
Epstein-Barr Virus Load Correlates with Multiple Sclerosis-Associated Retrovirus Envelope Expression

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Variables of the Study
- Laboratory variables: titers of antibodies against EBV (anti-VCA (EBV viral capsid antigen), anti-EBNA-1 (Epstein-Barr nuclear antigen) IgG antibodies), and HHV-6A/B (IgG and IgM antibodies); pHERV-W ENV/syncytin-1 protein expression; pHERV-W ENV and syncytin-1 gene expression; EBV and HHV-6A/B viral loads; MS-related class II HLA alleles genotype.
- Demographic variables: sex and age.
- Clinical variables: age of onset of MS, disease duration, treatment, disability (according to EDSS—expanded disability status scale—and MSSS—multiple sclerosis severity score), presence/absence of relapses, gadolinium-enhanced magnetic resonance imaging (MRI) lesions presence/absence, and progression of the disease after 12 months (defined depending on the baseline EDSS score: increase ≥1.5 points if baseline EDSS was 0; increase ≥1 point if baseline EDSS was ≥1 and ≤5; increase ≥0.5 points if baseline EDSS was ≥5.5).
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Monoclonal Antibody against pHERV-W ENV/syncytin-1 Obtention
2.5. Flow Cytometry
2.6. Nucleic Acids Isolation
