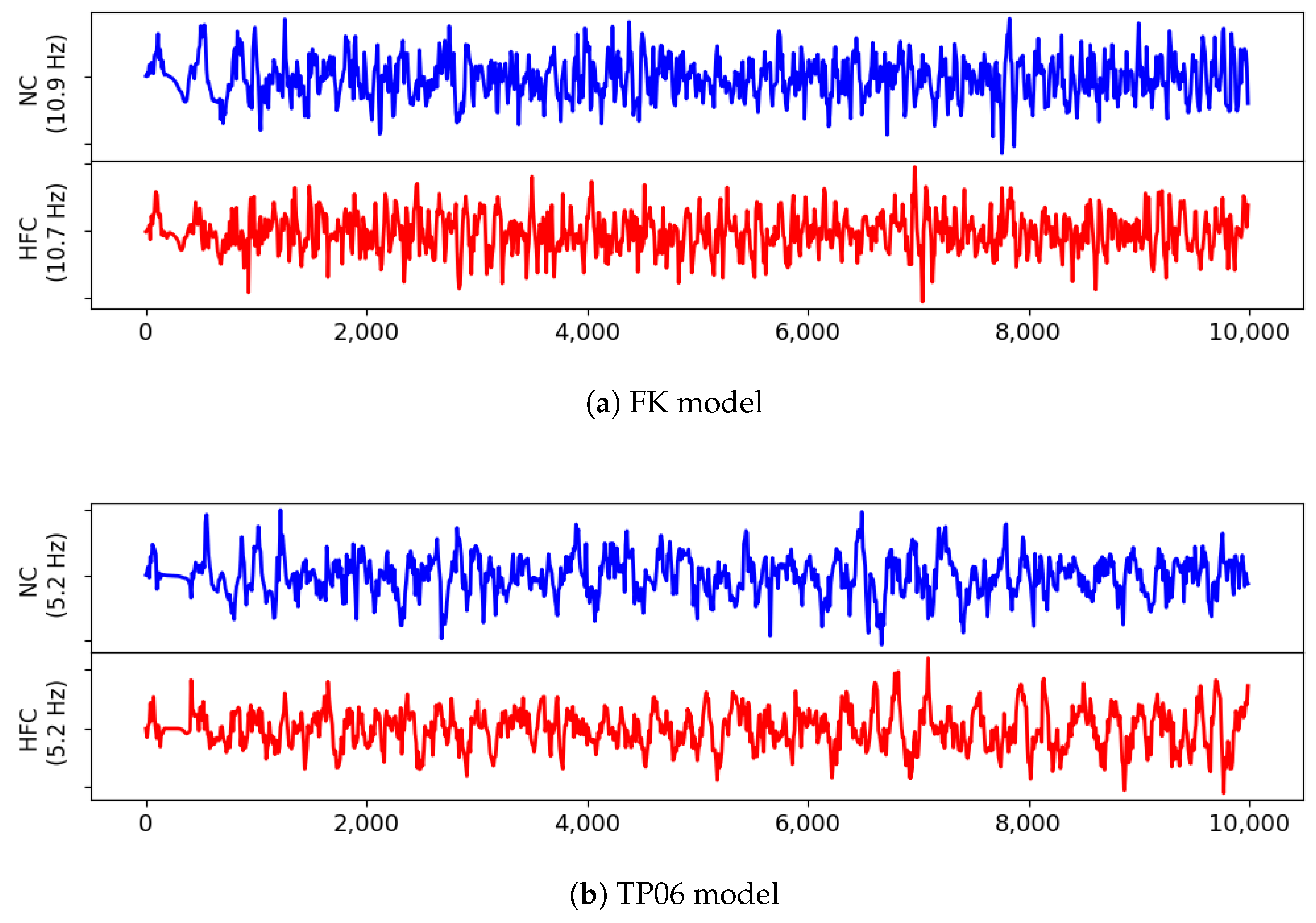
This result, consistent with the literature and specific kinetics described by the TP06 ionic model, was further confirmed by long-run simulations conducted for ventricular fibrillation. The statistical examination of the evolution and the number of vortex filaments provided an enhanced arrhythmogenicity in the diseased case. Constitutive differences among the two electrophysiological models appear in terms of restitution dynamics at large cycle lengths, stimulus intensity, and vulnerable windows. Such a discrepancy is due to the intrinsic complexity of the ionic models in delivering an external stimulation current signal among the different gating variables. The membrane capacitance and the surface-to-volume coefficients regulate the electrical current’s spreading, with a predominant role in ionic modeling cases. Therefore, we highlight the necessity of fine-tuning these parameters when using a physiological approach other than a phenomenological one. However, dissimalirities were negligible for ventricular fibrillation signature. Specifically, irrespective of the healthy or pathological geometry model, the fundamental frequency of the pseudo-ECG during sustained VF is constant. Moreover, the number of filaments in the HFC case was statistically equal among FK and TP06 models.
Accordingly, our results suggest that VF signatures are mainly controlled by anatomical and structural features rather than regional restitution properties. Therefore, given the large-scale computational models and clinical translation, the choice of a simplified phenomenological description seems favorable. From a computational point of view, phenomenological models allow the implementation of highly efficient and simple integration schemes, whereas biophysical-based models, such as the TP06, require specialized and adequate time integration schemes that demand a complex implementation. Several studies have tackled the development of time-integration strategies to cope with the stiff nature of these models [
34,
67]. Besides the high computational demand for solving the electrophysiology equations, the used programming language (
Python) is limited by its lack of performance and scalability. Moreover, more efficient numerical approaches, like spatio-temporal adaptivity or strong scalable solvers, are also needed to cope with the computational burden imposed by high-resolution electrophysiological problems. Future research may use a programming language better suited in terms of scalability and performance, such as C++, which may be used in conjunction with
Python and highly advanced numerical techniques [
60,
68]. Despite this, we solved a highly computational demanding problem in a relatively modest amount of time. Moreover, several parameters required for postprocessing (i.e., pseudo-ECG, APD distribution, and filaments) were computed during simulation time, saving time for analysis.
The current work can be extended in several directions. First, a general comparison between phenomenological and biophysical models requires the investigation of the bidomain theoretical framework to emphasize the role of structural material heterogeneities during defibrillation procedures [
69,
70,
71]. In this direction, a proper mathematical and computational modeling of torso will be critical for obtaining clinically relevant ECG signals, i.e., acceleration of forward ECG numerical modeling with the use of personalized torso geometries [
72]. Second, comparing linear, nonlinear, and fractional diffusion formulations would significantly help to identify the best computational approach to apply in subject-specific cardiac studies [
10,
73,
74,
75]. Third, characterization of the border zone of the scar and gray zones in the infarcted myocardium is related to small-scale components [
9,
76] that are accurately modeled by detailed biophysical constitutive models reproducing modified ion channel dynamics (electrical remodeling). However, reduced-order models have been shown to provide a viable alternative also in this context [
77]. This is key in clinical-related applications such as drug delivery and stem cell delivery in which disease characterization and treatment depend on the electrochemical dynamics of the cell membrane and the associated local heterogeneities.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






