
Lipedema: Insights into Morphology, Pathophysiology, and Challenges

Abstract
:1. Introduction
2. Lipedema: Morphology
2.1. Components of Adipose Tissue and Its Heterogeneity
2.2. Morphology of Lipedema SAT
2.3. Excess Interstitial Fluid, Contributing Factors, and Consequences
2.4. Stages of Lipedema
3. Mechanisms Implicated in the Pathogenesis of Lipedema
3.1. Altered Gene Expression of Adipogenic and Hormonal Markers in Lipedema
3.2. Impairment of Endothelial Junction and Increased Permeability of Endothelial Cells in Lipedema
3.3. The Altered States of ECM in Lipedema
3.4. Immune-Cell Recruitment and Altered Cytokine Profile in Lipedema
3.5. Vasculature of Lipedema Adipose Tissue
3.6. Altered Lipid Composition and Metabolic Phenotype in Lipedema
3.7. Role of Selenium and Sodium in the Pathology of Lipedema
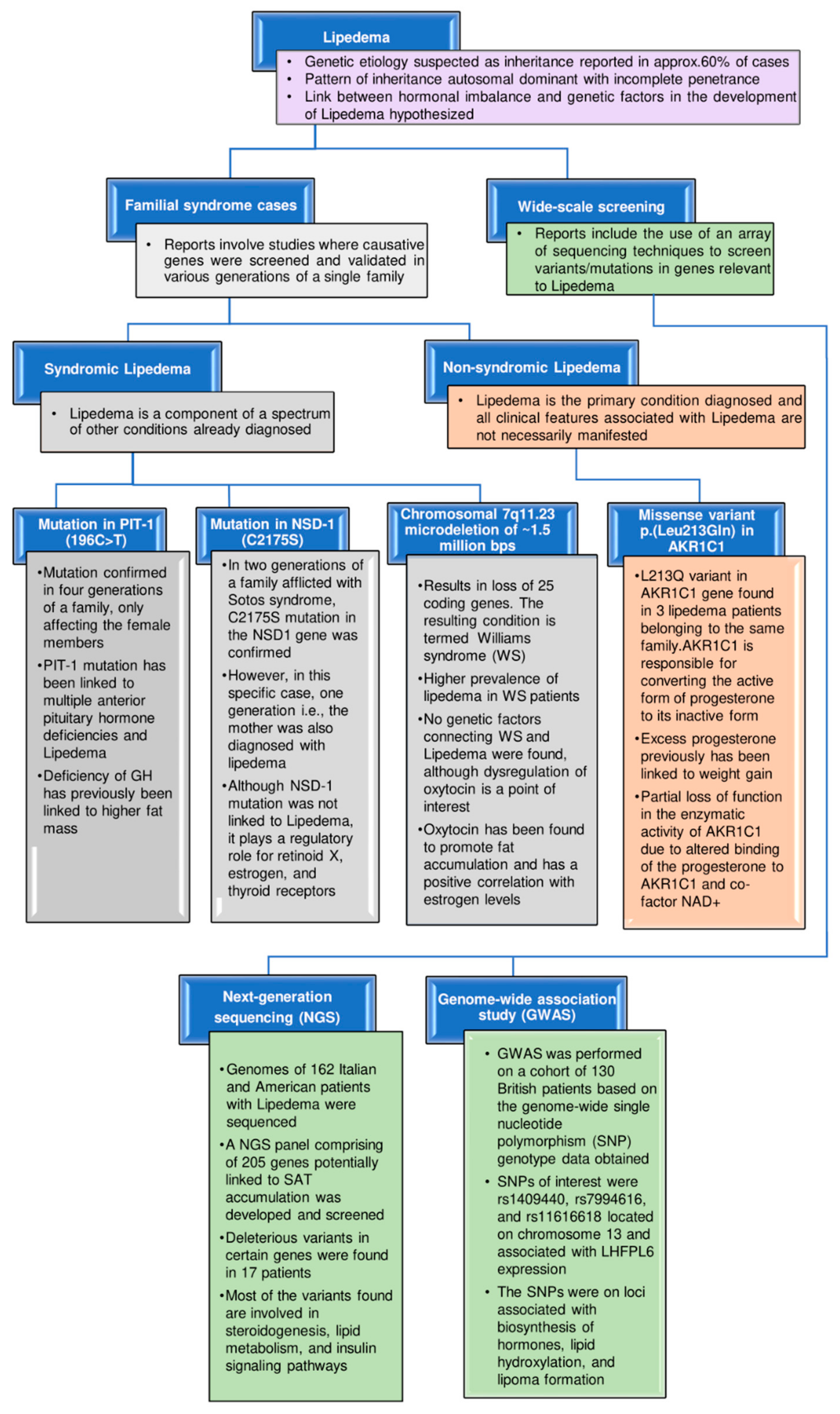
4. Genetic Implications
5. Biomarkers: Current and Future Perspectives to Combat Lipedema
5.1. Platelet Factor 4
5.2. Sodium
5.3. Extracellular Vesicles and Their Contents
5.4. Lipid Profile
5.5. Cytokines
6. Potential Techniques to Study Lipedema
7. Diagnostic Tools and Clinical Studies
8. Current Challenges with Lipedema

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes (Involved in) | Known Functi [188] on | Expression Level in Lipedema Samples (Lipoaspirates/SVF/2D ADSCs/3D Spheroids Derived from ADSCs) as Compared to Healthy Lean Controls |
|---|---|---|
| Synthesis of adipokines | ||
| ADIPOQ | ||
| LEP | ||
| CFD |
|
|
| Transcriptional regulation of adipogenesis | ||
| PPARG | ||
| C/EBPD |
|
|
| KLF4 |
|
|
| NCOR2 |
|
|
| ZNF423 |
|
|
| Modulation of insulin sensitivity | ||
| INSR |
|
|
| GLUT4 |
| |
| Modulation of cell cycle and proliferation | ||
| CCND1 |
|
|
| CDC20 |
|
|
| CENPF |
|
|
| BIRC5 |
|
|
| KIF14 |
|
|
| BUB1 |
| |
| Lipid metabolism | ||
| LPL |
|
|
| FABP4 |
|
|
| Lymphatic function | ||
| VEGFR-3 |
| |
| PDPN |
| |
| PROX-1 |
| |
| LYVE1 |
| |
| CCL21 |
|
|
| Maintaining endothelial barrier | ||
| CDH5 |
| |
| TIE-2 | ||
| Thermogenic function | ||
| UCP1 |
| |
| Inflammatory processes | ||
| CD11c (ITGAX) |
|
|
| Altered micro-RNAs | ||
| miR-16-5p |
|
|
| miR-29a-3p |
|
|
| miR-454-3p |
|
|
| Hormonal regulation | ||
| CYP19A1 |
|
|
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- La Torre, Y.S.-D.; Wadeea, R.; Rosas, V.; Herbst, K.L. Lipedema: Friend and Foe. Horm. Mol. Biol. Clin. Investig. 2018, 33, 1. [Google Scholar] [CrossRef] [PubMed]
- Buso, G.; Depairon, M.; Tomson, D.; Raffoul, W.; Vettor, R.; Mazzolai, L. Lipedema: A Call to Action! Obesity 2019, 27, 1567. [Google Scholar] [CrossRef] [PubMed]
- Shavit, E.; Wollina, U.; Alavi, A. Lipoedema Is Not Lymphoedema: A Review of Current Literature. Int. Wound J. 2018, 15, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Herbst, K.L. Rare Adipose Disorders (RADs) Masquerading as Obesity. Acta Pharmacol. Sin. 2012, 33, 155–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, D.W.; Herbst, K.L. Lipedema: A Relatively Common Disease with Extremely Common Misconceptions. Plast. Reconstr. Surg. Glob. Open 2016, 4, e1043. [Google Scholar] [CrossRef] [Green Version]
- Herbst, K.L.; Kahn, L.A.; Iker, E.; Ehrlich, C.; Wright, T.; McHutchison, L.; Schwartz, J.; Sleigh, M.; Donahue, P.M.C.; Lisson, K.H.; et al. Standard of Care for Lipedema in the United States. Phlebology 2021, 36, 779–796. [Google Scholar] [CrossRef]
- Suga, H.; Araki, J.; Aoi, N.; Kato, H.; Higashino, T.; Yoshimura, K. Adipose Tissue Remodeling in Lipedema: Adipocyte Death and Concurrent Regeneration. J. Cutan. Pathol. 2009, 36, 1293–1298. [Google Scholar] [CrossRef]
- Crescenzi, R.; Marton, A.; Donahue, P.M.C.; Mahany, H.B.; Lants, S.K.; Wang, P.; Beckman, J.A.; Donahue, M.J.; Titze, J. Tissue Sodium Content Is Elevated in the Skin and Subcutaneous Adipose Tissue in Women with Lipedema. Obesity 2018, 26, 310–317. [Google Scholar] [CrossRef]
- Forner-Cordero, I.; Szolnoky, G.; Forner-Cordero, A.; Kemény, L. Lipedema: An Overview of Its Clinical Manifestations, Diagnosis and Treatment of the Disproportional Fatty Deposition Syndrome—Systematic Review. Clin. Obes. 2012, 2, 86–95. [Google Scholar] [CrossRef]
- Child, A.H.; Gordon, K.D.; Sharpe, P.; Brice, G.; Ostergaard, P.; Jeffery, S.; Mortimer, P.S. Lipedema: An Inherited Condition. Am. J. Med. Genet. Part A 2010, 152, 970–976. [Google Scholar] [CrossRef]
- Reich-Schupke, S.; Schmeller, W.; Brauer, W.J.; Cornely, M.E.; Faerber, G.; Ludwig, M.; Lulay, G.; Miller, A.; Rapprich, S.; Richter, D.F.; et al. S1 Guidelines: Lipedema. JDDG J. Dtsch. Dermatol. Ges. 2017, 15, 758–767. [Google Scholar] [CrossRef]
- Bauer, A.T.; Von Lukowicz, D.; Lossagk, K.; Hopfner, U.; Kirsch, M.; Moog, P.; Bauer, H.; Machens, H.G.; Schmauss, D. Adipose Stem Cells from Lipedema and Control Adipose Tissue Respond Differently to Adipogenic Stimulation In Vitro. Plast. Reconstr. Surg. 2019, 144, 623–632. [Google Scholar] [CrossRef]
- Lee, M.J.; Wu, Y.; Fried, S.K. Adipose Tissue Heterogeneity: Implication of Depot Differences in Adipose Tissue for Obesity Complications. Mol. Asp. Med. 2013, 34, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, E.; Wing, A.; Holtrup, B.; Sebo, Z.; Kaplan, J.L.; Saavedra-Peña, R.; Church, C.D.; Colman, L.; Berry, R.; Rodeheffer, M.S. The Adipose Tissue Microenvironment Regulates Depot-Specific Adipogenesis in Obesity. Cell Metab. 2016, 24, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Ioannidou, A.; Fisher, R.M.; Hagberg, C.E. The Multifaceted Roles of the Adipose Tissue Vasculature. Obes. Rev. 2022, 23, e13403. [Google Scholar] [CrossRef]
- Van Dijk, C.G.M.; Nieuweboer, F.E.; Pei, J.Y.; Xu, Y.J.; Burgisser, P.; Van Mulligen, E.; El Azzouzi, H.; Duncker, D.J.; Verhaar, M.C.; Cheng, C. The Complex Mural Cell: Pericyte Function in Health and Disease. Int. J. Cardiol. 2015, 190, 75–89. [Google Scholar] [CrossRef]
- Krüger-Genge, A.; Blocki, A.; Franke, R.P.; Jung, F. Vascular Endothelial Cell Biology: An Update. Int. J. Mol. Sci. 2019, 20, 4411. [Google Scholar] [CrossRef] [Green Version]
- Hasan, S.S.; Fischer, A. The Endothelium: An Active Regulator of Lipid and Glucose Homeostasis. Trends Cell Biol. 2021, 31, 37–49. [Google Scholar] [CrossRef]
- Hammarstedt, A.; Gogg, S.; Hedjazifar, S.; Nerstedt, A.; Smith, U. Impaired Adipogenesis and Dysfunctional Adipose Tissue in Human Hypertrophic Obesity. Physiol. Rev. 2018, 98, 1911–1941. [Google Scholar] [CrossRef] [Green Version]
- Yucel, N.; Arany, Z. Fat, Obesity, and the Endothelium. Curr. Opin. Physiol. 2019, 12, 44–50. [Google Scholar] [CrossRef]
- Rutkowski, J.M.; Davis, K.E.; Scherer, P.E. Mechanisms of Obesity and Related Pathologies: The Macro- and Microcirculation of Adipose Tissue. FEBS J. 2009, 276, 5738–5746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felmerer, G.; Stylianaki, A.; Hägerling, R.; Wang, A.; Ströbel, P.; Hollmén, M.; Lindenblatt, N.; Gousopoulos, E. Adipose Tissue Hypertrophy, an Aberrant Biochemical Profile and Distinct Gene Expression in Lipedema. J. Surg. Res. 2020, 253, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghadban, S.; Cromer, W.; Allen, M.; Ussery, C.; Badowski, M.; Harris, D.; Herbst, K.L. Dilated Blood and Lymphatic Microvessels, Angiogenesis, Increased Macrophages, and Adipocyte Hypertrophy in Lipedema Thigh Skin and Fat Tissue. J. Obes. 2019, 2019, 8747461. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, M.; Bandara, N.; Morgan, S.; Nowell, C.; Mehdi, A.M.; Lyu, R.; McCarthy, D.; Anderson, D.; Creek, D.J.; Achen, M.G.; et al. Key Signaling Networks Are Dysregulated in Patients with the Adipose Tissue Disorder, Lipedema. Int. J. Obes. 2022, 46, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, M.; Eekema, A.; Ussery, C.; Neuhardt, D.; Garby, K.; Herbst, K.L. Subcutaneous Adipose Tissue Therapy Reduces Fat by Dual X-ray Absorptiometry Scan and Improves Tissue Structure by Ultrasound in Women with Lipoedema and Dercum Disease. Clin. Obes. 2018, 8, 398–406. [Google Scholar] [CrossRef]
