
Analysis of Polyphenol Patterns of Pleurotus ostreatus Cultivars by UHPLC-ESI-MS/MS; Application of FT-NIR and Chemometric Methods, Classification Options




Abstract
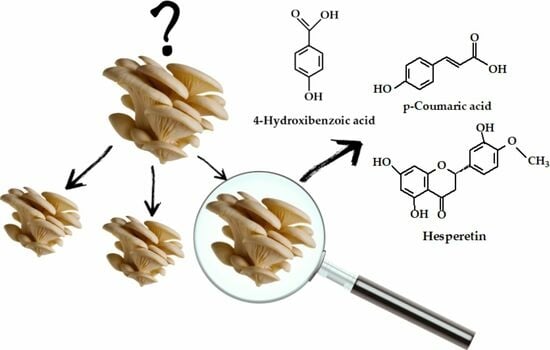
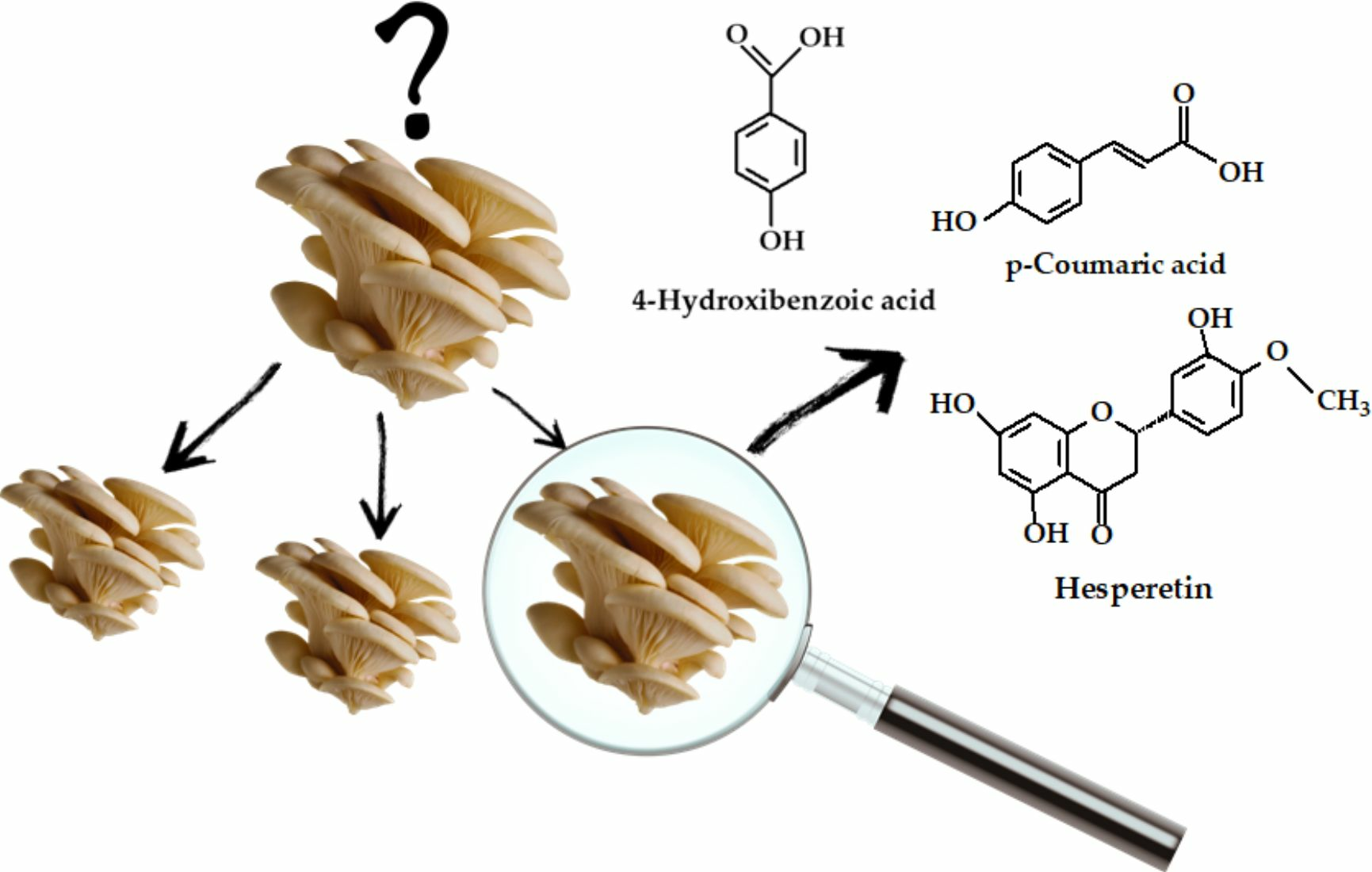
:
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Chemicals and Reagents
2.3. Polyphenol Determination by UHPLC-ESI-MS/MS
2.4. Total Polyphenol Content (TPC)
2.5. Antioxidant Capacity (FRAP)
2.6. Free Amino Acid Content (fAA)
2.7. Fourier-Transform near Infrared Spectroscopy (FT-NIR)
2.8. Statistical Evaluation
2.8.1. Normality Test
2.8.2. Correlation Analysis
2.8.3. Nonparametric Tests for Difference—Kruskal–Wallis
2.8.4. Principal Component Analysis (PCA)
2.8.5. Software Applications
3. Results
3.1. Polyphenol Composition
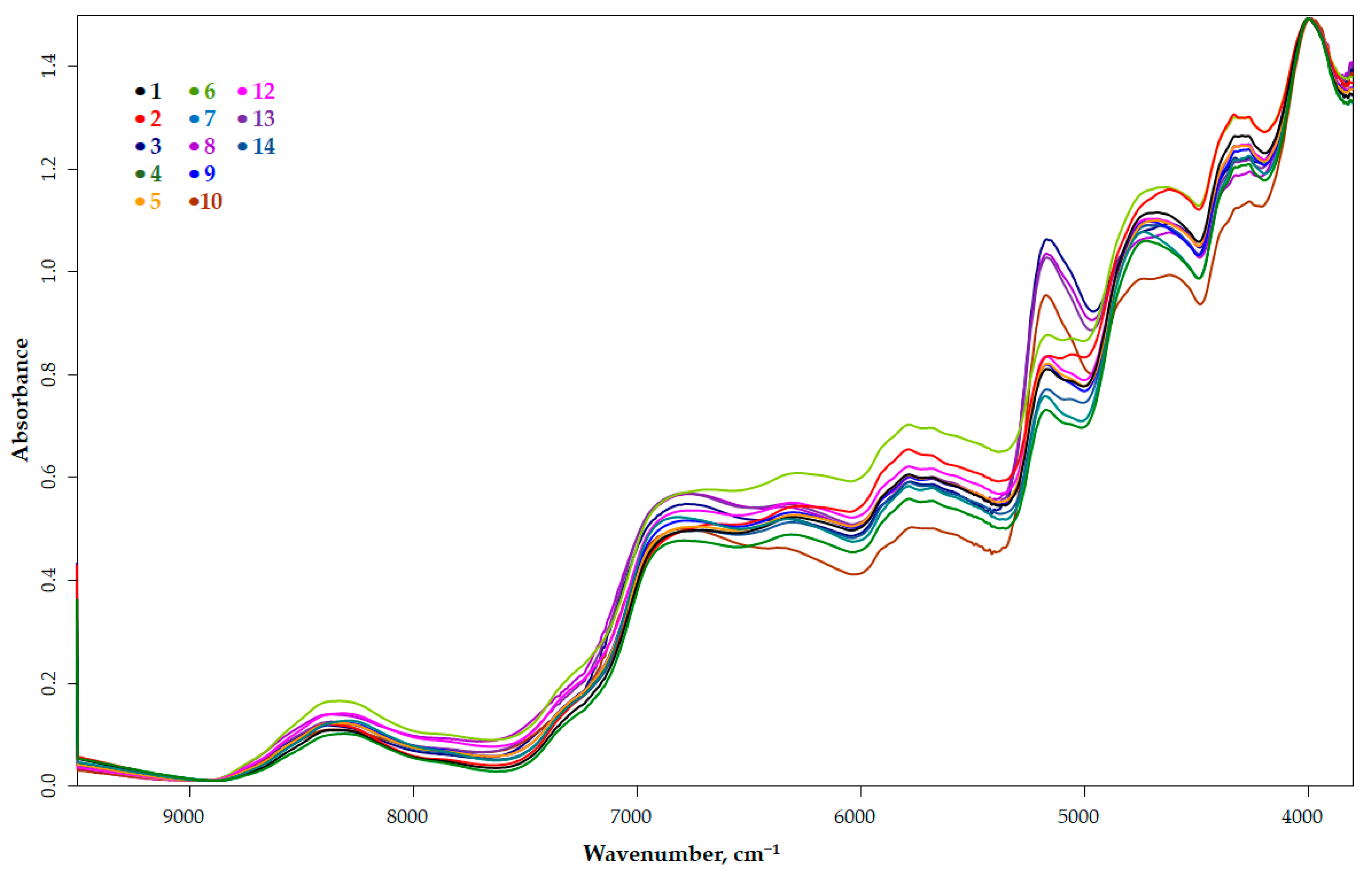
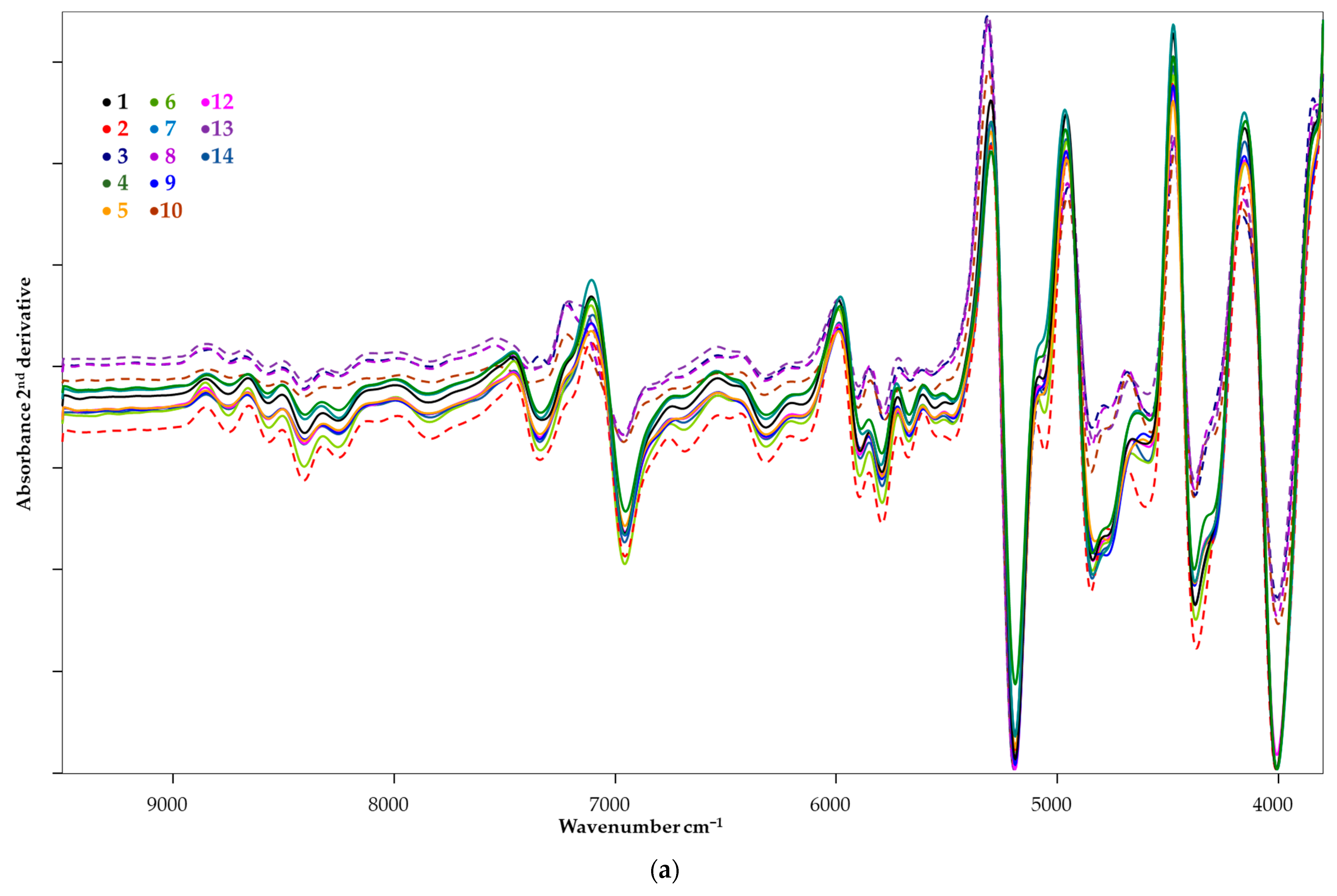
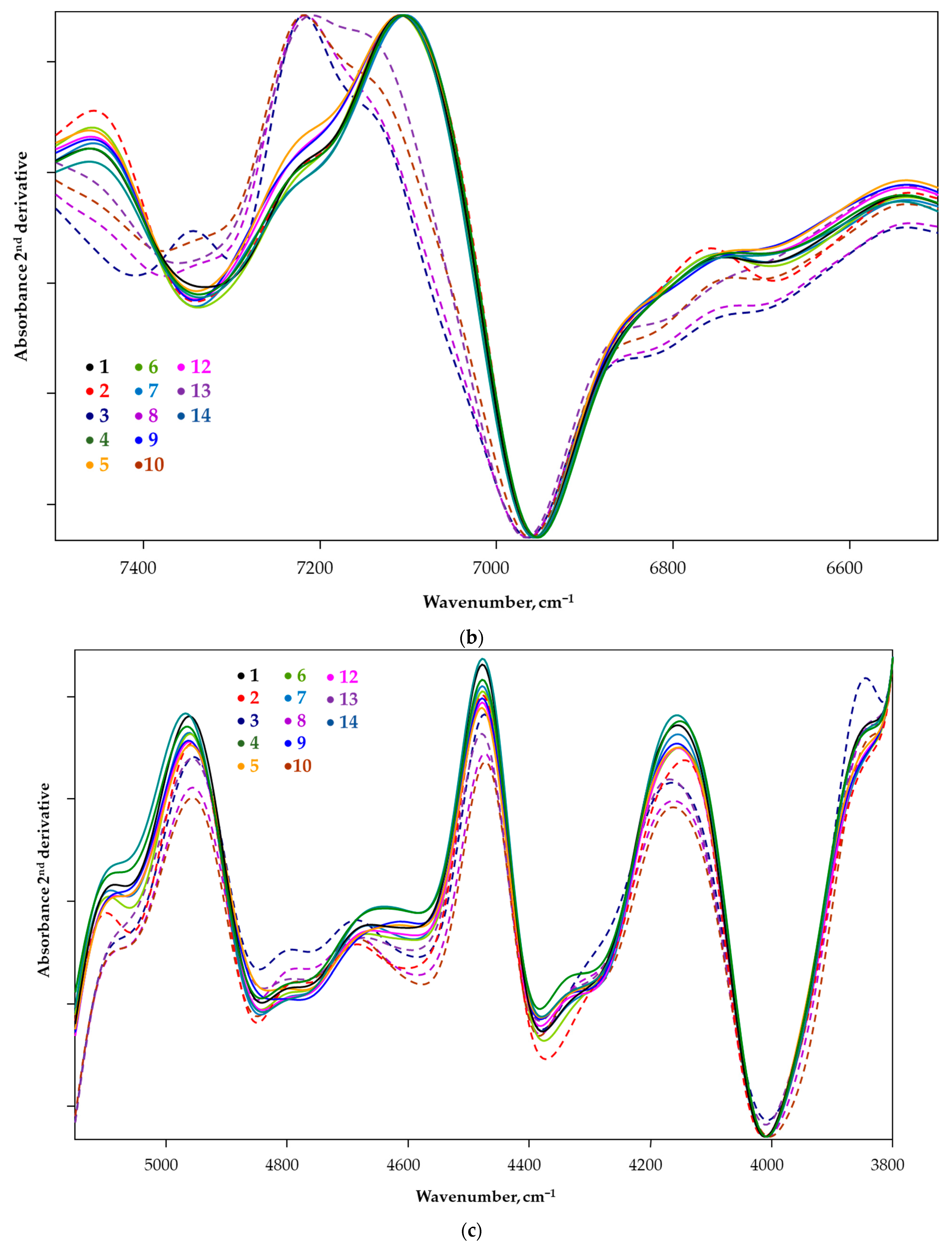
3.2. Spectrum Profiling
3.3. Chemometric Evaluation of Polyphenol Data
3.3.1. Normality Testing
3.3.2. Correlation Analysis
3.3.3. Kruskal–Wallis Test
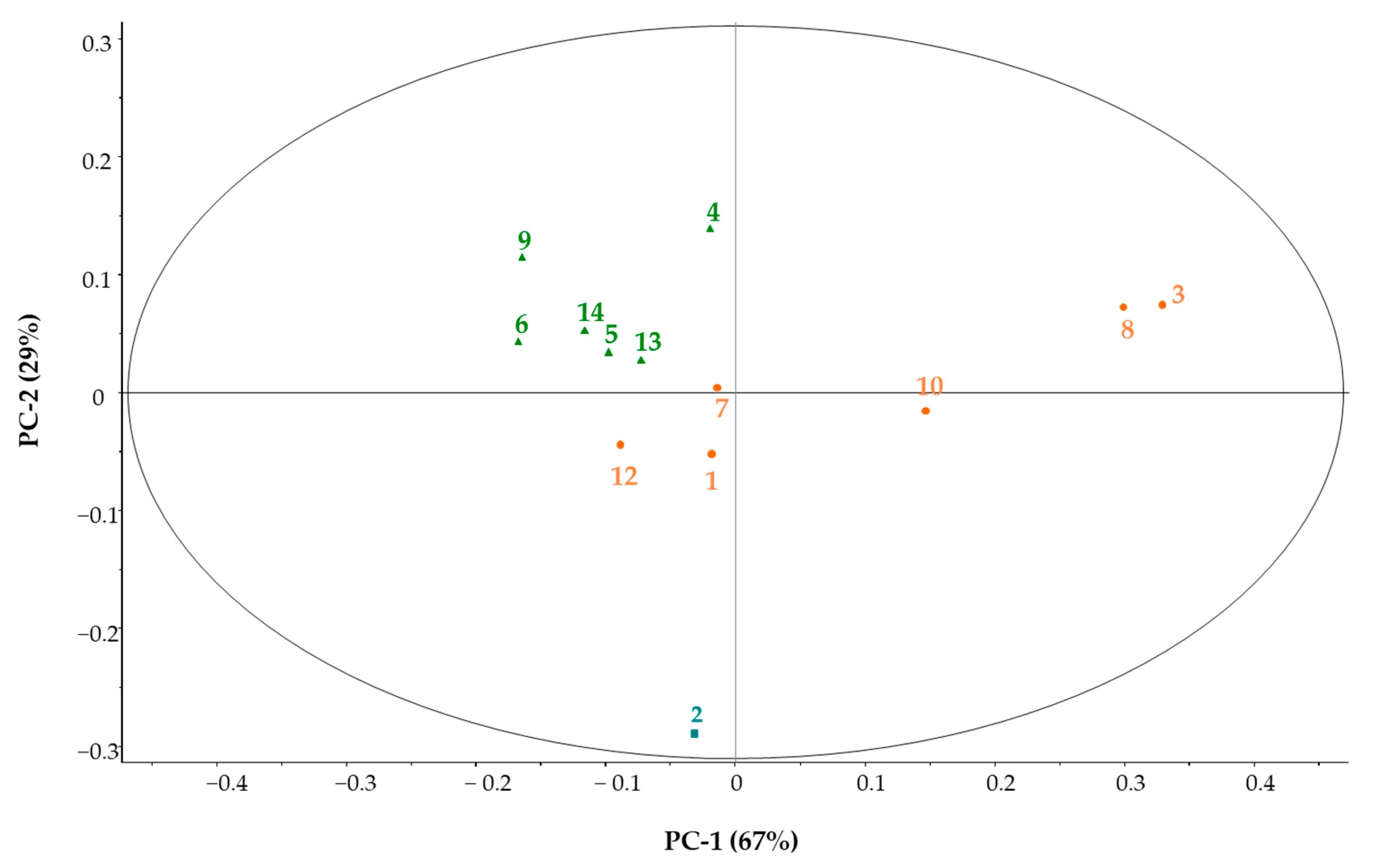
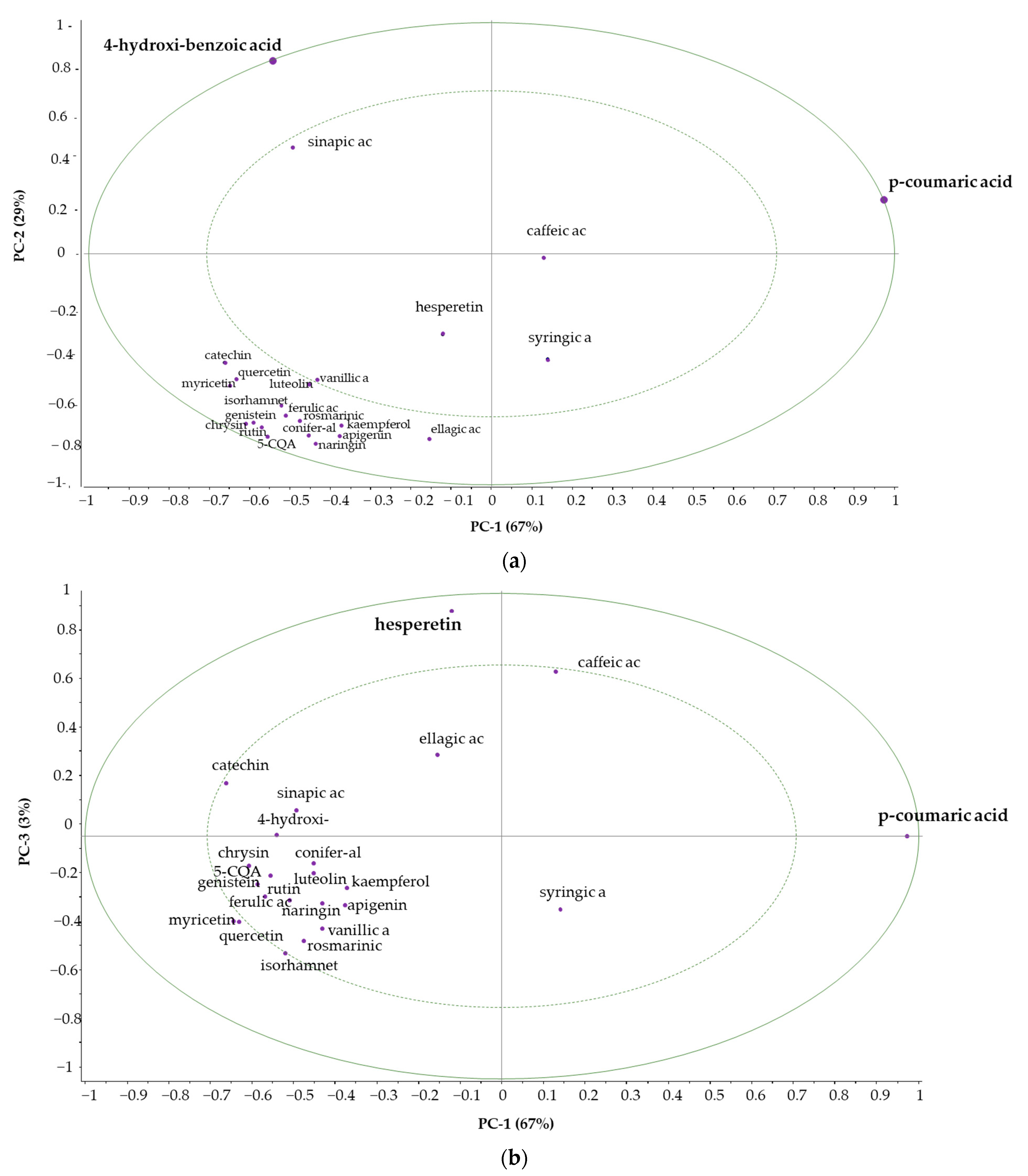
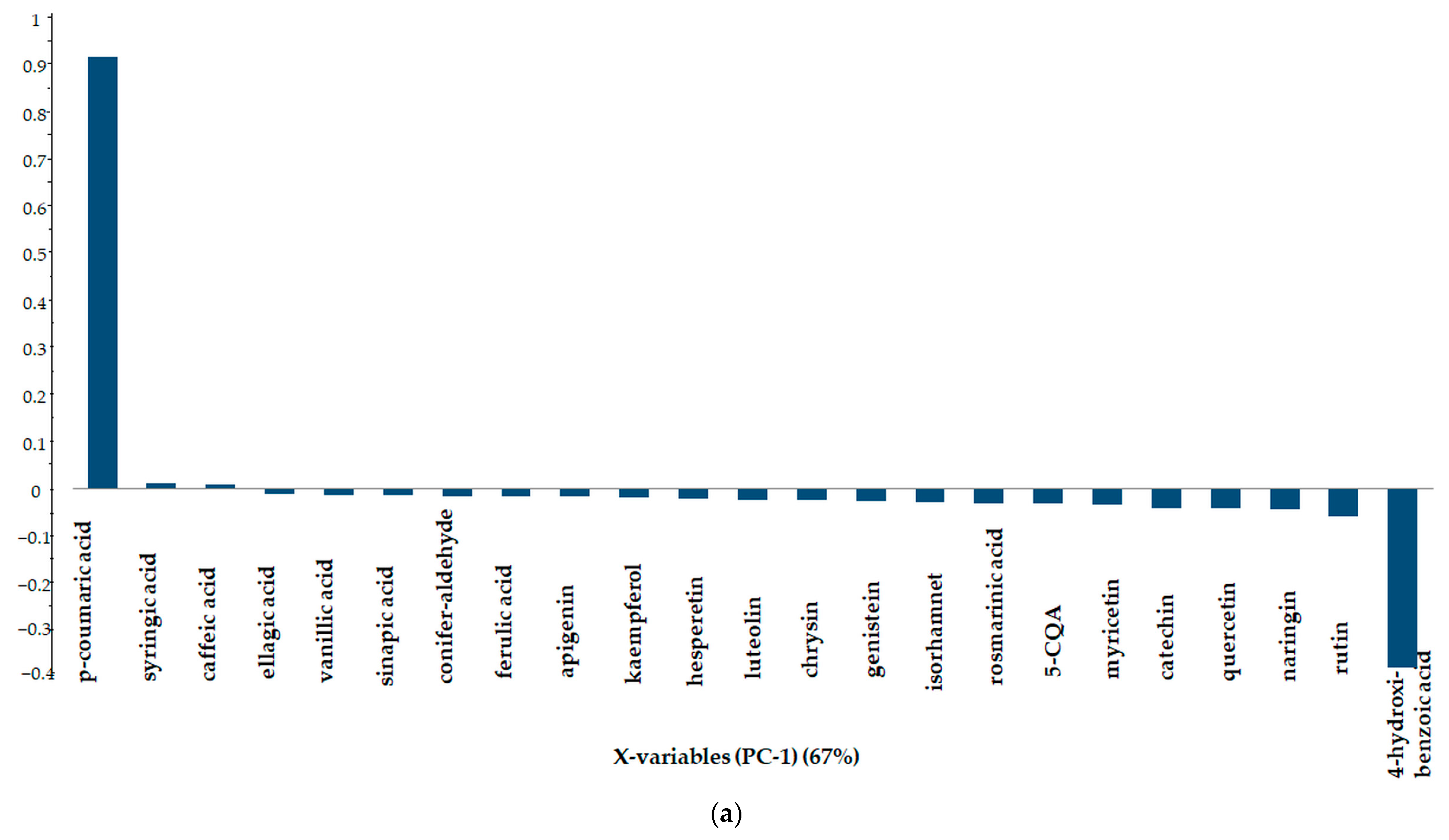
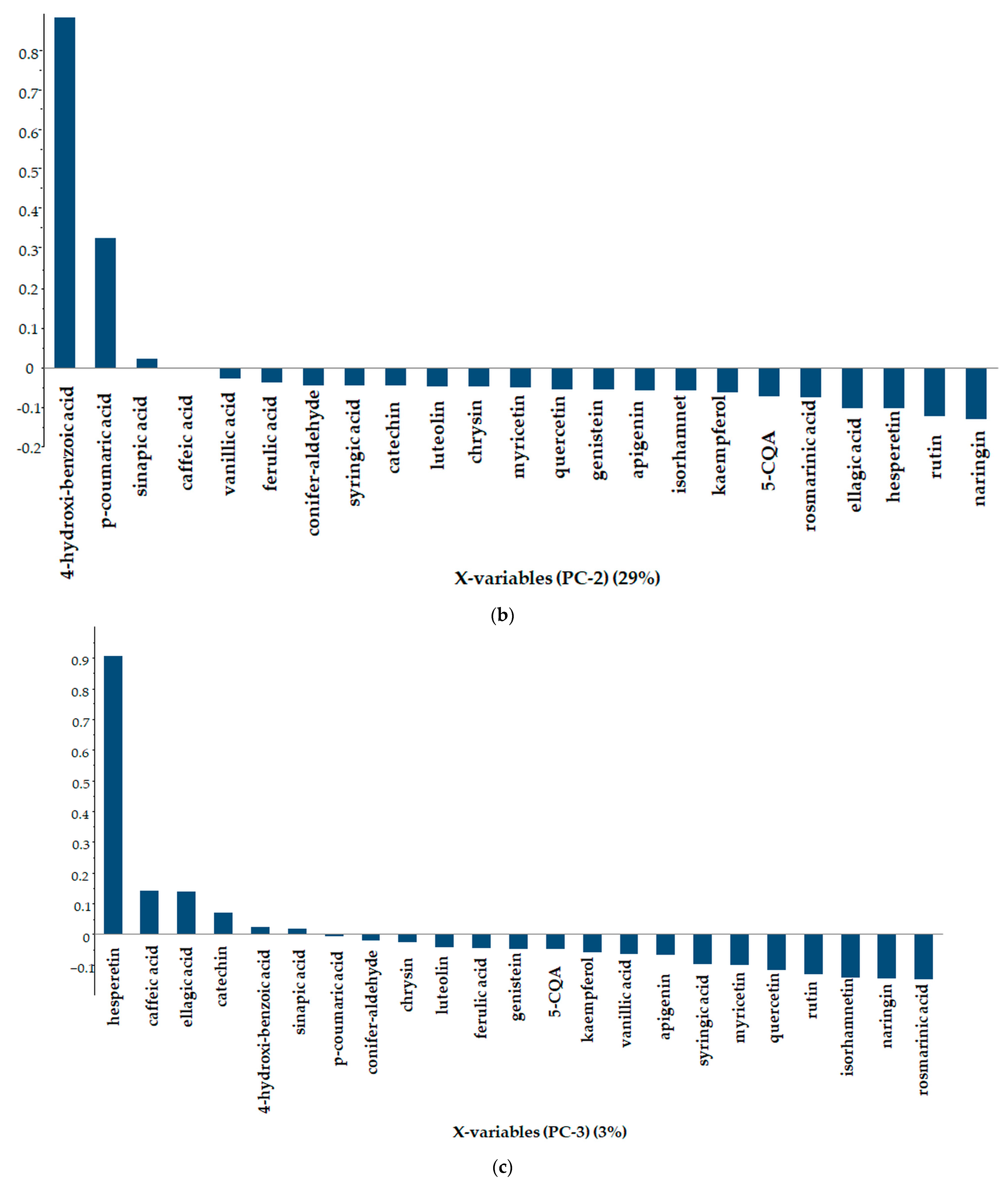
3.3.4. Principal Component Analysis
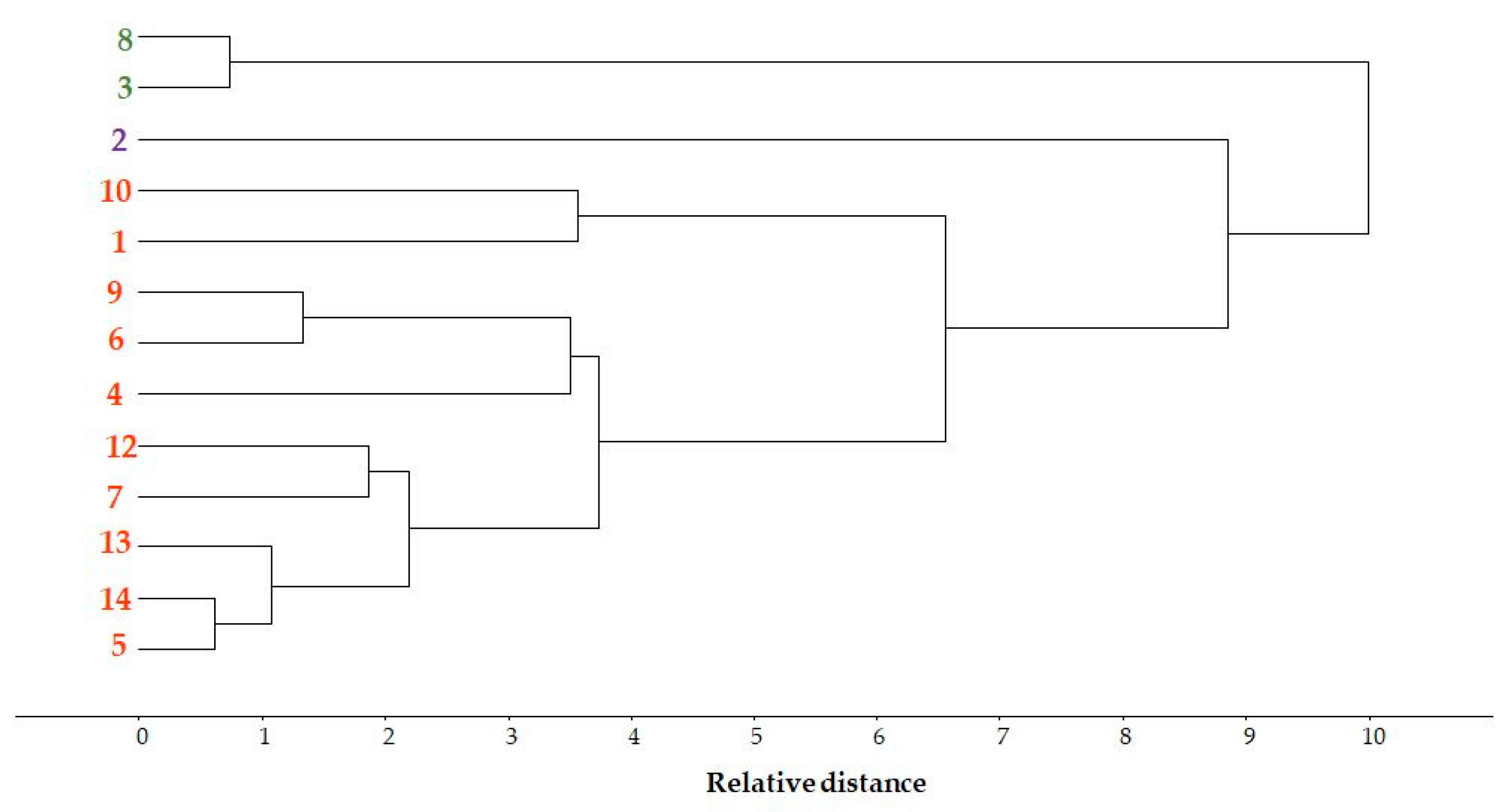
3.3.5. Pattern Recognition by Cluster Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wan Mahari, W.A.; Peng, W.; Nam, W.L.; Yang, H.; Lee, X.Y.; Lee, Y.K.; Liew, R.K.; Ma, N.L.; Mohammad, A.; Sonne, C.; et al. A Review on Valorization of Oyster Mushroom and Waste Generated in the Mushroom Cultivation Industry. J. Hazard. Mater. 2020, 400, 123156. [Google Scholar] [CrossRef]
- Bellettini, M.B.; Fiorda, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Júnior, A.M.; Ribani, R.H. Factors Affecting Mushroom Pleurotus Spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef]
- Muswati, C.; Simango, K.; Tapfumaneyi, L.; Mutetwa, M.; Ngezimana, W. The Effects of Different Substrate Combinations on Growth and Yield of Oyster Mushroom (Pleurotus ostreatus). Int. J. Agron. 2021, 2021, 9962285. [Google Scholar] [CrossRef]
