
Myotendinous Junction: Exercise Protocols Can Positively Influence Their Development in Rats

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
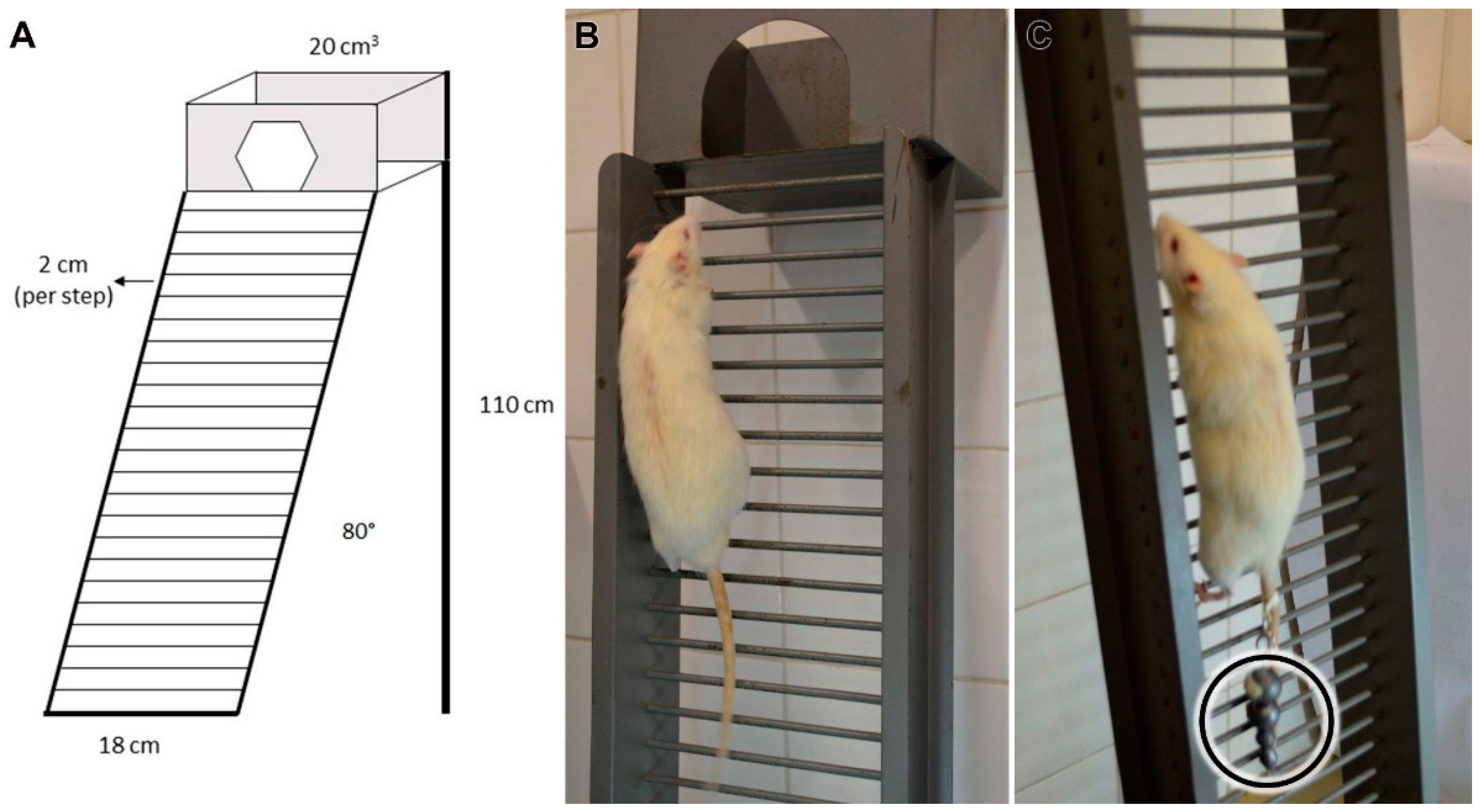
2.1.1. Climbing Protocol
2.1.2. Overload Climbing Protocol
2.1.3. Climbing and Overload Climbing Protocol
2.2. Light Microscopy—Image Area and Fractal Dimension
2.3. Transmission Electron Microscopy (TEM)
2.4. Immunofluorescence
3. Results
3.1. Light Microscopy—Image Area and Fractal Dimension
3.2. Transmission Electron Microscopy
3.2.1. Sarcomeres
3.2.2. Myotendinous Junction
3.2.3. Extracellular Matrix
4. Immunofluorescence
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciena, A.; Luques, I.U.; Dias, F.J.; de Almeida, S.R.Y.; Iyomasa, M.M.; Watanabe, I.-S. Ultrastructure of the myotendinous junction of the medial pterygoid muscle of adult and aged Wistar rats. Micron 2010, 41, 1011–1014. [Google Scholar] [CrossRef] [PubMed]
- Ciena, A.P.; De Almeida, S.R.Y.; Bolina, C.D.S.; Bolina-Matos, R.D.S.; Rici, R.E.G.; Da Silva, M.C.P.; Miglino, M.A.; Watanabe, I.-S. Ultrastructural features of the myotendinous junction of the sternomastoid muscle in Wistar rats: From newborn to aging. Microsc. Res. Tech. 2012, 75, 1292–1296. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G.; Quan, D.M. Modifications in myotendinous junction structure following denervation. Acta Neuropathol. 1992, 84, 135–140. [Google Scholar] [CrossRef]
- Kojima, H.; Sakuma, E.; Mabuchi, Y.; Mizutani, J.; Horiuchi, O.; Wada, I.; Horiba, M.; Yamashita, Y.; Herbert, D.C.; Soji, T.; et al. Ultrastructural changes at the myotendinous junction induced by exercise. J. Orthop. Sci. 2008, 13, 233–239. [Google Scholar] [CrossRef]
- Charvet, B.; Ruggiero, F.; Le Guellec, D. The development of the myotendinous junction: A review. Muscle Ligaments Tendons J. 2012, 2, 53–63. [Google Scholar]
- Tidball, J.G.; Chan, M. Adhesive strength of single muscle cells to basement membrane at myotendinous junctions. J. Appl. Physiol. 1989, 67, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Guzman, M.; Montero, J.A.; Santesteban, E.; Gañan, Y.; Macias, D.; Hurle, J.M. Tendon-muscle crosstalk controls muscle bellies morphogenesis, which is mediated by cell death and retinoic acid signaling. Dev. Biol. 2007, 302, 267–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenbaker, D.G.; De Smet, A.A. MR imaging of muscle injuries. Appl. Radiol. 2004, 33, 14–20. [Google Scholar]
- Fiorentino, N.; Epstein, F.H.; Blemker, S.S. Activation and aponeurosis morphology affect in vivo muscle tissue strains near the myotendinous junction. J. Biomech. 2012, 45, 647–652. [Google Scholar] [CrossRef] [Green Version]