- Shin, B.W.; Sim, Y.-J.; Jeong, H.J.; Kim, G.C. Lipedema, a Rare Disease. Ann. Rehabil. Med. 2011, 35, 922–927. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.; Schwartz, M.; Herbst, K.L. Interstitial Fluid in Lipedema and Control Skin. Women’s Health Rep. 2020, 1, 480–487. [Google Scholar] [CrossRef]
- L’Hermitte, F.; Behar, A.; Pariès, J.; Cohen-Boulakia, F.; Attali, J.R.; Valensi, P. Impairment of Lymphatic Function in Women with Gynoid Adiposity and Swelling Syndrome. Metabolism 2003, 52, 805–809. [Google Scholar] [CrossRef]
- Shah, J.P.; Danoff, J.V.; Desai, M.J.; Parikh, S.; Nakamura, L.Y.; Phillips, T.M.; Gerber, L. Biochemicals Associated with Pain and Inflammation Are Elevated in Sites Near to and Remote from Active Myofascial Trigger Points. Arch. Phys. Med. Rehabil. 2008, 89, 16–23. [Google Scholar] [CrossRef]
- Szolnoky, G.; Nagy, N.; Kovács, R.K.; Dósa-Rácz, É.; Szabó, A.; Bársony, K.; Balogh, M.; Kemény, L. Complex Decongestive Physiotherapy Decreases Capillary Fragility in Lipedema. Lymphology 2008, 41, 161–166. [Google Scholar]
- Guimberteau, J.C. Approche Endoscopique des Aponévroses. Continuité Fibrillaire dans les Fascia et Aponévroses. Ann. Chir. Plast. Esthétique 2012, 57, 465–466. [Google Scholar] [CrossRef]
- Felmerer, G.; Stylianaki, A.; Hollmén, M.; Ströbel, P.; Stepniewski, A.; Wang, A.; Frueh, F.S.; Kim, B.S.; Giovanoli, P.; Lindenblatt, N.; et al. Increased Levels of VEGF-C and Macrophage Infiltration in Lipedema Patients without Changes in Lymphatic Vascular Morphology. Sci. Rep. 2020, 10, 10947. [Google Scholar] [CrossRef]
- Paolacci, S.; Precone, V.; Acquaviva, F.; Chiurazzi, P.; Fulcheri, E.; Pinelli, M.; Buffelli, F.; Michelini, S.; Herbst, K.L.; Unfer, V.; et al. Genetics of Lipedema: New Perspectives on Genetic Research and Molecular Diagnoses. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5581–5594. [Google Scholar] [CrossRef]
- Eklof, B.; Perrin, M.; Delis, K.T.; Rutherford, R.B.; Gloviczki, P. Updated Terminology of Chronic Venous Disorders: The VEIN-TERM Transatlantic Interdisciplinary Consensus Document. J. Vasc. Surg. 2009, 49, 498–501. [Google Scholar] [CrossRef] [Green Version]
- Wiedner, M.; Aghajanzadeh, D.; Richter, D.F. Differential Diagnoses and Treatment of Lipedema. Plast. Aesthetic Res. 2020, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Al-Ghadban, S.; Pursell, I.A.; Diaz, Z.T.; Herbst, K.L.; Bunnell, B.A. 3D Spheroids Derived from Human Lipedema Ascs Demonstrated Similar Adipogenic Differentiation Potential and Ecm Remodeling to Non-Lipedema Ascs In Vitro. Int. J. Mol. Sci. 2020, 21, 8350. [Google Scholar] [CrossRef]
- Fife, C.E.; Maus, E.A.; Carter, M.J. Lipedema: A Frequently Misdiagnosed and Misunderstood Fatty Deposition Syndrome. Adv. Ski. Wound Care 2010, 23, 81–92. [Google Scholar] [CrossRef]
- Herbst, K.L.; Mirkovskaya, L.; Bharhagava, A.; Chava, Y.; Te, C. Lipedema Fat and Signs and Symptoms of Illness, Increase with Advancing Stage. Arch. Med. 2015, 7, 1–8. [Google Scholar]
- Rasmussen, J.C.; Aldrich, M.B.; Fife, C.E.; Herbst, K.L.; Sevick-Muraca, E.M. Lymphatic Function and Anatomy in Early Stages of Lipedema. Obesity 2022, 30, 1391–1400. [Google Scholar] [CrossRef]
- Katzer, K.; Hill, J.L.; McIver, K.B.; Foster, M.T. Lipedema and the Potential Role of Estrogen in Excessive Adipose Tissue Accumulation. Int. J. Mol. Sci. 2021, 22, 11720. [Google Scholar] [CrossRef]
- Siems, W.; Grune, T.; Voss, P.; Brenke, R. Anti-Fibrosclerotic Effects of Shock Wave Therapy in Lipedema and Cellulite. BioFactors 2005, 24, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Senocak, F.S.; Si, Y.; Moya, C.; Russell, W.K.; Russell, D.H.; Lee, K.; Jayaraman, A. Effect of Uncoupling Protein-1 Expression on 3T3-L1 Adipocyte Gene Expression. FEBS Lett. 2007, 581, 5865–5871. [Google Scholar] [CrossRef] [PubMed]
- Strohmeier, K.; Hofmann, M.; Jacak, J.; Narzt, M.-S.; Wahlmueller, M.; Mairhofer, M.; Schaedl, B.; Holnthoner, W.; Barsch, M.; Sandhofer, M.; et al. Multi-Level Analysis of Adipose Tissue Reveals the Relevance of Perivascular Subpopulations and an Increased Endothelial Permeability in Early-Stage Lipedema. Biomedicines 2022, 10, 1163. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghadban, S.; Diaz, Z.T.; Singer, H.J.; Mert, K.B.; Bunnell, B.A. Increase in Leptin and PPAR-γ Gene Expression in Lipedema Adipocytes Differentiated In Vitro from Adipose-Derived Stem Cells. Cells 2020, 9, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamm, J.K.; El Jack, A.K.; Pilch, P.F.; Farmer, S.R. Role of PPARγ in Regulating Adipocyte Differentiation and Insulin-Responsive Glucose Uptake. Ann. N. Y. Acad. Sci. 1999, 892, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPα Induces Adipogenesis through PPARγ: A Unified Pathway. Genes Dev. 2002, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Keith, L.; Seo, C.A.; Rowsemitt, C.; Pfeffer, M.; Wahi, M.; Staggs, M.; Dudek, J.; Gower, B.; Carmody, M. Ketogenic Diet as a Potential Intervention for Lipedema. Med. Hypotheses 2021, 146, 110435. [Google Scholar] [CrossRef]
- Klöting, N.; Koch, L.; Wunderlich, T.; Kern, M.; Ruschke, K.; Krone, W.; Brüning, J.C.; Blüher, M. Autocrine IGF-1 Action in Adipocytes Controls Systemic IGF-1 Concentrations and Growth. Diabetes 2008, 57, 2074. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hudak, C.; Sul, H.S. Role of Preadipocyte Factor 1 in Adipocyte Differentiation. Clin. Lipidol. 2010, 5, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Nøhr, J.; Jensen, C.H.; Petersen, R.K.; Bachmann, E.; Teisner, B.; Larsen, L.K.; Mandrup, S.; Kristiansen, K. Insulin-like Growth Factor-1/Insulin Bypasses Pref-1/FA1-Mediated Inhibition of Adipocyte Differentiation. J. Biol. Chem. 2003, 278, 20906–20914. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.Q.; Otto, T.C.; Daniel Lane, M. Mitotic Clonal Expansion: A Synchronous Process Required for Adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhao, Y.; Wu, R.; Jiang, Q.; Cai, M.; Bi, Z.; Liu, Y.; Yao, Y.; Feng, J.; Wang, Y.; et al. RNA Biology ZFP217 Regulates Adipogenesis by Controlling Mitotic Clonal Expansion in a METTL3-m 6 A Dependent Manner. RNA Biol. 2019, 16, 1785–1793. [Google Scholar] [CrossRef]
- He, Y.; Wu, Y.; Mei, B.; Fang, X.; Cai, G.; Cai, N.; Wu, Q.; Huang, Z.; Ge, C.; Liang, H.; et al. A Small Nucleolar RNA, SNORD126, Promotes Adipogenesis in Cells and Rats by Activating the PI3K–AKT Pathway. J. Cell. Physiol. 2021, 236, 3001–3014. [Google Scholar] [CrossRef]