2.7. qPCR for pHERV-W ENV and syncytin-1 Gene Expression Quantification
2.8. qPCR for EBV and HHV-6A/B Viral Loads Determination
2.9. MS-Related HLA Class-II Alleles Genotyping
2.10. Statistics
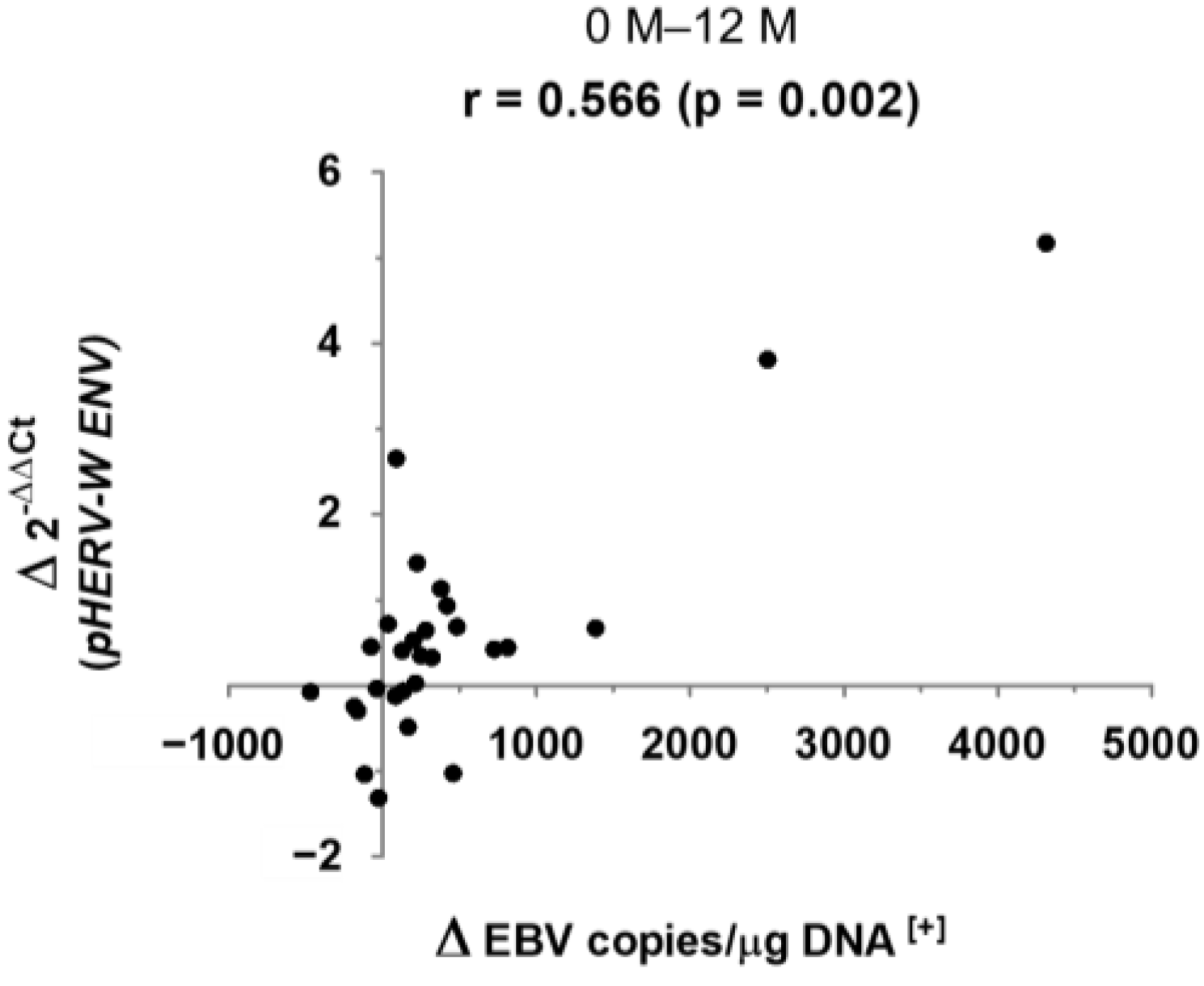
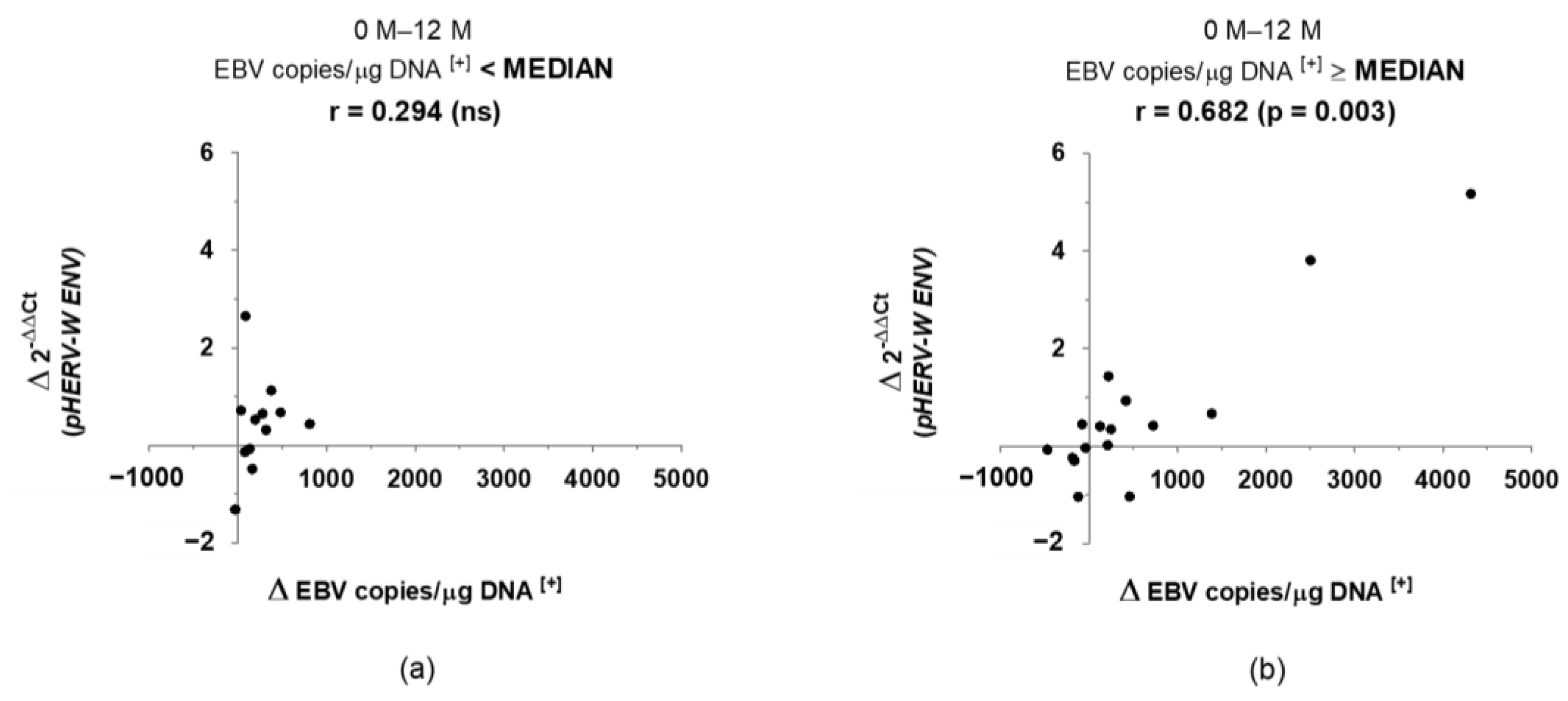
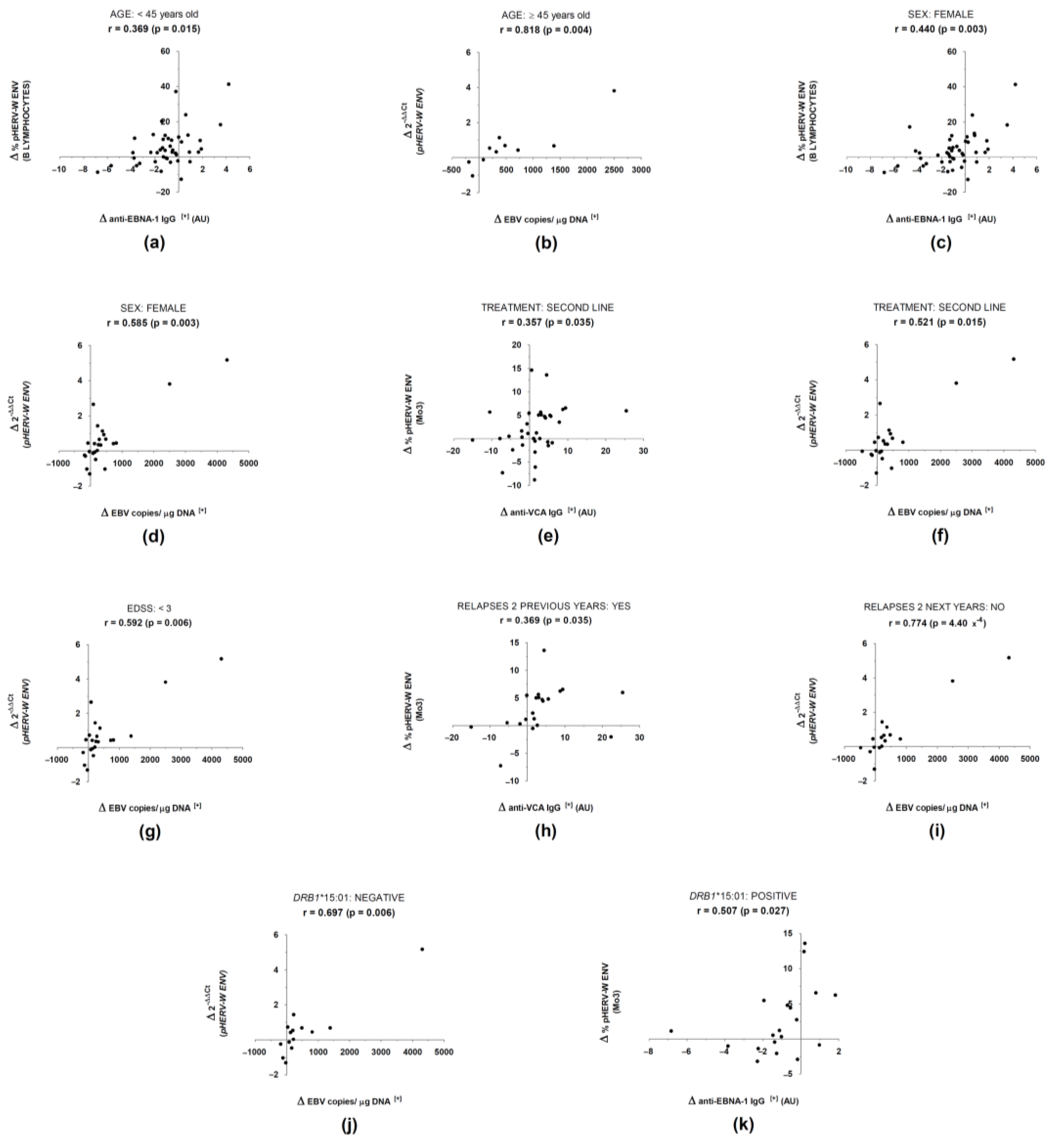
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of Multiple Sclerosis: 2017 Revisions of the McDonald Criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- International Multiple Sclerosis Genetics Consortium; Wellcome Trust Case Control Consortium 2; Sawcer, S.; Hellenthal, G.; Pirinen, M.; Spencer, C.C.A.; Patsopoulos, N.A.; Moutsianas, L.; Dilthey, A.; Su, Z.; et al. Genetic Risk and a Primary Role for Cell-Mediated Immune Mechanisms in Multiple Sclerosis. Nature 2011, 476, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Moutsianas, L.; Jostins, L.; Beecham, A.H.; Dilthey, A.T.; Xifara, D.K.; Ban, M.; Shah, T.S.; Patsopoulos, N.A.; Alfredsson, L.; Anderson, C.A.; et al. Class II HLA Interactions Modulate Genetic Risk for Multiple Sclerosis. Nat. Genet. 