- Ucar, T.M.; Karadag, A. The Effects of Vacuum and Freeze-Drying on the Physicochemical Properties and in Vitro Digestibility of Phenolics in Oyster Mushroom (Pleurotus ostreatus). Food Meas. 2019, 13, 2298–2309. [Google Scholar] [CrossRef]
- Assemie, A.; Abaya, G. The Effect of Edible Mushroom on Health and Their Biochemistry. Int. J. Microbiol. 2022, 2022, 8744788. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, H.; Sun, Y.; Xia, R.; Hou, Z.; Li, Y.; Wang, Y.; Pan, S.; Li, L.; Zhao, C.; et al. Effect of Light on Quality of Preharvest and Postharvest Edible Mushrooms and Its Action Mechanism: A Review. Trends Food Sci. Technol. 2023, 139, 104119. [Google Scholar] [CrossRef]
- Rangsinth, P.; Sharika, R.; Pattarachotanant, N.; Duangjan, C.; Wongwan, C.; Sillapachaiyaporn, C.; Nilkhet, S.; Wongsirojkul, N.; Prasansuklab, A.; Tencomnao, T.; et al. Potential Beneficial Effects and Pharmacological Properties of Ergosterol, a Common Bioactive Compound in Edible Mushrooms. Foods 2023, 12, 2529. [Google Scholar] [CrossRef]
- Plant Metabolites under Environmental Stress: Mechanisms, Responses, and Adaptation Strategies, 1st ed.; Desai, N.M.; Patil, M.; Pawar, U.R. (Eds.) Apple Academic Press: Palm Bay, FL, USA, 2023; ISBN 978-1-77491-094-8. [Google Scholar]
- Abdelshafy, A.M.; Belwal, T.; Liang, Z.; Wang, L.; Li, D.; Luo, Z.; Li, L. A Comprehensive Review on Phenolic Compounds from Edible Mushrooms: Occurrence, Biological Activity, Application and Future Prospective. Crit. Rev. Food Sci. Nutr. 2022, 62, 6204–6224. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.S.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Chemical Composition and Nutritional Value of the Most Widely Appreciated Cultivated Mushrooms: An Inter-Species Comparative Study. Food Chem. Toxicol. 2012, 50, 191–197. [Google Scholar] [CrossRef]
- Lesa, K.N.; Khandaker, M.U.; Mohammad Rashed Iqbal, F.; Sharma, R.; Islam, F.; Mitra, S.; Emran, T.B. Nutritional Value, Medicinal Importance, and Health-Promoting Effects of Dietary Mushroom (Pleurotus ostreatus). J. Food Qual. 2022, 2022, 2454180. [Google Scholar] [CrossRef]
