
Regenerative Effects of Exosomes-Derived MSCs: An Overview on Spinal Cord Injury Experimental Studies

Abstract
:1. Introduction
2. Methodology
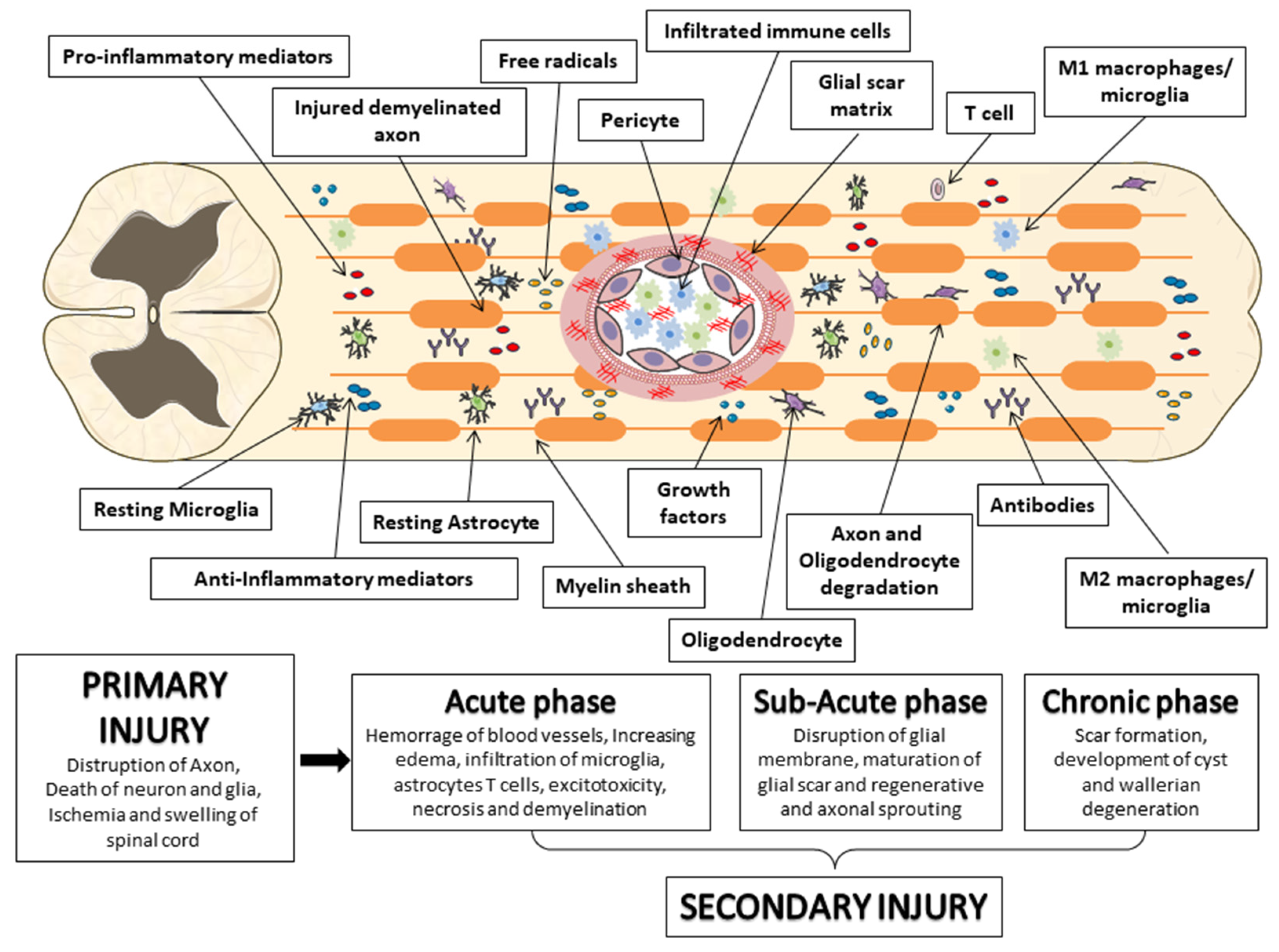
3. Spinal Cord Injury (SCI)
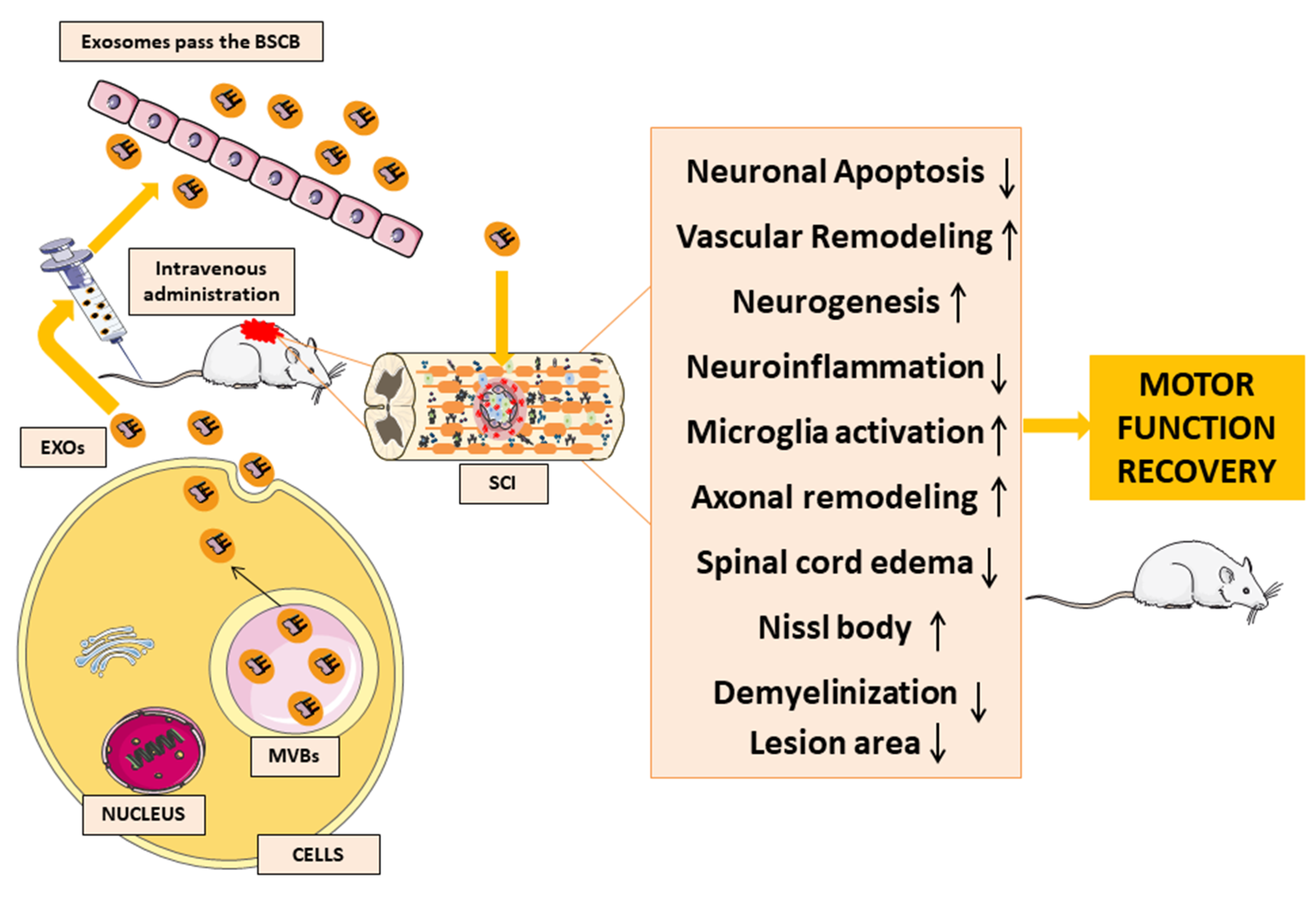
4. MSC-EXOs
5. MSC-EXOs as a Potential Therapeutic Tool in SCI
6. MSC-EXOs as a Potential Therapeutic Tool in SCI Experimental Studies
7. Challenges and Future Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, G.U.; Sung, S.E.; Kang, K.K.; Choi, J.H.; Lee, S.; Sung, M.; Yang, S.Y.; Kim, S.K.; Kim, Y.I.; Lim, J.H.; et al. Therapeutic Potential of Mesenchymal Stem Cells (MSCs) and MSC-Derived Extracellular Vesicles for the Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 3672. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Thery, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Aimaletdinov, A.M.; Gomzikova, M.O. Tracking of Extracellular Vesicles’ Biodistribution: New Methods and Approaches. Int. J. Mol. Sci. 2022, 23, 1312. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Izadpanah, M.; Seddigh, A.; Ebrahimi Barough, S.; Fazeli, S.A.S.; Ai, J. Potential of Extracellular Vesicles in Neurodegenerative Diseases: Diagnostic and Therapeutic Indications. J. Mol. Neurosci. 2018, 66, 172–179. [Google Scholar] [CrossRef]
- Yang, Z.; Li, Y.; Wang, Z. Recent Advances in the Application of Mesenchymal Stem Cell-Derived Exosomes for Cardiovascular and Neurodegenerative Disease Therapies. Pharmaceutics 2022, 14, 618. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, G.; Zhang, K.; Cao, Q.; Liu, T.; Li, J. Mesenchymal stem cells-derived exosomes for drug delivery. Stem Cell Res. Ther. 2021, 12, 561. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Aziz, I.; Che Ramli, M.D.; Mohd Zain, N.S.; Sanusi, J. Behavioral and Histopathological Study of Changes in Spinal Cord Injured Rats Supplemented with Spirulina platensis. Evid.-Based Complement. Altern. Med. 2014, 2014, 871657. [Google Scholar] [CrossRef]