2015, 47, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Dobson, R.; Giovannoni, G. Multiple Sclerosis–A Review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrodan, M.; Alessandro, L.; Farez, M.F.; Correale, J. The Role of Infections in Multiple Sclerosis. Mult. Scler. Houndmills Basingstoke Engl. 2019, 25, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Bjornevik, K.; Cortese, M.; Healy, B.C.; Kuhle, J.; Mina, M.J.; Leng, Y.; Elledge, S.J.; Niebuhr, D.W.; Scher, A.I.; Munger, K.L.; et al. Longitudinal Analysis Reveals High Prevalence of Epstein-Barr Virus Associated with Multiple Sclerosis. Science 2022, 375, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Römer, C. Viruses and Endogenous Retroviruses as Roots for Neuroinflammation and Neurodegenerative Diseases. Front. Neurosci. 2021, 15, 648629. [Google Scholar] [CrossRef]
- Jakhmola, S.; Upadhyay, A.; Jain, K.; Mishra, A.; Jha, H.C. Herpesviruses and the Hidden Links to Multiple Sclerosis Neuropathology. J. Neuroimmunol. 2021, 358, 577636. [Google Scholar] [CrossRef]
- Küry, P.; Nath, A.; Créange, A.; Dolei, A.; Marche, P.; Gold, J.; Giovannoni, G.; Hartung, H.-P.; Perron, H. Human Endogenous Retroviruses in Neurological Diseases. Trends Mol. Med. 2018, 24, 379–394. [Google Scholar] [CrossRef] [Green Version]