- Gupta, R.K.; Arany, Z.; Seale, P.; Mepani, R.J.; Ye, L.; Conroe, H.M.; Roby, Y.A.; Kulaga, H.; Reed, R.R.; Spiegelman, B.M. Transcriptional Control of Preadipocyte Determination by Zfp423. Nature 2010, 464, 619–623. [Google Scholar] [CrossRef] [Green Version]
- Ünlü, Z.; ÇAVUŞOĞLU, T.; Kosova, F.; Aydin, A. The Relationship Between Adipokines and the Thickness of Subcutaneous Adipose Tissue in Lipedema. Celal Bayar Üniversitesi Sağlık Bilimleri Enstitüsü Derg. 2022, 9, 296–304. [Google Scholar] [CrossRef]
- Krotkiewski, M.; Bjorntorp, P.; Sjostrom, L.; Smith, U. Impact of Obesity on Metabolism in Men and Women. Importance of Regional Adipose Tissue Distribution. J. Clin. Investig. 1983, 72, 1150–1162. [Google Scholar] [CrossRef]
- Lee, C.G.; Carr, M.C.; Murdoch, S.J.; Mitchell, E.; Woods, N.F.; Wener, M.H.; Chandler, W.L.; Boyko, E.J.; Brunzell, J.D. Adipokines, Inflammation, and Visceral Adiposity across the Menopausal Transition: A Prospective Study. J. Clin. Endocrinol. Metab. 2009, 94, 1104–1110. [Google Scholar] [CrossRef] [Green Version]
- Ingle, J.N.; Liu, M.; Lawrence Wickerham, D.; Schaid, D.J.; Wang, L.; Mushiroda, T.; Kubo, M.; Costantino, J.P.; Vogel, V.G.; Paik, S.; et al. Selective Estrogen Receptor Modulators and Pharmacogenomic Variation in ZNF423 Regulation of BRCA1 Expression: Individualized Breast Cancer Prevention. Cancer Discov. 2013, 3, 812–825. [Google Scholar] [CrossRef] [Green Version]
- Priglinger, E.; Wurzer, C.; Steffenhagen, C.; Maier, J.; Hofer, V.; Peterbauer, A.; Nuernberger, S.; Redl, H.; Wolbank, S.; Sandhofer, M. The Adipose Tissue–Derived Stromal Vascular Fraction Cells from Lipedema Patients: Are They Different? Cytotherapy 2017, 19, 849–860. [Google Scholar] [CrossRef]
- Wolf, S.; Deuel, J.W.; Hollmén, M.; Felmerer, G.; Kim, B.S.; Vasella, M.; Grünherz, L.; Giovanoli, P.; Lindenblatt, N.; Gousopoulos, E. A Distinct Cytokine Profile and Stromal Vascular Fraction Metabolic Status without Significant Changes in the Lipid Composition Characterizes Lipedema. Int. J. Mol. Sci. 2021, 22, 3313. [Google Scholar] [CrossRef]
- Schneider, M.; Conway, E.M.; Carmeliet, P. Lymph Makes You Fat. Nat. Genet. 2005, 37, 1023–1024. [Google Scholar] [CrossRef] [PubMed]
- Andrei, A.M.; Berbecaru-Iovan, A.; Din-Anghel, F.R.I.; Stănciulescu, C.E.; Berbecaru-Iovan, S.; Baniţă, I.M.; Pisoschi, C.G.C.G.; Din-Anghel, F.R.I.; Stanciulescu, C.E.; Berbecaru-Iovan, S.; et al. Interplay between Hypoxia, Inflammation and Adipocyte Remodeling in the Metabolic Syndrome. Hypoxia Hum. Dis. 2017. [Google Scholar] [CrossRef]
- Parikh, S.M. Angiopoietins and Tie2 in Vascular Inflammation. Curr. Opin. Hematol. 2017, 24, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.A.; Mendez, U.; Gilbert, R.J.; Keim, A.P.; Goldman, J. Increased Hyaluronan Expression at Distinct Time Points in Acute Lymphedema. Lymphat. Res. Biol. 2012, 10, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Pessentheiner, A.R.; Ducasa, G.M.; Gordts, P.L.S.M.S.M.; Reijmers, R.M.; Apice, L.D.; Pessentheiner, A.R.; Gordts, P.L.S.M.; Ducasa, G.M. Proteoglycans in Obesity-Associated Metabolic Dysfunction and Meta-Inflammation. Front. Immunol. 2020, 11, 769. [Google Scholar] [CrossRef]
- Levick, J.R.; Michel, C.C.; Charles Michel, C. Microvascular Fluid Exchange and the Revised Starling Principle. Cardiovasc. Res. 2010, 87, 198–210. [Google Scholar] [CrossRef]
- Tarbell, J.M.; Simon, S.I.; Curry, F.R.E. Mechanosensing at the Vascular Interface. Annu. Rev. Biomed. Eng. 2014, 16, 505–532. [Google Scholar] [CrossRef] [Green Version]
- Schierke, F.; Wyrwoll, M.J.; Wisdorf, M.; Niedzielski, L.; Maase, M.; Ruck, T.; Meuth, S.G.; Kusche-Vihrog, K. Nanomechanics of the Endothelial Glycocalyx Contribute to Na+-Induced Vascular Inflammation. Sci. Rep. 2017, 7, srep46476. [Google Scholar] [CrossRef] [Green Version]
- Huber, J.; Löffler, M.; Bilban, M.; Reimers, M.; Kadl, A.; Todoric, J.; Zeyda, M.; Geyeregger, R.; Schreiner, M.; Weichhart, T.; et al. Prevention of High-Fat Diet-Induced Adipose Tissue Remodeling in Obese Diabetic Mice by n-3 Polyunsaturated Fatty Acids. Int. J. Obes. 2006, 31, 1004–1013. [Google Scholar] [CrossRef] [Green Version]
- Goddi, A.; Carmona, A.; Schroedl, L.; White, J.M.; Piron, M.J.; De Leon, A.; Casimiro, I.; Hoffman, A.; Gonzalez Porras, M.A.; Brey, E.M.; et al. Laminin-A4 Is Upregulated in Both Human and Murine Models of Obesity. Front. Endocrinol. 2021, 12, 886. [Google Scholar] [CrossRef]
- Pasarica, M.; Gowronska-Kozak, B.; Burk, D.; Remedios, I.; Hymel, D.; Gimble, J.; Ravussin, E.; Bray, G.A.; Smith, S.R. Adipose Tissue Collagen VI in Obesity. J. Clin. Endocrinol. Metab. 2009, 94, 5155. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fu, Y.; Yan, Z.; Zhang, X.B.; Pei, M. Impact of Fibronectin Knockout on Proliferation and Differentiation of Human Infrapatellar Fat Pad-Derived Stem Cells. Front. Bioeng. Biotechnol. 2019, 7, 321. [Google Scholar] [CrossRef]
- Mao, Y.; Schwarzbauer, J.E. Fibronectin Fibrillogenesis, a Cell-Mediated Matrix Assembly Process. Matrix Biol. 2005, 24, 389–399. [Google Scholar] [CrossRef]
- Arcidiacono, B.; Chiefari, E.; Laria, A.E.; Messineo, S.; Bilotta, F.L.; Britti, D.; Foti, D.P.; Foryst-Ludwig, A.; Kintscher, U.; Brunetti, A. Expression of Matrix Metalloproteinase-11 Is Increased under Conditions of Insulin Resistance. World J. Diabetes 2017, 8, 422. [Google Scholar] [CrossRef]
- Gesta, S.; Guntur, K.; Majumdar, I.D.; Akella, S.; Vishnudas, V.K.; Sarangarajan, R.; Narain, N.R. Reduced Expression of Collagen VI Alpha 3 (COL6A3) Confers Resistance to Inflammation-Induced MCP1 Expression in Adipocytes. Obesity 2016, 24, 1695–1703. [Google Scholar] [CrossRef]
- Kim, H.N.; Chung, H.S. Caveolin-1 Inhibits Membrane-Type 1 Matrix Metalloproteinase Activity. BMB Rep. 2008, 41, 858–862. [Google Scholar] [CrossRef] [Green Version]
- Kruglikov, I.L.; Joffin, N.; Scherer, P.E. The MMP14–Caveolin Axis and Its Potential Relevance for Lipoedema. Nat. Rev. Endocrinol. 2020, 16, 669–674. [Google Scholar] [CrossRef]
- Priglinger, E.; Strohmeier, K.; Weigl, M.; Lindner, C.; Auer, D.; Gimona, M.; Barsch, M.; Jacak, J.; Redl, H.; Grillari, J.; et al. SVF-Derived Extracellular Vesicles Carry Characteristic MiRNAs in Lipedema. Sci. Rep. 2020, 10, 7211. [Google Scholar] [CrossRef]
- Pan, Z.; Zhou, Z.; Zhang, H.; Zhao, H.; Song, P.; Wang, D.; Yin, J.; Zhao, W.; Xie, Z.; Wang, F.; et al. CD90 Serves as Differential Modulator of Subcutaneous and Visceral Adipose-Derived Stem Cells by Regulating AKT Activation That Influences Adipose Tissue and Metabolic Homeostasis. Stem Cell Res. Ther. 2019, 10, 355. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Liu, J.; Chen, G.; Du, J.; Cai, H.; Chen, X.; Ye, G.; Luo, Y.; Luo, Y.; Zhang, L.; et al. CD146 Is a Novel ANGPTL2 Receptor That Promotes Obesity by Manipulating Lipid Metabolism and Energy Expenditure. Adv. Sci. 2021, 8, 2004032. [Google Scholar] [CrossRef]