- Guerini, H.; Pluot, E.; Pessis, E.; Thevenin, F.; Campagna, R.; Feydy, A.; Gaudin, P.; Drapé, J. Tears at the myotendinous junction of the infraspinatus: Ultrasound findings. Diagn. Interv. Imaging 2015, 96, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Rocha, L.C.; Pimentel-Neto, J.P.; De Sant’Ana, J.S.; Jacob, C.D.S.; Barbosa, G.K.; Krause-Neto, W.; Watanabe, I.; Ciena, A.P. Repercussions on sarcomeres of the myotendinous junction and the myofibrillar type adaptations in response to different trainings on vertical ladder. Microsc. Res. Tech. 2020, 83, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- Sierra, L.R.; Fávaro, G.; Cerri, B.R.; Rocha, L.C.; Almeida, S.R.D.Y.D.; Watanabe, I.-S.; Ciena, A.P. Myotendinous junction plasticity in aged ovariectomized rats submitted to aquatic training. Microsc. Res. Tech. 2018, 81, 816–822. [Google Scholar] [CrossRef]
- Pimentel-Neto, J.P.; Rocha, L.C.; Barbosa, G.K.; Jacob, C.D.S.; Neto, W.K.; Watanabe, I.-S.; Ciena, A.P. Myotendinous junction adaptations to ladder-based resistance training: Identification of a new telocyte niche. Sci. Rep. 2020, 10, 14124. [Google Scholar] [CrossRef] [PubMed]
- Grillo, B.A.C.; Rocha, L.C.; Martinez, G.Z.; Neto, J.P.; Jacob, C.D.S.; Watanabe, I.-S.; Ciena, A.P. Myotendinous Junction Components of Different Skeletal Muscles Present Morphological Changes in Obese Rats. Microsc. Microanal. 2021, 27, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual Hematoxylin and Eosin Staining of Mouse Tissue Sections. Cold Spring Harb. Protoc. 2014, 2014, 655–658. [Google Scholar] [CrossRef]
- Bhutda, S.; Surve, M.; Anil, A.; Kamath, K.; Singh, N.; Modi, D.; Banerjee, A. Histochemical Staining of Collagen and Identification of Its Subtypes by Picrosirius Red Dye in Mouse Reproductive Tissues. Bio Protoc. 2017, 7, e2598. [Google Scholar] [CrossRef] [PubMed]
- Cury, S.S.; Freire, P.P.; Martinucci, B.; dos Santos, V.C.; de Oliveira, G.; Ferretti, R.; Dal-Pai-Silva, M.; Pacagnelli, F.L.; Delella, F.K.; Carvalho, R.F. Fractal dimension analysis reveals skeletal muscle disorganization in mdx mice. Biochem. Biophys. Res. Commun. 2018, 503, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, G.K.; Jacob, C.D.S.; Pimentel-Neto, J.P.; De Oliveira, M.F.; Rici, R.E.G.; Watanabe, I.; Ciena, A.P. Structural and ultrastructural characteristics of the tongue of the Collared Peccary (Pecari tajacu, Linnaeus, 1758). Anat. Histol. Embryol. 2020, 49, 532–540. [Google Scholar] [CrossRef]
- Jacob, C.D.S.; Rocha, L.C.; Pimentel-Neto, J.P.; Watanabe, I.-S.; Ciena, A.P. Effects of physical training on sarcomere lengths and muscle-tendon interface of the cervical region in an experimental model of menopause. Eur. J. Histochem. 2019, 63, 131–135. [Google Scholar] [CrossRef]