- Dolei, A. MSRV/HERV-W/Syncytin and Its Linkage to Multiple Sclerosis: The Usability and the Hazard of a Human Endogenous Retrovirus. J. Neurovirol. 2005, 11, 232–235. [Google Scholar] [CrossRef]
- Antony, J.M.; Deslauriers, A.M.; Bhat, R.K.; Ellestad, K.K.; Power, C. Human Endogenous Retroviruses and Multiple Sclerosis: Innocent Bystanders or Disease Determinants? Biochim. Biophys. Acta 2011, 1812, 162–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mameli, G.; Poddighe, L.; Astone, V.; Delogu, G.; Arru, G.; Sotgiu, S.; Serra, C.; Dolei, A. Novel Reliable Real-Time PCR for Differential Detection of MSRVenv and Syncytin-1 in RNA and DNA from Patients with Multiple Sclerosis. J. Virol. Methods 2009, 161, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Dolei, A.; Perron, H. The Multiple Sclerosis-Associated Retrovirus and Its HERV-W Endogenous Family: A Biological Interface between Virology, Genetics, and Immunology in Human Physiology and Disease. J. Neurovirol. 2009, 15, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic Criteria for Multiple Sclerosis: 2010 Revisions to the McDonald Criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Protocols: Monoclonal Antibody Production Process. EuroMAbNet. Available online: https://www.euromabnet.com/protocols/ (accessed on 17 November 2021).