- Navarro, R.; Compte, M.; Álvarez-Vallina, L.; Sanz, L. Immune Regulation by Pericytes: Modulating Innate and Adaptive Immunity. Front. Immunol. 2016, 7, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, T.A.; Shousha, W.G.; Abdo, S.M.; Mohamed, I.K.; El-Badri, N. Human Adipose-Derived Pericytes: Biological Characterization and Reprogramming into Induced Pluripotent Stem Cells. Cell. Physiol. Biochem. 2020, 54, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, M.; Sul, O.J.; Choi, E.K.; Kim, J.E.; Suh, J.H.; Choi, H.S. MCP-1 Deficiency Enhances Browning of Adipose Tissue via Increased M2 Polarization. J. Endocrinol. 2019, 242, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mei, H.; Chang, X.; Chen, F.; Zhu, Y.; Han, X. Adipocyte-Derived Microvesicles from Obese Mice Induce M1 Macrophage Phenotype through Secreted MiR-155. J. Mol. Cell Biol. 2016, 8, 505–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, D.; Apovian, C. Macrophage Functions in Lean and Obese Adipose Tissue. Metabolism 2017, 72, 120–143. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Shi, J.; Chen, L.; Lv, Z.; Chen, X.; Cao, H.; Xiang, Z.; Han, X. M2 Macrophages Promote Myofibroblast Differentiation of LR-MSCs and Are Associated with Pulmonary Fibrogenesis 11 Medical and Health Sciences 1102 Cardiorespiratory Medicine and Haematology. Cell Commun. Signal. 2018, 16, 89. [Google Scholar] [CrossRef] [Green Version]
- Freund, A.; Chauveau, C.; Brouillet, J.P.; Lucas, A.; Lacroix, M.; Licznar, A.; Vignon, F.; Lazennec, G. IL-8 Expression and Its Possible Relationship with Estrogen-Receptor-Negative Status of Breast Cancer Cells. Oncogene 2003, 22, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Huang, R.P.; Chen, L.; Li, S.; Shi, Q.; Jordan, C.; Huang, R.P. Identification of Interleukin-8 as Estrogen Receptor-Regulated Factor Involved in Breast Cancer Invasion and Angiogenesis by Protein Arrays. Int. J. Cancer 2004, 109, 507–515. [Google Scholar] [CrossRef]
- Rissanen, T.T.; Markkanen, J.E.; Gruchala, M.; Heikura, T.; Puranen, A.; Kettunen, M.I.; Kholová, I.; Kauppinen, R.A.; Achen, M.G.; Stacker, S.A.; et al. VEGF-D Is the Strongest Angiogenic and Lymphangiogenic Effector Among VEGFs Delivered into Skeletal Muscle via Adenoviruses. Circ. Res. 2003, 92, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.O. Vascular Endothelial Growth Factors and Vascular Permeability. Cardiovasc. Res. 2010, 87, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, W.L.; Jain, P.; Ambujam, G.; Zaleska, M.; Cakala, M. Where Do Lymph and Tissue Fluid Accumulate in Lymphedema of the Lower Limbs Caused by Obliteration of Lymphatic Collectors? Lymphology 2009, 42, 105–111. [Google Scholar]
- Taylor, M.J. The Nature and Significance of Body Image Disturbance. Ph.D. Thesis, University of Cambridge, Cambridge, UK, 1987. [Google Scholar]
- Curri, S.B.; Ryan, T.J. Panniculopathy and Fibrosclerosis of the Female Breast and Thigh. Clin. Dermatol. 1989, 7, 107–119. [Google Scholar] [CrossRef]
- Bilancini, S.; Lucchi, M.; Tucci, S.; Eleuteri, P. Functional Lymphatic Alterations in Patients Suffering from Lipedema. Angiology 1995, 46, 333–339. [Google Scholar] [CrossRef]
- Forner-Cordero, I.; Oliván-Sasot, P.; Ruiz-Llorca, C.; Muñoz-Langa, J. Lymphoscintigraphic Findings in Patients with Lipedema. Rev. Española Med. Nucl. E Imagen Mol. 2018, 37, 341–348. [Google Scholar] [CrossRef]
- Blum, K.S.; Karaman, S.; Proulx, S.T.; Ochsenbein, A.M.; Luciani, P.; Leroux, J.C.; Wolfrum, C.; Detmar, M. Chronic High-Fat Diet Impairs Collecting Lymphatic Vessel Function in Mice. PLoS ONE 2014, 9, e94713. [Google Scholar] [CrossRef]
- Harvey, N.L.; Srinivasan, R.S.; Dillard, M.E.; Johnson, N.C.; Witte, M.H.; Boyd, K.; Sleeman, M.W.; Oliver, G. Lymphatic Vascular Defects Promoted by Prox1 Haploinsufficiency Cause Adult-Onset Obesity. Nat. Genet. 2005, 37, 1072–1081. [Google Scholar] [CrossRef]
- Cueni, L.N.; Detmar, M. The Lymphatic System in Health and Disease. Lymphat. Res. Biol. 2008, 6, 109–122. [Google Scholar] [CrossRef]
- Van Geest, A.J.; Esten, S.C.A.M.; Cambier, J.P.R.A.; Gielen, E.G.J.; Kessels, A.; Neumann, H.A.M.; Van Kroonenburgh, M.J.P.G. Lymphatic Disturbances in Lipoedema. Phlebologie 2003, 32, 138–142. [Google Scholar] [CrossRef]
- Van de Pas, C.B.; Sm Boonen, R.; Stevens, S.; Willemsen, S.; Valkema, R.; Neumann, M.; Boonen, R.S.M.; Stevens, S.; Willemsen, S.; Valkema, R.; et al. Does Tumescent Liposuction Damage the Lymph Vessels in Lipoedema Patients? Phlebology 2020, 35, 231–236. [Google Scholar] [CrossRef]
- Lohrmann, C.; Foeldi, E.; Langer, M. MR Imaging of the Lymphatic System in Patients with Lipedema and Lipo-Lymphedema. Microvasc. Res. 2009, 77, 335–339. [Google Scholar] [CrossRef]
- Escobedo, N.; Proulx, S.T.; Karaman, S.; Dillard, M.E.; Johnson, N.; Detmar, M.; Oliver, G. Restoration of Lymphatic Function Rescues Obesity in Prox1-Haploinsufficient Mice. JCI Insight 2016, 1, 85096. [Google Scholar] [CrossRef]
- Fatima, L.A.; Campello, R.S.; Santos, R.D.S.; Freitas, H.S.; Frank, A.P.; Machado, U.F.; Clegg, D.J. Estrogen Receptor 1 (ESR1) Regulates VEGFA in Adipose Tissue. Sci. Rep. 2017, 7, 16716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, M.R.; Simonsen, L.; Karlsmark, T.; Lanng, C.; Bülow, J. Higher Vascular Endothelial Growth Factor-C Concentration in Plasma Is Associated with Increased Forearm Capillary Filtration Capacity in Breast Cancer-Related Lymphedema. Physiol. Rep. 2015, 3, e12403. [Google Scholar] [CrossRef] [PubMed]
- Gousopoulos, E.; Proulx, S.T.; Bachmann, S.B.; Dieterich, L.C.; Scholl, J.; Karaman, S.; Bianchi, R.; Detmar, M. An Important Role of VEGF-C in Promoting Lymphedema Development. J. Investig. Dermatol. 2017, 137, 1995–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.C.; Kim, B.R.; Lee, S.Y.; Park, T.S. Sphingolipid Metabolism and Obesity-Induced Inflammation. Front. Endocrinol. 2013, 4, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Cuenca, S.; Pellegrinelli, V.; Campbell, M.; Oresic, M.; Vidal-Puig, A. Sphingolipids and Glycerophospholipids—The “Ying and Yang” of Lipotoxicity in Metabolic Diseases. Prog. Lipid Res. 2017, 66, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Hammerschmidt, P.; Ostkotte, D.; Nolte, H.; Gerl, M.J.; Jais, A.; Brunner, H.L.; Sprenger, H.G.; Awazawa, M.; Nicholls, H.T.; Turpin-Nolan, S.M.; et al. CerS6-Derived Sphingolipids Interact with Mff and Promote Mitochondrial Fragmentation in Obesity. Cell 2019, 177, 1536–1552.e23. [Google Scholar] [CrossRef]
- Sedger, L.M.; Tull, D.L.; McConville, M.J.; De Souza, D.P.; Rupasinghe, T.W.T.; Williams, S.J.; Dayalan, S.; Lanzer, D.; Mackie, H.; Lam, T.C.; et al. Lipidomic Profiling of Adipose Tissue Reveals an Inflammatory Signature in Cancer-Related and Primary Lymphedema. PLoS ONE 2016, 11, e0154650. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jiang, C.T.; Song, J.Y.; Song, Q.Y.; Ma, J.; Wang, H.J. Lipidomic Profile Revealed the Association of Plasma Lysophosphatidylcholines with Adolescent Obesity. Biomed Res. Int. 2019, 2019, 1382418. [Google Scholar] [CrossRef] [Green Version]
- Piro, M.C.; Tesauro, M.; Lena, A.M.; Gentileschi, P.; Sica, G.; Rodia, G.; Annicchiarico-Petruzzelli, M.; Rovella, V.; Cardillo, C.; Melino, G.; et al. Free-Amino Acid Metabolic Profiling of Visceral Adipose Tissue from Obese Subjects. Amino Acids 2020, 52, 1125–1137. [Google Scholar] [CrossRef]
- Galván-Peña, S.; O’Neill, L.A.J. Metabolic Reprograming in Macrophage Polarization. Front. Immunol. 2014, 5, 420. [Google Scholar] [CrossRef] [Green Version]