- Rocha, L.C.; Jacob, C.D.S.; Barbosa, G.K.; Pimentel-Neto, J.P.; Krause Neto, W.; Gama, E.F.; Ciena, A.P. Remodeling of the skeletal muscle and postsynaptic component after short-term joint immobilization and aquatic training. Histochem. Cell Biol. 2020, 154, 621–628. [Google Scholar] [CrossRef]
- Crameri, R.M.; Langberg, H.; Magnusson, P.; Jensen, C.H.; Schrøder, H.D.; Olesen, J.L.; Suetta, C.; Teisner, B.; Kjaer, M. Changes in satellite cells in human skeletal muscle after a single bout of high intensity exercise. J. Physiol. 2004, 558, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Wackerhage, H.; Schoenfeld, B.J.; Hamilton, D.; Lehti, M.; Hulmi, J.J. Stimuli and sensors that initiate skeletal muscle hypertrophy following resistance exercise. J. Appl. Physiol. 2019, 126, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Korolj, A.; Wu, H.-T.; Radisic, M. A healthy dose of chaos: Using fractal frameworks for engineering higher-fidelity biomedical systems. Biomaterials 2019, 219, 119363. [Google Scholar] [CrossRef] [PubMed]
- Curzi, D.; Salucci, S.; Marini, M.; Esposito, F.; Agnello, L.; Veicsteinas, A.; Burattini, S.; Falcieri, E. How physical exercise changes rat myotendinous junctions: An ultrastructural study. Eur. J. Histochem. 2012, 56, e19. [Google Scholar] [CrossRef] [Green Version]
- Guzzoni, V.; Selistre-De-Araújo, H.S.; Marqueti, R.D.C. Tendon Remodeling in Response to Resistance Training, Anabolic Androgenic Steroids and Aging. Cells 2018, 7, 251. [Google Scholar] [CrossRef] [Green Version]
- Hart, D.A.; Zernicke, R.F. Optimal Human Functioning Requires Exercise Across the Lifespan: Mobility in a 1g Environment Is Intrinsic to the Integrity of Multiple Biological Systems. Front. Physiol. 2020, 11, 156. [Google Scholar] [CrossRef] [Green Version]
- Curzi, D.; Baldassarri, V.; De Matteis, R.; Salamanna, F.; Bolotta, A.; Frizziero, A. Morphological adaptation and protein modulation of myotendinous junction following moderate aerobic training. Histol. Histopathol. 2015, 30, 465–472. [Google Scholar]
- Hohenester, E.; Yurchenco, P.D. Laminins in basement membrane assembly. Cell Adhes. Migr. 2013, 7, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Popescu, L.M.; Faussone-Pellegrini, M.-S. Telocytes—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to Telocytes. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Rayagiri, S.S.; Ranaldi, D.; Raven, A.; Azhar, N.I.F.M.; Lefebvre, O.; Zammit, P.; Borycki, A.-G. Basal lamina remodeling at the skeletal muscle stem cell niche mediates stem cell self-renewal. Nat. Commun. 2018, 9, 1075. [Google Scholar] [CrossRef] [Green Version]