- Sykova, E.; Cizkova, D.; Kubinova, S. Mesenchymal Stem Cells in Treatment of Spinal Cord Injury and Amyotrophic Lateral Sclerosis. Front. Cell Dev. Biol. 2021, 9, 695900. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Tetreault, L.; Kalsi-Ryan, S.; Nouri, A.; Fehlings, M.G. Global prevalence and incidence of traumatic spinal cord injury. Clin. Epidemiol. 2014, 6, 309–331. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Lim, J.; Mekary, R.A.; Rattani, A.; Dewan, M.C.; Sharif, S.Y.; Osorio-Fonseca, E.; Park, K.B. Traumatic Spinal Injury: Global Epidemiology and Worldwide Volume. World Neurosurg. 2018, 113, e345–e363. [Google Scholar] [CrossRef]
- Chen, W.C.; Liu, W.F.; Bai, Y.Y.; Zhou, Y.Y.; Zhang, Y.; Wang, C.M.; Lin, S.; He, H.F. Transplantation of mesenchymal stem cells for spinal cord injury: A systematic review and network meta-analysis. J. Transl. Med. 2021, 19, 178. [Google Scholar] [CrossRef]
- Liau, L.L.; Looi, Q.H.; Chia, W.C.; Subramaniam, T.; Ng, M.H.; Law, J.X. Treatment of spinal cord injury with mesenchymal stem cells. Cell Biosci. 2020, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Morales, I.I.; Toscano-Tejeida, D.; Ibarra, A. Non Pharmacological Strategies to Promote Spinal Cord Regeneration: A View on Some Individual or Combined Approaches. Curr. Pharm. Des. 2016, 22, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Tyler, J.Y.; Xu, X.M.; Cheng, J.X. Nanomedicine for treating spinal cord injury. Nanoscale 2013, 5, 8821–8836. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, E.S.; McDonald, J.W. Rodent models for treatment of spinal cord injury: Research trends and progress toward useful repair. Curr. Opin. Neurol. 2004, 17, 121–131. [Google Scholar] [CrossRef]
- Kumar, H.; Ropper, A.E.; Lee, S.H.; Han, I. Propitious Therapeutic Modulators to Prevent Blood-Spinal Cord Barrier Disruption in Spinal Cord Injury. Mol. Neurobiol. 2017, 54, 3578–3590. [Google Scholar] [CrossRef]