- Vats, D.; Mukundan, L.; Odegaard, J.I.; Zhang, L.; Smith, K.L.; Morel, C.R.; Greaves, D.R.; Murray, P.J.; Chawla, A. Oxidative Metabolism and PGC-1β Attenuate Macrophage-Mediated Inflammation. Cell Metab. 2006, 4, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Mirebeau-Prunier, D.; Le Pennec, S.; Jacques, C.; Gueguen, N.; Poirier, J.; Malthiery, Y.; Savagner, F. Estrogen-Related Receptor α and PGC-1-Related Coactivator Constitute a Novel Complex Mediating the Biogenesis of Functional Mitochondria. FEBS J. 2010, 277, 713–725. [Google Scholar] [CrossRef]
- Jung, S.B.; Choi, M.J.; Ryu, D.; Yi, H.S.; Lee, S.E.; Chang, J.Y.; Chung, H.K.; Kim, Y.K.; Kang, S.G.; Lee, J.H.; et al. Reduced Oxidative Capacity in Macrophages Results in Systemic Insulin Resistance. Nat. Commun. 2018, 9, 1551. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The Role of Selenium in Inflammation and Immunity: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Choi, S.I.; Lee, H.R.; Hwang, I.S.; Lee, Y.J.; An, B.S.; Lee, S.H.; Kim, H.J.; Kang, B.C.; Hwang, D.Y. Selenium Significantly Inhibits Adipocyte Hypertrophy and Abdominal Fat Accumulation in OLETF Rats via Induction of Fatty Acid β-Oxidation. Biol. Trace Elem. Res. 2012, 150, 360–370. [Google Scholar] [CrossRef]
- Pfister, C.; Dawczynski, H.; Schingale, F.J. Selenium Deficiency in Lymphedema and Lipedema—A Retrospective Cross-Sectional Study from a Specialized Clinic. Nutrients 2020, 12, 1211. [Google Scholar] [CrossRef]
- Petersen, K.J.; Garza, M.; Donahue, P.M.C.; Harkins, K.D.; Marton, A.; Titze, J.; Donahue, M.J.; Crescenzi, R. Neuroimaging of Cerebral Blood Flow and Sodium in Women with Lipedema. Obesity 2020, 28, 1292–1300. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The Role of Estrogens in Control of Energy Balance and Glucose Homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [Green Version]
- Michelini, S.; Chiurazzi, P.; Marino, V.; Dell’orco, D.; Manara, E.; Baglivo, M.; Fiorentino, A.; Maltese, P.E.; Pinelli, M.; Herbst, K.L.; et al. Aldo-Keto Reductase 1c1 (Akr1c1) as the First Mutated Gene in a Family with Nonsyndromic Primary Lipedema. Int. J. Mol. Sci. 2020, 21, 6264. [Google Scholar] [CrossRef]
- Zhang, Y.; Nadeau, M.; Faucher, F.; Lescelleur, O.; Biron, S.; Daris, M.; Rhéaume, C.; Luu-The, V.; Tchernof, A. Progesterone Metabolism in Adipose Cells. Mol. Cell. Endocrinol. 2009, 298, 76–83. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.N.; Welter, B.H.; Mantzke, K.A.; Price, T.M. Identification of Progesterone Receptor in Human Subcutaneous Adipose Tissue. J. Clin. Endocrinol. Metab. 1998, 83, 509–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.M.; Wade, G.N. Food Intake, Body Weight, and Adiposity in Female Rats: Actions and Interactions of Progestins and Antiestrogens. Am. J. Physiol. Metab. 1981, 240, E474–E481. [Google Scholar] [CrossRef]
- Lof, M.; Hilakivi-Clarke, L.; Sandin, S.S.; de Assis, S.; Yu, W.; Weiderpass, E. Dietary Fat Intake and Gestational Weight Gain in Relation to Estradiol and Progesterone Plasma Levels during Pregnancy: A Longitudinal Study in Swedish Women. BMC Womens. Health 2009, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, G.; Mansour, S.; Brice, G.; Ostergaard, P.; Mortimer, P.S.; Jeffery, S.; Nussey, S. Pit-1 Mutation and Lipoedema in a Family. Exp. Clin. Endocrinol. Diabetes 2010, 118, 377–380. [Google Scholar] [CrossRef]
- Precone, V.; Barati, S.; Paolacci, S.; Salgarello, M.; Visconti, G.; Gentileschi, S.; Guerri, G.; Gagliardi, L.; Aquilanti, B.; Matera, G.; et al. Genetic Syndromes with Localized Subcutaneous Fat Tissue Accumulation. Acta Biomed. 2019, 90, 90–92. [Google Scholar] [CrossRef]
- Span, J.P.T.; Pieters, G.F.F.M.; Sweep, C.G.J.; Hermus, A.R.M.M.; Smals, A.G.H. Gender Difference in Insulin-Like Growth Factor I Response to Growth Hormone (GH) Treatment in GH-Deficient Adults: Role of Sex Hormone Replacement. J. Clin. Endocrinol. Metab. 2000, 85, 1121–1125. [Google Scholar] [CrossRef]
- Krattenmacher, R.; Knauthe, R.; Parczyk, K.; Walker, A.; Hilgenfeldt, U.; Fritzemeier, K.H. Estrogen Action on Hepatic Synthesis of Angiotensinogen and IGF-I: Direct and Indirect Estrogen Effects. J. Steroid Biochem. Mol. Biol. 1994, 48, 207–214. [Google Scholar] [CrossRef]
- Zechner, U.; Kohlschmidt, N.; Kempf, O.; Gebauer, K.; Haug, K.; Engels, H.; Haaf, T.; Bartsch, O. Familial Sotos Syndrome Caused by a Novel Missense Mutation, C2175S, in NSD1 and Associated with Normal Intelligence, Insulin Dependent Diabetes, Bronchial Asthma, and Lipedema. Eur. J. Med. Genet. 2009, 52, 306–310. [Google Scholar] [CrossRef]
- Visser, R.; Landman, E.B.M.; Goeman, J.; Wit, J.M.; Karperien, M. Sotos Syndrome Is Associated with Deregulation of the MAPK/ERK-Signaling Pathway. PLoS ONE 2012, 7, e49229. [Google Scholar] [CrossRef]
- Huang, N.; Vom Baur, E.; Garnier, J.-M.; Lerouge, T.; Vonesch, J.-L.; Lutz, Y.; Chambon, P.; Losson, R.G. Two Distinct Nuclear Receptor Interaction Domains in NSD1, a Novel SET Protein That Exhibits Characteristics of Both Corepressors and Coactivators. EMBO J. 1998, 17, 3398–3412. [Google Scholar] [CrossRef] [Green Version]
- Waxler, J.L.; Guardino, C.; Feinn, R.S.; Lee, H.; Pober, B.R.; Stanley, T.L. Altered Body Composition, Lipedema, and Decreased Bone Density in Individuals with Williams Syndrome: A Preliminary Report. Eur. J. Med. Genet. 2017, 60, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Michelini, S.; Herbst, K.L.; Precone, V.; Manara, E.; Marceddu, G.; Dautaj, A.; Maltese, P.E.; Paolacci, S.; Ceccarini, M.R.; Beccari, T.; et al. A Multi-Gene Panel to Identify Lipedema-Predisposing Genetic Variants by a Next-Generation Sequencing Strategy. J. Pers. Med. 2022, 12, 268. [Google Scholar] [CrossRef]
- Grigoriadis, D.; Sackey, E.; Riches, K.; van Zanten, M.; Brice, G.; England, R.; Mills, M.; Dobbins, S.E.; Lee, L.L.; Jeffery, S.; et al. Investigation of Clinical Characteristics and Genome Associations in the ‘UK Lipoedema’ Cohort. PLoS ONE 2022, 17, e0274867. [Google Scholar] [CrossRef]
- Eder, K.; Baffy, N.; Falus, A.; Fulop, A.K. The Major Inflammatory Mediator Interleukin-6 and Obesity. Inflamm. Res. 2009, 58, 727–736. [Google Scholar] [CrossRef]
- El-Mikkawy, D.M.E.; EL-Sadek, M.A.; EL-Badawy, M.A.; Samaha, D. Circulating Level of Interleukin-6 in Relation to Body Mass Indices and Lipid Profile in Egyptian Adults with Overweight and Obesity. Egypt. Rheumatol. Rehabil. 2020, 47, 7. [Google Scholar] [CrossRef]
- Di Renzo, L.; Gualtieri, P.; Alwardat, N.; De Santis, G.; Zomparelli, S.; Romano, L.; Marchetti, M.; Michelini, S.; Capacci, A.; Piccioni, A.; et al. The Role of IL-6 Gene Polymorphisms in the Risk of Lipedema. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3236–3244. [Google Scholar] [CrossRef]
- Ricke, R.M.; Jeganathan, K.B.; van Deursen, J.M. Bub1 Overexpression Induces Aneuploidy and Tumor Formation through Aurora B Kinase Hyperactivation. J. Cell Biol. 2011, 193, 1049–1064. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.J.; Pan, Y.; Chen, X.Y.; Hou, P.F. BUB1 Promotes Proliferation of Liver Cancer Cells by Activating SMAD2 Phosphorylation. Oncol. Lett. 2020, 19, 3506. [Google Scholar] [CrossRef]
- Jiang, N.; Liao, Y.; Wang, M.; Wang, Y.; Wang, K.; Guo, J.; Wu, P.; Zhong, B.; Guo, T.; Wu, C. BUB1 Drives the Occurrence and Development of Bladder Cancer by Mediating the STAT3 Signaling Pathway. J. Exp. Clin. Cancer Res. 2021, 40, 378. [Google Scholar] [CrossRef]