- Dahiya, S.; Bhatnagar, S.; Hindi, S.M.; Jiang, C.; Paul, P.K.; Kuang, S.; Kumar, A. Elevated levels of active matrix metalloproteinase-9 cause hypertrophy in skeletal muscle of normal and dystrophin-deficient mdx mice. Hum. Mol. Genet. 2011, 20, 4345–4359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, N.A.; Wang, Y.X.; Rudnicki, M.A. Intrinsic and extrinsic mechanisms regulating satellite cell function. Development 2015, 142, 1572–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanes, J.R. The Basement Membrane/Basal Lamina of Skeletal Muscle. J. Biol. Chem. 2003, 278, 12601–12604. [Google Scholar] [CrossRef] [Green Version]
- Gumucio, J.P.; Sugg, K.B.; Mendias, C.L. TGF-β Superfamily Signaling in Muscle and Tendon Adaptation to Resistance Exercise. Exerc. Sport Sci. Rev. 2015, 43, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.E.; Gumucio, J.P.; Sugg, K.B.; Bedi, A.; Mendias, C.L. MMP inhibition as a potential method to augment the healing of skeletal muscle and tendon extracellular matrix. J. Appl. Physiol. 2013, 115, 884–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjaer, M. Role of Extracellular Matrix in Adaptation of Tendon and Skeletal Muscle to Mechanical Loading. Physiol. Rev. 2004, 84, 649–698. [Google Scholar] [CrossRef] [PubMed]
- Mendias, C.L.; Schwartz, A.J.; Grekin, J.A.; Gumucio, J.P.; Sugg, K.B. Changes in muscle fiber contractility and extracellular matrix production during skeletal muscle hypertrophy. J. Appl. Physiol. 2017, 122, 571–579. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, S.; An, T.; Tarique, I.; Vistro, W.A.; Liu, Y.; Wang, Z.; Zhang, H.; Shi, Y.; Haseeb, A.; et al. Telocytes as a Novel Structural Component in the Muscle Layers of the Goat Rumen. Cell Transplant. 2019, 28, 955–966. [Google Scholar] [CrossRef]
- Marini, M.; Rosa, I.; Ibba-Manneschi, L.; Manetti, M. Telocytes in skeletal, cardiac and smooth muscle interstitium: Morphological and functional aspects. Histol. Histopathol. 2018, 33, 1151–1165. [Google Scholar]
- Rosa, I.; Taverna, C.; Novelli, L.; Marini, M.; Ibba-Manneschi, L.; Manetti, M. Telocytes constitute a widespread interstitial meshwork in the lamina propria and underlying striated muscle of human tongue. Sci. Rep. 2019, 9, 5858. [Google Scholar] [CrossRef] [Green Version]
- Bosco, C.; Díaz, E.; Gutiérrez, R.; González, J.; Pérez, J. Ganglionar nervous cells and telocytes in the pancreas of Octodon degus: Extra and intrapancreatic ganglionar cells and telocytes in the degus. Auton. Neurosci. 2013, 177, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liang, Y.; Wang, S.; Tarique, I.; Vistro, W.A.; Zhang, H.; Haseeb, A.; Gandahi, N.S.; Iqbal, A.; An, T.; et al. Identification and characterization of telocytes in rat testis. Aging 2019, 11, 5757–5768. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, Y.; Song, Y.; Huang, Q.; Wang, F.; Yang, W.; Yang, C. Telocytes in Liver. Curr. Stem Cell Res. Ther. 2016, 11, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.; Barbosa, G.; Pimentel-Neto, J.P.; Jacob, C.; Knudsen, A.; Watanabe, I.-S.; Ciena, A. Aquatic Training after Joint Immobilization in Rats Promotes Adaptations in Myotendinous Junctions. Int. J. Mol. Sci. 2021, 22, 6983. [Google Scholar] [CrossRef]