- Spanish National Cancer Research Centre, CNIO (Monoclonal Antibodies Unit) Syncytin1 CALI222B Validation File. Available online: https://www.cnio.es/downloads/investigacion-e-innovacion/servicios/anticuerpos-monoclonales/validation-files/Syncytin1-CALI222B-Validation-File.pdf (accessed on 15 November 2021).
- Pérez-Pérez, S.; Domínguez-Mozo, M.I.; García-Martínez, M.Á.; García-Frontini, M.C.; Villarrubia, N.; Costa-Frossard, L.; Villar, L.M.; Arroyo, R.; Álvarez-Lafuente, R. Anti-Human Herpesvirus 6 A/B Antibodies Titers Correlate With Multiple Sclerosis-Associated Retrovirus Envelope Expression. Front. Immunol. 2021, 12, 5098. [Google Scholar] [CrossRef]
- Pérez-Pérez, S.; Domínguez-Mozo, M.I.; García-Martínez, M.Á.; Aladro, Y.; Martínez-Ginés, M.; García-Domínguez, J.M.; López de Silanes, C.; Casanova, I.; Ortega-Madueño, I.; López-Lozano, L.; et al. Study of the Possible Link of 25-Hydroxyvitamin D with Epstein-Barr Virus and Human Herpesvirus 6 in Patients with Multiple Sclerosis. Eur. J. Neurol. 2018, 25, 1446–1453. [Google Scholar] [CrossRef]
- Picón, C.; Tejeda-Velarde, A.; Fernández-Velasco, J.I.; Comabella, M.; Álvarez-Lafuente, R.; Quintana, E.; Sainz de la Maza, S.; Monreal, E.; Villarrubia, N.; Álvarez-Cermeño, J.C.; et al. Identification of the Immunological Changes Appearing in the CSF During the Early Immunosenescence Process Occurring in Multiple Sclerosis. Front. Immunol. 2021, 12, 2742. [Google Scholar] [CrossRef]
- Leray, E.; Yaouanq, J.; Le Page, E.; Coustans, M.; Laplaud, D.; Oger, J.; Edan, G. Evidence for a Two-Stage Disability Progression in Multiple Sclerosis. Brain 2010, 133, 1900–1913. [Google Scholar] [CrossRef] [Green Version]
- Mameli, G.; Cocco, E.; Frau, J.; Marrosu, M.G.; Sechi, L.A. Epstein Barr Virus and Mycobacterium Avium Subsp. Paratuberculosis Peptides Are Recognized in Sera and Cerebrospinal Fluid of MS Patients. Sci. Rep. 2016, 6, 22401. [Google Scholar] [CrossRef]
- Mameli, G.; Cossu, D.; Cocco, E.; Masala, S.; Frau, J.; Marrosu, M.G.; Sechi, L.A. EBNA-1 IgG Titers in Sardinian Multiple Sclerosis Patients and Controls. J. Neuroimmunol. 2013, 264, 120–122. [Google Scholar] [CrossRef]
- Cossu, D.; Mameli, G.; Galleri, G.; Cocco, E.; Masala, S.; Frau, J.; Marrosu, M.G.; Manetti, R.; Sechi, L.A. Human Interferon Regulatory Factor 5 Homologous Epitopes of Epstein-Barr Virus and Mycobacterium Avium Subsp. Paratuberculosis Induce a Specific Humoral and Cellular Immune Response in Multiple Sclerosis Patients. Mult. Scler. J. 2015, 21, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Cossu, D.; Cocco, E.; Masala, S.; Frau, J.; Marrosu, M.G.; Sechi, L.A. Epstein-Barr Virus and Mycobacterium Avium Subsp. Paratuberculosis Peptides Are Cross Recognized by Anti-Myelin Basic Protein Antibodies in Multiple Sclerosis Patients. J. Neuroimmunol. 