- Zhao, M.L.; Rabiee, A.; Kovary, K.M.; Bahrami-Nejad, Z.; Taylor, B.; Teruel, M.N. Molecular Competition in G1 Controls When Cells Simultaneously Commit to Terminally Differentiate and Exit the Cell Cycle. Cell Rep. 2020, 31, 107769. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Gerin, I.; Miao, H.; Vu-Phan, D.; Johnson, C.N.; Xu, R.; Chen, X.W.; Cawthorn, W.P.; MacDougald, O.A.; Koenig, R.J. Multiple Roles for the Non-Coding RNA SRA in Regulation of Adipogenesis and Insulin Sensitivity. PLoS ONE 2010, 5, e14199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Gil, H.J.; Escobedo, N.; Benito-Martín, A.; Ximénez-Embún, P.; Muñoz, J.; Peinado, H.; Rockson, S.G.; Oliver, G. Platelet Factor 4 Is a Biomarker for Lymphatic-Promoted Disorders. JCI Insight 2020, 5, e135109. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, Z.; Zhu, K.; Bao, R.; Meng, Y.; Bian, J.; Wan, X.; Yang, T. Effects of CXCL4/CXCR3 on the Lipopolysaccharide-Induced Injury in Human Umbilical Vein Endothelial Cells. J. Cell. Physiol. 2019, 234, 22378–22385. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.; Meiser, A.; McDonagh, E.M.; Fox, J.M.; Petit, S.J.; Xanthou, G.; Williams, T.J.; Pease, J.E. CXCL4-Induced Migration of Activated T Lymphocytes Is Mediated by the Chemokine Receptor CXCR3. J. Leukoc. Biol. 2008, 83, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Avraham, T.; Zampell, J.C.; Yan, A.; Elhadad, S.; Weitman, E.S.; Rockson, S.G.; Bromberg, J.; Mehrara, B.J. Th2 Differentiation Is Necessary for Soft Tissue Fibrosis and Lymphatic Dysfunction Resulting from Lymphedema. FASEB J. 2013, 27, 1114–1126. [Google Scholar] [CrossRef] [Green Version]
- Swartz, M.A. The Physiology of the Lymphatic System. Adv. Drug Deliv. Rev. 2001, 50, 3–20. [Google Scholar] [CrossRef]
- Nijs, J.; Leysen, L.; Adriaenssens, N.; Aguilar Ferrándiz, M.E.; Devoogdt, N.; Tassenoy, A.; Ickmans, K.; Goubert, D.; van Wilgen, C.P.; Wijma, A.J.; et al. Pain Following Cancer Treatment: Guidelines for the Clinical Classification of Predominant Neuropathic, Nociceptive and Central Sensitization Pain. Acta Oncol. 2016, 55, 659–663. [Google Scholar] [CrossRef] [Green Version]
- Karastergiou, K.; Mohamed-Ali, V. The Autocrine and Paracrine Roles of Adipokines. Mol. Cell. Endocrinol. 2010, 318, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Scheja, L.; Heeren, J. The Endocrine Function of Adipose Tissues in Health and Cardiometabolic Disease. Nat. Rev. Endocrinol. 2019, 15, 507–524. [Google Scholar] [CrossRef]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 Complexes Carry a Population of Circulating MicroRNAs Independent of Vesicles in Human Plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [Green Version]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of Extracellular Circulating MicroRNA. Nucleic Acids Res. 2011, 39, 7223. [Google Scholar] [CrossRef]
- Li, M.; Marin-Muller, C.; Bharadwaj, U.; Chow, K.H.; Yao, Q.; Chen, C. MicroRNAs: Control and Loss of Control in Human Physiology and Disease. World J. Surg. 2009, 33, 667–684. [Google Scholar] [CrossRef]
- Samanta, S.; Rajasingh, S.; Drosos, N.; Zhou, Z.; Dawn, B.; Rajasingh, J. Exosomes: New Molecular Targets of Diseases. Acta Pharmacol. Sin. 2017, 39, 501–513. [Google Scholar] [CrossRef]
- Malloci, M.; Perdomo, L.; Veerasamy, M.; Andriantsitohaina, R.; Simard, G.; Martínez, M.C. Extracellular Vesicles: Mechanisms in Human Health and Disease. Acta Pharmacol. Sin. 2019, 30, 813–856. [Google Scholar] [CrossRef]
- Rupp, A.K.; Rupp, C.; Keller, S.; Brase, J.C.; Ehehalt, R.; Fogel, M.; Moldenhauer, G.; Marmé, F.; Sültmann, H.; Altevogt, P. Loss of EpCAM Expression in Breast Cancer Derived Serum Exosomes: Role of Proteolytic Cleavage. Gynecol. Oncol. 2011, 122, 437–446. [Google Scholar] [CrossRef]
- Liang, B.; Peng, P.; Chen, S.; Li, L.; Zhang, M.; Cao, D.; Yang, J.; Li, H.; Gui, T.; Li, X.; et al. Characterization and Proteomic Analysis of Ovarian Cancer-Derived Exosomes. J. Proteom. 2013, 80, 171–182. [Google Scholar] [CrossRef]
- Park, Y.H.; Shin, H.W.; Jung, A.R.; Kwon, O.S.; Choi, Y.J.; Park, J.; Lee, J.Y. Prostate-Specific Extracellular Vesicles as a Novel Biomarker in Human Prostate Cancer. Sci. Rep. 2016, 6, 30386. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Shen, A.; Guo, F.; Song, Y.; Jing, N.; Ding, X.; Pan, M.; Zhang, H.; Wang, J.; Wu, L.; et al. Urinary Exosomal MiRNA-4534 as a Novel Diagnostic Biomarker for Diabetic Kidney Disease. Front. Endocrinol. 2020, 11, 590. [Google Scholar] [CrossRef]
- Lin, J.; Li, J.; Huang, B.; Liu, J.; Chen, X.; Chen, X.M.; Xu, Y.M.; Huang, L.F.; Wang, X.Z. Exosomes: Novel Biomarkers for Clinical Diagnosis. Sci. World J. 2015, 2015, 657086. [Google Scholar] [CrossRef] [Green Version]
- Van Balkom, B.W.M.; Eisele, A.S.; Michiel Pegtel, D.; Bervoets, S.; Verhaar, M.C. Quantitative and Qualitative Analysis of Small RNAs in Human Endothelial Cells and Exosomes Provides Insights into Localized RNA Processing, Degradation and Sorting. J. Extracell. Vesicles 2015, 4, 26760. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tani, S.; Yagi, T.; Atsumi, W.; Kawauchi, K.; Matsuo, R.; Hirayama, A. Relation between Low-Density Lipoprotein Cholesterol/Apolipoprotein B Ratio and Triglyceride-Rich Lipoproteins in Patients with Coronary Artery Disease and Type 2 Diabetes Mellitus: A Cross-Sectional Study. Cardiovasc. Diabetol. 2017, 16, 123. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Chiu, C.J.; Kuan, C.H.; Chen, F.H.; Shen, Y.C.; Wu, C.H.; Hsu, Y.H. IL-29 Promoted Obesity-Induced Inflammation and Insulin Resistance. Cell. Mol. Immunol. 2020, 17, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, M.P.; Pankhong, P.; Laddy, D.J.; Schoenly, K.A.; Yan, J.; Cisper, N.; Weiner, D.B. Comparative Ability of IL-12 and IL-28B to Regulate Treg Populations and Enhance Adaptive Cellular Immunity. Blood 2009, 113, 5868–5877. [Google Scholar] [CrossRef] [Green Version]
- Keller, D.; Du, X.; Srour, E.; Hoffman, R.; Williams, D. Interleukin-11 Inhibits Adipogenesis and Stimulates Myelopoiesis in Human Long-Term Marrow Cultures. Blood 1993, 82, 1428–1435. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zhang, S.; Ou, T.; Jiang, H.; Jia, D.; Qi, Z.; Zou, Y.; Qian, J.; Sun, A.; Ge, J. Interleukin-11 Regulates the Fate of Adipose-Derived Mesenchymal Stem Cells via STAT3 Signalling Pathways. Cell Prolif. 2020, 53, e12771. [Google Scholar] [CrossRef]
- Kim, S.-J.; Park, J.; Byun, H.; Park, Y.-W.; Major, L.G.; Lee, D.Y.; Choi, Y.S.; Shin, H. Hydrogels with an Embossed Surface: An All-in-One Platform for Mass Production and Culture of Human Adipose-Derived Stem Cell Spheroids. Biomaterials 2019, 188, 198–212. [Google Scholar] [CrossRef]
- Compera, N.; Atwell, S.; Wirth, J.; von Törne, C.; Hauck, S.M.; Meier, M. Adipose Microtissue-on-Chip: A 3D Cell Culture Platform for Differentiation, Stimulation, and Proteomic Analysis of Human Adipocytes. Lab Chip 2022, 22, 3172–3186. [Google Scholar] [CrossRef]
- Zheng, G.X.Y.; Terry, J.M.; Belgrader, P.; Ryvkin, P.; Bent, Z.W.; Wilson, R.; Ziraldo, S.B.; Wheeler, T.D.; McDermott, G.P.; Zhu, J.; et al. Massively Parallel Digital Transcriptional Profiling of Single Cells. Nat. Commun. 2017, 8, 14049. [Google Scholar] [CrossRef] [Green Version]