- Ahuja, C.S.; Nori, S.; Tetreault, L.; Wilson, J.; Kwon, B.; Harrop, J.; Choi, D.; Fehlings, M.G. Traumatic Spinal Cord Injury-Repair and Regeneration. Neurosurgery 2017, 80, S9–S22. [Google Scholar] [CrossRef]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From basics to clinical: A comprehensive review on spinal cord injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef]
- Ramer, L.M.; Ramer, M.S.; Steeves, J.D. Setting the stage for functional repair of spinal cord injuries: A cast of thousands. Spinal Cord 2005, 43, 134–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; He, X.; Ren, Y. Function of microglia and macrophages in secondary damage after spinal cord injury. Neural Regen. Res. 2014, 9, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Houghtling, R.A.; MacArthur, L.; Bayer, B.M.; Bregman, B.S. Differences in cytokine gene expression profile between acute and secondary injury in adult rat spinal cord. Exp. Neurol. 2003, 184, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Zhivotovsky, B.; Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium 2011, 50, 211–221. [Google Scholar] [CrossRef]
- Park, E.; Velumian, A.A.; Fehlings, M.G. The role of excitotoxicity in secondary mechanisms of spinal cord injury: A review with an emphasis on the implications for white matter degeneration. J. Neurotrauma 2004, 21, 754–774. [Google Scholar] [CrossRef]
- Dong, H.; Fazzaro, A.; Xiang, C.; Korsmeyer, S.J.; Jacquin, M.F.; McDonald, J.W. Enhanced oligodendrocyte survival after spinal cord injury in Bax-deficient mice and mice with delayed Wallerian degeneration. J. Neurosci. 2003, 23, 8682–8691. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Chan, P.H. Reactive oxygen radicals and pathogenesis of neuronal death after cerebral ischemia. Antioxid. Redox Signal. 2003, 5, 597–607. [Google Scholar] [CrossRef]