2014, 270, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Libbey, J.E.; Cusick, M.F.; Fujinami, R.S. Role of Pathogens in Multiple Sclerosis. Int. Rev. Immunol. 2014, 33, 266–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, F.C.; Lin, M.; Tai, A.; Chen, G.; Huber, B.T. Cutting Edge: Epstein-Barr Virus Transactivates the HERV-K18 Superantigen by Docking to the Human Complement Receptor 2 (CD21) on Primary B Cells. J. Immunol. 2006, 177, 2056–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perron, H.; Geny, C.; Laurent, A.; Mouriquand, C.; Pellat, J.; Perret, J.; Seigneurin, J.M. Leptomeningeal Cell Line from Multiple Sclerosis with Reverse Transcriptase Activity and Viral Particles. Res. Virol. 1989, 140, 551–561. [Google Scholar] [CrossRef]
- Perron, H.; Garson, J.A.; Bedin, F.; Beseme, F.; Paranhos-Baccala, G.; Komurian-Pradel, F.; Mallet, F.; Tuke, P.W.; Voisset, C.; Blond, J.L.; et al. Molecular Identification of a Novel Retrovirus Repeatedly Isolated from Patients with Multiple Sclerosis. Proc. Natl. Acad. Sci. USA 1997, 94, 7583–7588. [Google Scholar] [CrossRef] [Green Version]
- Perron, H.; Germi, R.; Bernard, C.; Garcia-Montojo, M.; Deluen, C.; Farinelli, L.; Faucard, R.; Veas, F.; Stefas, I.; Fabriek, B.O.; et al. Human Endogenous Retrovirus Type W Envelope Expression in Blood and Brain Cells Provides New Insights into Multiple Sclerosis Disease. Mult. Scler. Houndmills Basingstoke Engl. 2012, 18, 1721. [Google Scholar] [CrossRef]
- Arru, G.; Sechi, E.; Mariotto, S.; Farinazzo, A.; Mancinelli, C.; Alberti, D.; Ferrari, S.; Gajofatto, A.; Capra, R.; Monaco, S.; et al. Antibody Response against HERV-W Env Surface Peptides Differentiates Multiple Sclerosis and Neuromyelitis Optica Spectrum Disorder. Mult. Scler. J.-Exp. Transl. Clin. 2017, 3, 2055217317742425. [Google Scholar] [CrossRef]
- Arru, G.; Sechi, E.; Mariotto, S.; Zarbo, I.R.; Ferrari, S.; Gajofatto, A.; Monaco, S.; Deiana, G.A.; Bo, M.; Sechi, L.A.; et al. Antibody Response against HERV-W in Patients with MOG-IgG Associated Disorders, Multiple Sclerosis and NMOSD. J. Neuroimmunol. 2020, 338, 577110. [Google Scholar] [CrossRef] [Green Version]
- Mameli, G.; Madeddu, G.; Mei, A.; Uleri, E.; Poddighe, L.; Delogu, L.G.; Maida, I.; Babudieri, S.; Serra, C.; Manetti, R.; et al. Activation of MSRV-Type Endogenous Retroviruses during Infectious Mononucleosis and Epstein-Barr Virus Latency: The Missing Link with Multiple Sclerosis? PLoS ONE 2013, 8, e78474. [Google Scholar] [CrossRef]