- Rajbhandari, P.; Arneson, D.; Hart, S.K.; Ahn, I.S.; Diamante, G.; Santos, L.C.; Zaghari, N.; Feng, A.C.; Thomas, B.J.; Vergnes, L.; et al. Single Cell Analysis Reveals Immune Cell-Adipocyte Crosstalk Regulating the Transcription of Thermogenic Adipocytes. Elife 2019, 8, e49501. [Google Scholar] [CrossRef]
- Jiang, Y.; Yeung, J.L.H.; Lee, J.H.; An, J.; Steadman, P.E.; Kim, J.R.; Sung, H.K. Visualization of 3D White Adipose Tissue Structure Using Whole-Mount Staining. J. Vis. Exp. 2018, 2018, e58683. [Google Scholar] [CrossRef] [Green Version]
- Hansmeier, N.R.; Büschlen, I.S.; Behncke, R.Y.; Ulferts, S.; Bisoendial, R.; Hägerling, R. 3D Visualization of Human Blood Vascular Networks Using Single-Domain Antibodies Directed against Endothelial Cell-Selective Adhesion Molecule (ESAM). Int. J. Mol. Sci. 2022, 23, 4369. [Google Scholar] [CrossRef]
- Zhao, R.; Kaakati, R.; Liu, X.; Xu, L.; Lee, A.K.; Bachelder, R.; Li, C.Y.; Hollenbeck, S.T. CRISPR/Cas9-Mediated BRCA1 Knockdown Adipose Stem Cells Promote Breast Cancer Progression. Plast. Reconstr. Surg. 2019, 143, 747. [Google Scholar] [CrossRef]
- Tsagkaraki, E.; Nicoloro, S.M.; DeSouza, T.; Solivan-Rivera, J.; Desai, A.; Lifshitz, L.M.; Shen, Y.; Kelly, M.; Guilherme, A.; Henriques, F.; et al. CRISPR-Enhanced Human Adipocyte Browning as Cell Therapy for Metabolic Disease. Nat. Commun. 2021, 12, 6931. [Google Scholar] [CrossRef]
- Hill, J.L.; McIver, K.B.; Katzer, K.; Foster, M.T. Capillary Western Immunoassay Optimization of Estrogen Related Factors in Human Subcutaneous Adipose Tissue. Methods Protoc. 2022, 5, 34. [Google Scholar] [CrossRef]
- Birkballe, S.; Jensen, M.R.; Noerregaard, S.; Gottrup, F.; Karlsmark, T. Can Tissue Dielectric Constant Measurement Aid in Differentiating Lymphoedema from Lipoedema in Women with Swollen Legs? Br. J. Dermatol. 2014, 170, 96–102. [Google Scholar] [CrossRef]
- Peled, A.W.; Kappos, E.A. Lipedema: Diagnostic and Management Challenges. Int. J. Womens Health 2016, 8, 389. [Google Scholar] [CrossRef] [Green Version]
- Hodge, L.M.; King, H.H.; Williams, A.G.; Reder, S.J.; Belavadi, T.; Simecka, J.W.; Stoll, S.T.; Downey, H.F. Abdominal Lymphatic Pump Treatment Increases Leukocyte Count and Flux in Thoracic Duct Lymph. Lymphat. Res. Biol. 2007, 5, 127–133. [Google Scholar] [CrossRef]
- Moeller, J.A.; Cleveland, S.D.; Schaverien, M.V. Lymphedema Management: Nonoperative Treatment of Lymphedema. Semin. Plast. Surg. 2018, 32, 17. [Google Scholar] [CrossRef]
- Wilburn, O.; Wilburn, P.; Rockson, S.G. A Pilot, Prospective Evaluation of a Novel Alternative for Maintenance Therapy of Breast Cancer-Associated Lymphedema [ISRCTN76522412]. BMC Cancer 2006, 6, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellini, E.; Grieco, M.P.; Raposio, E. A Journey through Liposuction and Liposculture: Review. Ann. Med. Surg. 2017, 24, 53. [Google Scholar] [CrossRef] [PubMed]
- Szél, E.; Kemény, L.; Groma, G.; Szolnoky, G. Pathophysiological Dilemmas of Lipedema. Med. Hypotheses 2014, 83, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Czerwińska, M.; Ostrowska, P.; Hansdorfer-Korzon, R. Lipoedema as a Social Problem. A Scoping Review. Int. J. Environ. Res. Public Health 2021, 18, 10223. [Google Scholar] [CrossRef]
- Goodliffe, J.M.; Ormerod, J.O.M.; Beale, A.; Ramcharitar, S. An Under-Diagnosed Cause of Leg Swelling. BMJ Case Rep. 2013, 2013, bcr2013009538. [Google Scholar] [CrossRef] [Green Version]
- Barré-Sinoussi, F.; Montagutelli, X. Animal Models Are Essential to Biological Research: Issues and Perspectives. Futur. Sci. OA 2015, 1, FSO63. [Google Scholar] [CrossRef] [Green Version]
- Warodomwichit, D.; Shen, J.; Arnett, D.K.; Tsai, M.Y.; Kabagambe, E.K.; Peacock, J.M.; Hixson, J.E.; Straka, R.J.; Province, M.A.; An, P.; et al. The Monounsaturated Fatty Acid Intake Modulates the Effect of ADIPOQ Polymorphisms on Obesity. Obesity 2009, 17, 510. [Google Scholar] [CrossRef] [Green Version]
- Siitonen, N.; Pulkkinen, L.; Lindström, J.; Kolehmainen, M.; Eriksson, J.G.; Venojärvi, M.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Tuomilehto, J.; Uusitupa, M. Association of ADIPOQ Gene Variants with Body Weight, Type 2 Diabetes and Serum Adiponectin Concentrations: The Finnish Diabetes Prevention Study. BMC Med. Genet. 2011, 12, 5. [Google Scholar] [CrossRef]
- Zhao, S.; Li, N.; Zhu, Y.; Straub, L.; Zhang, Z.; Wang, M.Y.; Zhu, Q.; Kusminski, C.M.; Elmquist, J.K.; Scherer, P.E. Partial Leptin Deficiency Confers Resistance to Diet-Induced Obesity in Mice. Mol. Metab. 2020, 37, 100995. [Google Scholar] [CrossRef]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. 2021, 12, 563. [Google Scholar] [CrossRef]
- Song, N.-J.; Kim, S.; Jang, B.-H.; Chang, S.-H.; Yun, U.J.; Park, K.-M.; Waki, H.; Li, D.Y.; Tontonoz, P.; Park, K.W. Small Molecule-Induced Complement Factor D (Adipsin) Promotes Lipid Accumulation and Adipocyte Differentiation. PLoS ONE 2016, 11, e0162228. [Google Scholar] [CrossRef] [Green Version]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J.; Liu, X.S.; et al. PPARγ and C/EBP Factors Orchestrate Adipocyte Biology via Adjacent Binding on a Genome-Wide Scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Barak, Y.; Hevener, A.; Olson, P.; Liao, D.; Le, J.; Nelson, M.; Ong, E.; Olefsky, J.M.; Evans, R.M. Adipose-Specific Peroxisome Proliferator-Activated Receptor γ Knockout Causes Insulin Resistance in Fat and Liver but Not in Muscle. Proc. Natl. Acad. Sci. USA 2003, 100, 15712–15717. [Google Scholar] [CrossRef]
- Hishida, T.; Nishizuka, M.; Osada, S.; Imagawa, M. The Role of C/EBPdelta in the Early Stages of Adipogenesis. Biochimie 2009, 91, 654–657. [Google Scholar] [CrossRef]
- Min, Y.; Ghose, S.; Boelte, K.; Li, J.; Yang, L.; Lin, P.C. C/EBP-δ Regulates VEGF-C Autocrine Signaling in Lymphangiogenesis and Metastasis of Lung Cancer through HIF-1α. Oncogene 2011, 30, 4901–4909. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Sharma, N.; Kapadia, F.; Zhou, G.; Lu, Y.; Hong, H.; Paruchuri, K.; Mahabeleshwar, G.H.; Dalmas, E.; Venteclef, N.; et al. Krüppel-like Factor 4 Regulates Macrophage Polarization. J. Clin. Investig. 2011, 121, 2736–2749. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Markan, K.; Temple, K.A.; Deplewski, D.; Brady, M.J.; Cohen, R.N. The Nuclear Receptor Corepressors NCoR and SMRT Decrease Peroxisome Proliferator-Activated Receptor Gamma Transcriptional Activity and Repress 3T3-L1 Adipogenesis. J. Biol. Chem. 2005, 280, 13600–13605. [Google Scholar] [CrossRef] [Green Version]
- Bond, H.M.; Scicchitano, S.; Chiarella, E.; Amodio, N.; Lucchino, V.; Aloisio, A.; Montalcini, Y.; Mesuraca, M.; Morrone, G. ZNF423: A New Player in Estrogen Receptor-Positive Breast Cancer. Front. Endocrinol. 2018, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Cignarelli, A.; Genchi, V.A.; Perrini, S.; Natalicchio, A.; Laviola, L.; Giorgino, F. Insulin and Insulin Receptors in Adipose Tissue Development. Int. J. Mol. Sci. 2019, 20, 759. [Google Scholar] [CrossRef] [Green Version]
- Graham, T.E.; Yang, Q.; Blüher, M.; Hammarstedt, A.; Ciaraldi, T.P.; Henry, R.R.; Wason, C.J.; Oberbach, A.; Jansson, P.-A.; Smith, U.; et al. Retinol-Binding Protein 4 and Insulin Resistance in Lean, Obese, and Diabetic Subjects. N. Engl. J. Med. 2006, 354, 2552–2563. [Google Scholar] [CrossRef]