- Domokos-Szabolcsy, É.; Alshaal, T.; Koroknai, J.; Kovács, S.; Tóth, C.; Csilléry, G.; Jókai, Z.; Matkovits, A.; Makleit, P.; Veres, S.; et al. Phytochemical Evaluation of the Fruits and Green Biomass of Determinate-Type Sweet Pepper (Capsicum annuum L. fasciculatum) Grown in Terrestrial Bioregenerative Life-Support Research Facilities. J. Plant Interact. 2023, 18, 2268118. [Google Scholar] [CrossRef]
- Matkovits, A.; Nagy, K.; Fodor, M.; Jókai, Z. Analysis of Polyphenolic Components of Hungarian Acacia (Robinia pseudoacacia) Honey; Method Development, Statistical Evaluation. J. Food Compos. Anal. 2023, 120, 105336. [Google Scholar] [CrossRef]
- Farkas, Á.; Horváth, G.; Kuzma, M.; Mayer, M.; Kocsis, M. Phenolic Compounds in Hungarian Acacia, Linden, Milkweed and Goldenrod Honeys. Curr. Res. Food Sci. 2023, 6, 100526. [Google Scholar] [CrossRef]
- Rahimah, S.B.; Djunaedi, D.D.; Soeroto, A.Y.; Bisri, T. The The Phytochemical Screening, Total Phenolic Contents and Antioxidant Activities in Vitro of White Oyster Mushroom (Pleurotus ostreatus) Preparations. Open Access Maced. J. Med. Sci. 2019, 7, 2404–2412. [Google Scholar] [CrossRef]
- Istasse, T.; Jacquet, N.; Berchem, T.; Haubruge, E.; Nguyen, B.K.; Richel, A. Extraction of Honey Polyphenols: Method Development and Evidence of Cis Isomerization Ubertas Academica. Anal. Chem. Insights 2016, 11, S39739. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Kalogeropoulos, N.; Stathopoulos, P.; Kaliora, A.C.; Zervakis, G.I. Bioactive Compounds and Antioxidant Activity Exhibit High Intraspecific Variability in Pleurotus ostreatus Mushrooms and Correlate Well with Cultivation Performance Parameters. World J. Microbiol. Biotechnol. 2017, 33, 98. [Google Scholar] [CrossRef] [PubMed]
- Lettera, V.; Del Vecchio, C.; Piscitelli, A.; Sannia, G. Low Impact Strategies to Improve Ligninolytic Enzyme Production in Filamentous Fungi: The Case of Laccase in Pleurotus ostreatus. Comptes Rendus Biol. 2011, 334, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Barh, A.; Sharma, V.P.; Annepu, S.K.; Kamal, S.; Sharma, S.; Bhatt, P. Genetic Improvement in Pleurotus (Oyster Mushroom): A Review. 3 Biotech 2019, 9, 322. [Google Scholar] [CrossRef]