- Bergallo, M.; Pinon, M.; Galliano, I.; Montanari, P.; Daprà, V.; Gambarino, S.; Calvo, P.L. Epstein Barr Virus Induces HERV-K and HERV-W Expression in Pediatrics Liver Transplant Recipients? Minerva Pediatr. 2020, 72, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, B.C.; Kortmann, K.; Schäfer, M.; Mueller-Lantzsch, N.; Sester, U.; Kaul, H.; Pees, H. No Correlation in Epstein-Barr Virus Reactivation between Serological Parameters and Viral Load. J. Clin. Microbiol. 2000, 38, 2458. [Google Scholar] [CrossRef] [PubMed]
- Agostini, S.; Mancuso, R.; Guerini, F.R.; D’Alfonso, S.; Agliardi, C.; Hernis, A.; Zanzottera, M.; Barizzone, N.; Leone, M.A.; Caputo, D.; et al. HLA Alleles Modulate EBV Viral Load in Multiple Sclerosis. J. Transl. Med. 2018, 16, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartung, H.-P.; Derfuss, T.; Cree, B.A.; Sormani, M.P.; Selmaj, K.; Stutters, J.; Prados, F.; MacManus, D.; Schneble, H.-M.; Lambert, E.; et al. Efficacy and Safety of Temelimab in Multiple Sclerosis: Results of a Randomized Phase 2b and Extension Study. Mult. Scler. Houndmills Basingstoke Engl. 2021, 13524585211024996. [Google Scholar] [CrossRef]
- García-Montojo, M.; de la Hera, B.; Varadé, J.; de la Encarnación, A.; Camacho, I.; Domínguez-Mozo, M.; Árias-Leal, A.; García-Martínez, A.; Casanova, I.; Izquierdo, G.; et al. HERV-W Polymorphism in Chromosome X Is Associated with Multiple Sclerosis Risk and with Differential Expression of MSRV. Retrovirology 2014, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Sotgiu, S.; Serra, C.; Mameli, G.; Pugliatti, M.; Rosati, G.; Arru, G.; Dolei, A. Multiple Sclerosis–Associated Retrovirus and MS Prognosis: An Observational Study. Neurology 2002, 59, 1071–1073. [Google Scholar] [CrossRef]
- Sotgiu, S.; Arru, G.; Mameli, G.; Serra, C.; Pugliatti, M.; Rosati, G.; Dolei, A. Multiple Sclerosis-Associated Retrovirus in Early Multiple Sclerosis: A Six-Year Follow-up of a Sardinian Cohort. Mult. Scler. Houndmills Basingstoke Engl. 2006, 12, 698–703. [Google Scholar] [CrossRef]
- Mameli, G.; Cossu, D.; Cocco, E.; Frau, J.; Marrosu, M.G.; Niegowska, M.; Sechi, L.A. Epitopes of HERV-Wenv Induce Antigen-Specific Humoral Immunity in Multiple Sclerosis Patients. J. Neuroimmunol. 2015, 280, 66–68. [Google Scholar] [CrossRef]
- Arru, G.; Caggiu, E.; Leoni, S.; Mameli, G.; Pugliatti, M.; Sechi, G.P.; Sechi, L.A. Natalizumab Modulates the Humoral Response against HERV-Wenv73-88 in a Follow-up Study of Multiple Sclerosis Patients. J. Neurol. Sci. 2015, 357, 106–108. [Google Scholar] [CrossRef]
- Schmitt, K.; Richter, C.; Backes, C.; Meese, E.; Ruprecht, K.; Mayer, J. Comprehensive Analysis of Human Endogenous Retrovirus Group HERV-W Locus Transcription in Multiple Sclerosis Brain Lesions by High-Throughput Amplicon Sequencing. J. Virol. 2013, 87, 13837–13852. [Google Scholar] [CrossRef] [Green Version]