- Pant, R.; Alam, A.; Choksi, A.; Shah, V.K.; Firmal, P.; Chattopadhyay, S. Chromatin Remodeling Protein SMAR1 Regulates Adipogenesis by Modulating the Expression of PPARγ. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2021, 1866, 159045. [Google Scholar] [CrossRef] [PubMed]
- Pooley, R.D.; Moynihan, K.L.; Soukoulis, V.; Reddy, S.; Francis, R.; Lo, C.; Ma, L.J.; Bader, D.M. Murine CENPF Interacts with Syntaxin 4 in the Regulation of Vesicular Transport. J. Cell Sci. 2008, 121, 3413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, L.; Zhang, X.; Deng, Y.; Han, J.; Yang, J.; Chen, S.; Fang, Q.; Yang, Y.; Jia, W. Enhanced Expression of Survivin Has Distinct Roles in Adipocyte Homeostasis. Cell Death Dis. 2017, 8, e2533. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Hinkle, J.; Reed, J.N.; Aherrahrou, R.; Xu, Z.; Harris, T.E.; Stephenson, E.J.; Musunuru, K.; Keller, S.R.; Civelek, M. Adipocyte-Specific Modulation of KLF14 Expression in Mice Leads to Sex-Dependent Impacts on Adiposity and Lipid Metabolism. Diabetes 2022, 71, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; Weigelt, C.; Cherradi, M.L.; Niemeier, A.; Tödter, K.; Heeren, J.; Scheja, L. Effects of Adipocyte Lipoprotein Lipase on de Novo Lipogenesis and White Adipose Tissue Browning. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2013, 1831, 934–942. [Google Scholar] [CrossRef]
- Garin-Shkolnik, T.; Rudich, A.; Hotamisligil, G.S.; Rubinstein, M. FABP4 Attenuates PPARγ and Adipogenesis and Is Inversely Correlated With PPARγ in Adipose Tissues. Diabetes 2014, 63, 900–911. [Google Scholar] [CrossRef] [Green Version]
- Karaman, S.; Hollmén, M.; Robciuc, M.R.; Alitalo, A.; Nurmi, H.; Morf, B.; Buschle, D.; Alkan, H.F.; Ochsenbein, A.M.; Alitalo, K.; et al. Blockade of VEGF-C and VEGF-D Modulates Adipose Tissue Inflammation and Improves Metabolic Parameters under High-Fat Diet. Mol. Metab. 2015, 4, 93–105. [Google Scholar] [CrossRef]
- Su, J.L.; Yen, C.J.; Chen, P.S.; Chuang, S.E.; Hong, C.C.; Kuo, I.H.; Chen, H.Y.; Hung, M.C.; Kuo, M.L. The Role of the VEGF-C/VEGFR-3 Axis in Cancer Progression. Br. J. Cancer 2007, 96, 541–545. [Google Scholar] [CrossRef]
- Hong, Y.K.; Harvey, N.; Noh, Y.H.; Schacht, V.; Hirakawa, S.; Detmar, M.; Oliver, G. Prox1 Is a Master Control Gene in the Program Specifying Lymphatic Endothelial Cell Fate. Dev. Dyn. 2002, 225, 351–357. [Google Scholar] [CrossRef]
- Schacht, V.; Ramirez, M.I.; Hong, Y.K.; Hirakawa, S.; Feng, D.; Harvey, N.; Williams, M.; Dvorak, A.M.; Dvorak, H.F.; Oliver, G.; et al. T1α/Podoplanin Deficiency Disrupts Normal Lymphatic Vasculature Formation and Causes Lymphedema. EMBO J. 2003, 22, 3546. [Google Scholar] [CrossRef]
- Wigle, J.T.; Harvey, N.; Detmar, M.; Lagutina, I.; Grosveld, G.; Gunn, M.D.; Jackson, D.G.; Oliver, G. An Essential Role for Prox1 in the Induction of the Lymphatic Endothelial Cell Phenotype. EMBO J. 2002, 21, 1505. [Google Scholar] [CrossRef]
- Mishima, K.; Watabe, T.; Saito, A.; Yoshimatsu, Y.; Imaizumi, N.; Masui, S.; Hirashima, M.; Morisada, T.; Oike, Y.; Araie, M.; et al. Prox1 Induces Lymphatic Endothelial Differentiation via Integrin A9 and Other Signaling Cascades. Mol. Biol. Cell 2007, 18, 1421–1429. [Google Scholar] [CrossRef]
- Jackson, D.G.; Biology, J.D. Biology of the Lymphatic Marker LYVE-1 and Applications in Research into Lymphatic Trafficking and Lymphangiogenesis. APMIS 2004, 112, 526–538. [Google Scholar] [CrossRef]
- Cho, C.H.; Koh, Y.J.; Han, J.; Sung, H.K.; Lee, H.J.; Morisada, T.; Schwendener, R.A.; Brekken, R.A.; Kang, G.; Oike, Y.; et al. Angiogenic Role of LYVE-1–Positive Macrophages in Adipose Tissue. Circ. Res. 2007, 100, e47–e57. [Google Scholar] [CrossRef] [Green Version]
- Vaahtomeri, K.; Moussion, C.; Hauschild, R.; Sixt, M. Shape and Function of Interstitial Chemokine CCL21 Gradients Are Independent of Heparan Sulfates Produced by Lymphatic Endothelium. Front. Immunol. 2021, 12, 249. [Google Scholar] [CrossRef]
- Sauteur, L.; Krudewig, A.; Herwig, L.; Ehrenfeuchter, N.; Lenard, A.; Affolter, M.; Belting, H.G. Cdh5/VE-Cadherin Promotes Endothelial Cell Interface Elongation via Cortical Actin Polymerization during Angiogenic Sprouting. Cell Rep. 2014, 9, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Fukuhara, S.; Sako, K.; Noda, K.; Nagao, K.; Miura, K.; Mochizuki, N. Tie2 Is Tied at the Cell-Cell Contacts and to Extracellular Matrix by Angiopoietin-1. Exp. Mol. Med. 2009, 41, 133–139. [Google Scholar] [CrossRef]
- Leppänen, V.M.; Brouillard, P.; Korhonen, E.A.; Sipilä, T.; Kumar, S.; Revencu, N.; Labarque, V.; Fastrè, E.; Schlögel, M.; Ravoet, M.; et al. Characterization of ANGPT2 Mutations Associated with Primary Lymphedema. Sci. Transl. Med. 2020, 12, eaax8013. [Google Scholar] [CrossRef]
- Fedorenko, A.; Lishko, P.V.; Kirichok, Y. Mechanism of Fatty-Acid-Dependent UCP1 Uncoupling in Brown Fat Mitochondria. Cell 2012, 151, 400. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, S.; Oeckl, J.; Klingenspor, M.; Fromme, T. Substrate Fluxes in Brown Adipocytes upon Adrenergic Stimulation and Uncoupling Protein 1 Ablation. Life Sci. Alliance 2018, 1, e201800136. [Google Scholar] [CrossRef]
- Rabiee, A. Beige Fat Maintenance; Toward a Sustained Metabolic Health. Front. Endocrinol. 2020, 11, 634. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Perrard, X.D.; Wang, Q.; Perrard, J.L.; Polsani, V.R.; Jones, P.H.; Smith, C.W.; Ballantyne, C.M. CD11c Expression in Adipose Tissue and Blood and Its Role in Diet-Induced Obesity. Arter. Thromb. Vasc. Biol. 2010, 30, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, L.; Shu, G.; Wang, B. MicroRNA-16–5p Promotes 3T3-L1 Adipocyte Differentiation through Regulating EPT1. Biochem. Biophys. Res. Commun. 2019, 514, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Xu, Z.; Ou, D.; Liu, J.; Zhang, J. The MiR-15a/16 Gene Cluster in Human Cancer: A Systematic Review. J. Cell. Physiol. 2019, 234, 5496–5506. [Google Scholar] [CrossRef]
- Kristensen, M.M.; Davidsen, P.K.; Vigelsø, A.; Hansen, C.N.; Jensen, L.J.; Jessen, N.; Bruun, J.M.; Dela, F.; Helge, J.W. MiRNAs in Human Subcutaneous Adipose Tissue: Effects of Weight Loss Induced by Hypocaloric Diet and Exercise. Obesity 2017, 25, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Barros-Oliveira, M.d.C.; Costa-Silva, D.R.; Dos Santos, A.R.; Pereira, R.O.; Soares-Júnior, J.M.; da Silva, B.B. Influence of CYP19A1 Gene Expression Levels in Women with Breast Cancer: A Systematic Review of the Literature. Clinics 2021, 76, e2846. [Google Scholar] [CrossRef]




| Stage I | Stage II | Stage III | Stage IV |
|---|---|---|---|
|
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poojari, A.; Dev, K.; Rabiee, A. Lipedema: Insights into Morphology, Pathophysiology, and Challenges. Biomedicines 2022, 10, 3081. https://doi.org/10.3390/biomedicines10123081
Poojari A, Dev K, Rabiee A. Lipedema: Insights into Morphology, Pathophysiology, and Challenges. Biomedicines. 2022; 10(12):3081. https://doi.org/10.3390/biomedicines10123081
Chicago/Turabian StylePoojari, Ankita, Kapil Dev, and Atefeh Rabiee. 2022. "Lipedema: Insights into Morphology, Pathophysiology, and Challenges" Biomedicines 10, no. 12: 3081. https://doi.org/10.3390/biomedicines10123081
APA StylePoojari, A., Dev, K., & Rabiee, A. (2022). Lipedema: Insights into Morphology, Pathophysiology, and Challenges. Biomedicines, 10(12), 3081. https://doi.org/10.3390/biomedicines10123081