- Amirvaresi, A.; Parastar, H. Miniaturized NIR Spectroscopy and Chemometrics: A Smart Combination to Solve Food Authentication Challenges. Front. Anal. Sci. 2023, 3, 1118590. [Google Scholar] [CrossRef]
- Fodor, M.; Mikola, E.E.; Geösel, A.; Stefanovits-Bányai, É.; Mednyánszky, Z. Application of Near-Infrared Spectroscopy to Investigate Some Endogenic Properties of Pleurotus ostreatus. Cultiv. Sens. 2020, 20, 6632. [Google Scholar] [CrossRef] [PubMed]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant Properties of Phenolic Compounds Occurring in Edible Mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. [2] Ferric Reducing/Antioxidant Power Assay: Direct Measure of Total Antioxidant Activity of Biological Fluids and Modified Version for Simultaneous Measurement of Total Antioxidant Power and Ascorbic Acid Concentration. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 15–27. ISBN 978-0-12-182200-2. [Google Scholar]
- Sheard, J. Quantitative Data Analysis. In Research Methods; Elsevier: Amsterdam, The Netherlands, 2018; pp. 429–452. ISBN 978-0-08-102220-7. [Google Scholar]
- Ostertagová, E.; Ostertag, O.; Kováč, J. Methodology and Application of the Kruskal–Wallis Test. Appl. Mech. Mater. 2014, 611, 115–120. [Google Scholar] [CrossRef]
- Ringnér, M. What Is Principal Component Analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Seguin, P.; Ahn, J.-K.; Kim, J.-J.; Chun, S.-C.; Kim, E.-H.; Seo, S.-H.; Kang, E.-Y.; Kim, S.-L.; Park, Y.-J.; et al. Phenolic Compound Concentration and Antioxidant Activities of Edible and Medicinal Mushrooms from Korea. J. Agric. Food Chem. 2008, 56, 7265–7270. [Google Scholar] [CrossRef]
- Workman, J.; Weyer, L. Practical Guide to Interpretive Near-Infrared Spectroscopy; CRC: Boca Raton, FL, USA, 2008; ISBN 978-1-57444-784-2. [Google Scholar]