- Bergallo, M.; Galliano, I.; Montanari, P.; Zaniol, E.; Graziano, E.; Calvi, C.; Alliaudi, C.; Daprà, V.; Savino, F. Modulation of Human Endogenous Retroviruses -H, -W and -K Transcription by Microbes. Microbes Infect. 2020, 22, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Mozo, M.I.; Nieto-Guerrero, A.; Pérez-Pérez, S.; García-Martínez, M.Á.; Arroyo, R.; Álvarez-Lafuente, R. MicroRNAs of Human Herpesvirus 6A and 6B in Serum and Cerebrospinal Fluid of Multiple Sclerosis Patients. Front. Immunol. 2020, 11, 2142. [Google Scholar] [CrossRef] [PubMed]
- Kula, T.; Dezfulian, M.H.; Wang, C.I.; Abdelfattah, N.S.; Hartman, Z.C.; Wucherpfennig, K.W.; Lyerly, H.K.; Elledge, S.J. T-Scan: A Genome-Wide Method for the Systematic Discovery of T Cell Epitopes. Cell 2019, 178, 1016–1028.e13. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| RRMS | |
|---|---|
| n | 98 |
| Sex (% females) | 70.4 |
| Age (y.o., mean ± SD) | 40.2 ± 7.7 |
| Disease duration (m, median (P25–P75)) | 118.0 (63.3–178.0) |
| Treatment (%): | |
| First line (beta interferon, glatiramer acetate) | 43.9 |
| Second line (natalizumab) | 56.1 |
| Treatment duration (m, median (P25–P75)): | |
| First line | 45.0 (24.0–62.0) |
| Second line | 12.0 (2.0–27.0) |
| EDSS (median (P25–P75)) | 2.0 (1.0–3.0) |
| MSSS (median (P25–P75)) | 2.3 (0.8–4.2) |
| Annualized relapse rate (median (P25–P75)) | 0.6 (0.4–0.9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Pérez, S.; Domínguez-Mozo, M.I.; García-Martínez, M.Á.; Ballester-González, R.; Nieto-Gañán, I.; Arroyo, R.; Alvarez-Lafuente, R. Epstein-Barr Virus Load Correlates with Multiple Sclerosis-Associated Retrovirus Envelope Expression. Biomedicines 2022, 10, 387. https://doi.org/10.3390/biomedicines10020387
Pérez-Pérez S, Domínguez-Mozo MI, García-Martínez MÁ, Ballester-González R, Nieto-Gañán I, Arroyo R, Alvarez-Lafuente R. Epstein-Barr Virus Load Correlates with Multiple Sclerosis-Associated Retrovirus Envelope Expression. Biomedicines. 2022; 10(2):387. https://doi.org/10.3390/biomedicines10020387
Chicago/Turabian StylePérez-Pérez, Silvia, María Inmaculada Domínguez-Mozo, María Ángel García-Martínez, Rubén Ballester-González, Israel Nieto-Gañán, Rafael Arroyo, and Roberto Alvarez-Lafuente. 2022. "Epstein-Barr Virus Load Correlates with Multiple Sclerosis-Associated Retrovirus Envelope Expression" Biomedicines 10, no. 2: 387. https://doi.org/10.3390/biomedicines10020387
APA StylePérez-Pérez, S., Domínguez-Mozo, M. I., García-Martínez, M. Á., Ballester-González, R., Nieto-Gañán, I., Arroyo, R., & Alvarez-Lafuente, R. (2022). Epstein-Barr Virus Load Correlates with Multiple Sclerosis-Associated Retrovirus Envelope Expression. Biomedicines, 10(2), 387. https://doi.org/10.3390/biomedicines10020387