- Ravalli, S.; Federico, C.; Lauretta, G.; Saccone, S.; Pricoco, E.; Roggio, F.; Di Rosa, M.; Maugeri, G.; Musumeci, G. Morphological Evidence of Telocytes in Skeletal Muscle Interstitium of Exercised and Sedentary Rodents. Biomedicines 2021, 9, 807. [Google Scholar] [CrossRef]
- Urciuolo, A.; Quarta, M.; Morbidoni, V.; Gattazzo, F.; Molon, S.; Grumati, P.; Montemurro, F.; Tedesco, F.S.; Blaauw, B.; Cossu, G.; et al. Collagen VI regulates satellite cell self-renewal and muscle regeneration. Nat. Commun. 2013, 4, 1964. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | |||||
|---|---|---|---|---|---|
| C | OC | COC | |||
| Light Microscopy | Muscle | Nuclei FD (HE) | ↑↑ | ↑↑↑ | ↑ |
| Nuclei Area (HE) | ↑↑ | ↑ | ↓ | ||
| Collagen FD (PSR) | ↑↑ | ↑ | ↑↑↑ | ||
| Collagen Area (PSR) | ↑↑ | ↑↑↑ | ↑ | ||
| Muscle Fiber FD (PSR) | ↓ | ↓↓ | ↓↓↓ | ||
| Muscle Fiber Area (PSR) | ↓ | ↑ | ↑↑ | ||
| Transmission Electron Microscopy | Sarcomeres | Belly | ↑↑ | ↑ | ↓ |
| Proximal | ↑ | ↓ | ↓↓ | ||
| Distal | ↑ | ↓ | ↑↑ | ||
| Length | Sarcoplasmic Invaginations | ↓ | ↑↑ | ↑ | |
| Sarcoplasmic Evaginations | ↑ | ↑↑↑ | ↑↑ | ||
| Thickness | Sarcoplasmic Invaginations | ↑↑ | ↓ | ↑ | |
| Sarcoplasmic Evaginations | ↓↓ | ↓↓↓ | ↓ | ||
| Myotendinous Interface | ↑ | ↑↑↑ | ↑↑ | ||
| Extracellular Matrix | Basal Lamina | ↑ | ↑↑ | ↓ | |
| Support Collagen Layer | ↑↑ | ↑ | ↑↑↑ | ||
| Collagen Fiber Perimeter | ↑ | ↓ | ↑↑ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimentel Neto, J.; Rocha-Braga, L.C.; Jacob, C.d.S.; Tomiate, A.N.; Ciena, A.P. Myotendinous Junction: Exercise Protocols Can Positively Influence Their Development in Rats. Biomedicines 2022, 10, 480. https://doi.org/10.3390/biomedicines10020480
Pimentel Neto J, Rocha-Braga LC, Jacob CdS, Tomiate AN, Ciena AP. Myotendinous Junction: Exercise Protocols Can Positively Influence Their Development in Rats. Biomedicines. 2022; 10(2):480. https://doi.org/10.3390/biomedicines10020480
Chicago/Turabian StylePimentel Neto, Jurandyr, Lara Caetano Rocha-Braga, Carolina dos Santos Jacob, André Neri Tomiate, and Adriano Polican Ciena. 2022. "Myotendinous Junction: Exercise Protocols Can Positively Influence Their Development in Rats" Biomedicines 10, no. 2: 480. https://doi.org/10.3390/biomedicines10020480
APA StylePimentel Neto, J., Rocha-Braga, L. C., Jacob, C. d. S., Tomiate, A. N., & Ciena, A. P. (2022). Myotendinous Junction: Exercise Protocols Can Positively Influence Their Development in Rats. Biomedicines, 10(2), 480. https://doi.org/10.3390/biomedicines10020480