- Beć, K.B.; Huck, C.W. Breakthrough Potential in Near-Infrared Spectroscopy: Spectra Simulation. A Review of Recent Developments. Front. Chem. 2019, 7, 48. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Polyphenol Concentration of Cultivars (mg/kg) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | 5th | 6th | 7th | 8th | 9th | 10th | 12th | 13th | 14th | |
| 4-Hydroxibenzoic acid | 10.62 ± 0.81 | 98.23 ± 13.43 | 33.44 ± 1.47 | 26.44 ± 0.45 | 22.49 ± 1.12 | 17.72 ± 0.13 | 12.09 ± 0.41 | 27.90 ± 1.64 | 24.46 ± 0.78 | 19.99 ± 0.76 | 11.62 ± 0.29 | 22.18 ± 0.62 | 23.94 ± 0.94 |
| (-)-Catechin | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 |
| 5-CQA* | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Apigenin | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Caffeic acid | 0.59 ± 0.02 | 0.97 ± 0.12 | 1.54 ± 0.54 | 0.15 ± 0.01 | 0.10 ± 0.01 | <0.01 | 0.04 ± 0.01 | <0.01 | <0.01 | 0.34 ± 0.02 | <0.01 | <0.01 | 0.24 ± 0.02 |
| Ellagic acid | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 | <0.09 |
| Ferulic acid | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 |
| Hesperetin | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Isorhamnetin | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Kaempferol | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 | <0.06 |
| Myricetin | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Naringin | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 |
| p-Coumaric acid | 5.93 ± 0.09 | 39.13 ± 0.32 | 50.41 ± 0.85 | 11.44 ± 0.50 | 6.47 ± 0.80 | 2.80 ± 0.52 | 5.60 ± 0.51 | 38.26 ± 1.06 | 4.06 ± 0.82 | 18.08 ± 0.45 | 3.41 ± 0.50 | 7.43 ± 0.69 | 6.03 ± 0.80 |
| Quercetin | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Rosmarinic acid | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Rutin | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Sinapic acid | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 | <0.03 |
| Syringic acid | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 |
| Vanillic acid | <0.07 | <0.07 | 0.29 ± 0.11 | <0.07 | <0.07 | <0.07 | 0.13 ± 0.06 | <0.07 | <0.07 | <0.07 | <0.07 | <0.07 | <0.07 |
| Wavenumber, cm−1 | Spectra Structure | Material Type |
|---|---|---|
| 7353 | C–H methyl second combination band | Aliphatic hydrocarbons |
| 7300 | Combination of first overtone of methyl C–H stretching and CH3 bending | Aromatic hydrocarbons |
| 7194 | Combination of first overtone of methyl C–H stretching and CH3 bending | Aliphatic hydrocarbons |
| 7168 | C–H methylene second combination band | Aliphatic hydrocarbons |
| 7163 | Combination of first overtone of methyl C–H stretching and CH3 bending | Aliphatic hydrocarbons |
| 7200–7100 | First overtone of O–H stretching modes | Alcohols |
| 7140–6940 | First overtone of O–H stretching modes from phenols | Phenolic O–H |
| 7085–7067 | Combination of first overtone of methylene C–H stretching and methylene C–H bending modes | Aliphatic hydrocarbons |
| 7057 | Combination of first overtone of C–H and aryl C–H | Aromatic hydrocarbons |
| 7040 | First overtone of O-H | Phenolic O–H |
| 6835, 6743 | First overtone of N-H and CONH2 | Amide/protein |
| 6805 | N-H combination band from primary amides (R-C=O-NH2) | Amide |
| 6705–6683 | First overtone of NH2 | Aromatic amine |
| 5550–4550 | O-H broad band, which is combination of the stretching and bending of the O-H | Polyols |
| 5263 | Second overtone of C=O, C=OOH | Carboxylic acids |
| 5208 | Second overtone of C=O, C=ONH | Amide |
| 5181 | O-H stretching and HOH bending combination | Polysaccharides |
| 4878 | N-H in-plane bend and C-N stretching and N-H in-plane bend combination | Amides/proteins |
| 4866 | N-H stretching and C=O stretching (amide I) combination | Amides/proteins |
| 4854 | Stretching combination of second overtone of N-H and N-H | Amides/proteins |
| 4762 | O-H bending and C-O stretching combination | Polysaccharides |
| 4655–4615 | Combination of C-H stretching and C-C bending | C-H aryl |
| 4400 | CONH2 specifically due to peptide β-sheet structures | Proteins |
| 4365–4370 | CONH2 specifically due to the α-helix peptide structure | Proteins |
| 4360 | First overtone of CH stretching and C–H bending modes | C–H aryl |
| 4348 | Second overtone of C–H stretching mode | Amides |
| 4333 | Combination of first overtone of CH2 asymmetric stretching and CH2 bending modes | Aliphatic hydrocarbons |
| 4314 | Combination of first overtone of CH2 asymmetric stretching and CH2 bending modes | Aliphatic hydrocarbons |
| Sample | Amino Acid * | Polyphenol |
|---|---|---|
| Sample 2 | highest total free amino acid (13.6 ± 0.4 g/100 g) | highest concentration of 4-hydroxibenzoic acid (98.23 ± 1343 mg/kg) |
| highest content of methionine (216 ± 7.0 mg/g) | ||
| Sample 3 | highest concentration of cysteine (185.8 ± 2.6 mg/100 g), glutamic acid (2076 ± 162 mg/100 g) high total free amino acid (12.1 ± 0.2 g/100 g) | highest concentration of p-coumaric acid (50.41 ± 0.85 mg/kg), caffeic acid (1.54 ± 0.54 mg/kg), vanillic acid (0.29 ± 0.11 mg/kg) |
| Sample 8 | highest concentration of ornithine (1128 ± 204 mg/100 g) | high concentration of p-coumaric acid (38.26 ± 1.06 mg/kg) |
| Sample 10 | the lowest concentrations of the following amino acids: proline (68.1 ± 2.08 mg/100 g), glycine (55.6 ± 3.9 mg/100 g), valine (69.7 ± 11.8 mg/100 g), methionine (27.3 ± 3.3 mg/100 g), isoleucine (84.4 ± 10.1 mg/100 g), leucine (82.9 ± 2.2 mg/100 g), tyrosine (95.1 ± 9.8 mg/100 g), phenylalanine (73.3 ± 8.9 mg/100 g), histidine (80.2 ± 3.5 mg/100 g), serine (130 ± 15.3 mg/100 g) | difference from the others is not supported by the measured content parameters |
| Sample 13 | difference from the others is not supported by the measured content parameters | |
| Sample | Frequency | Sum of Ranks | Mean of Ranks | Groups | ||
|---|---|---|---|---|---|---|
| 2nd | 23 | 6148 | 267.304 | A | ||
| 13th | 23 | 3948 | 171.652 | B | ||
| 5th | 23 | 3930 | 170.870 | B | ||
| 12th | 23 | 3898 | 169.478 | B | ||
| 3rd | 23 | 3784 | 164.522 | B | C | |
| 10th | 23 | 3690 | 160.435 | B | C | |
| 9th | 23 | 3312 | 144.000 | B | C | |
| 6th | 23 | 3290 | 143.043 | B | C | |
| 8th | 23 | 3121 | 135.696 | B | C | |
| 11th | 23 | 2811 | 122.217 | B | C | |
| 4th | 23 | 2457 | 106.826 | B | C | |
| 7th | 23 | 2382 | 103.565 | B | C | |
| 1st | 23 | 2079 | 90.391 | C | ||
| 1st | 2nd | 3rd | 4th | 5th | 6th | 7th | 8th | 9th | 10th | 12th | 13th | 14th | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | No | Yes | No | No | Yes | No | No | No | No | No | No | Yes | Yes |
| 2nd | Yes | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| 3rd | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 4th | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 5th | Yes | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 6th | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 7th | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 8th | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 9th | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 10th | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 12th | No | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 13th | Yes | Yes | No | No | No | No | No | No | No | No | No | No | No |
| 14th | Yes | Yes | No | No | No | No | No | No | No | No | No | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matkovits, A.; Fodor, M.; Jókai, Z. Analysis of Polyphenol Patterns of Pleurotus ostreatus Cultivars by UHPLC-ESI-MS/MS; Application of FT-NIR and Chemometric Methods, Classification Options. Chemosensors 2024, 12, 19. https://doi.org/10.3390/chemosensors12020019
Matkovits A, Fodor M, Jókai Z. Analysis of Polyphenol Patterns of Pleurotus ostreatus Cultivars by UHPLC-ESI-MS/MS; Application of FT-NIR and Chemometric Methods, Classification Options. Chemosensors. 2024; 12(2):19. https://doi.org/10.3390/chemosensors12020019
Chicago/Turabian StyleMatkovits, Anna, Marietta Fodor, and Zsuzsa Jókai. 2024. "Analysis of Polyphenol Patterns of Pleurotus ostreatus Cultivars by UHPLC-ESI-MS/MS; Application of FT-NIR and Chemometric Methods, Classification Options" Chemosensors 12, no. 2: 19. https://doi.org/10.3390/chemosensors12020019
APA StyleMatkovits, A., Fodor, M., & Jókai, Z. (2024). Analysis of Polyphenol Patterns of Pleurotus ostreatus Cultivars by UHPLC-ESI-MS/MS; Application of FT-NIR and Chemometric Methods, Classification Options. Chemosensors, 12(2), 19. https://doi.org/10.3390/chemosensors12020019