- Donnelly, D.J.; Popovich, P.G. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef]
- Tran, A.P.; Warren, P.M.; Silver, J. The Biology of Regeneration Failure and Success After Spinal Cord Injury. Physiol. Rev. 2018, 98, 881–917. [Google Scholar] [CrossRef]
- Kwiecien, J.M.; Dabrowski, W.; Dabrowska-Bouta, B.; Sulkowski, G.; Oakden, W.; Kwiecien-Delaney, C.J.; Yaron, J.R.; Zhang, L.; Schutz, L.; Marzec-Kotarska, B.; et al. Prolonged inflammation leads to ongoing damage after spinal cord injury. PLoS ONE 2020, 15, e0226584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.M.; He, C. The glial scar in spinal cord injury and repair. Neurosci. Bull. 2013, 29, 421–435. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.R.; Forgione, N.; Fehlings, M.G. Emerging therapies for acute traumatic spinal cord injury. CMAJ 2013, 185, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Yin, X.; Liu, S.; Hui, H.; Yan, L.; He, B.; Hao, D. Letter: Ultra-Early (<12 Hours) Surgery Correlates With Higher Rate of American Spinal Injury Association Impairment Scale Conversion After Cervical Spinal Cord Injury. Neurosurgery 2019, 85, E399–E400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, B.K.; Curt, A.; Belanger, L.M.; Bernardo, A.; Chan, D.; Markez, J.A.; Gorelik, S.; Slobogean, G.P.; Umedaly, H.; Giffin, M.; et al. Intrathecal pressure monitoring and cerebrospinal fluid drainage in acute spinal cord injury: A prospective randomized trial. J. Neurosurg. 2009, 10, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Bracken, M.B. Steroids for acute spinal cord injury. Cochrane Database Syst. Rev. 2012, 1, CD001046. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Bathen, M.E.; Smith, J.S.; Levi, A.D.; Bhatia, N.N.; Steward, O. Advances in the management of spinal cord injury. J. Am. Acad. Orthop. Surg. 2010, 18, 210–222. [Google Scholar] [CrossRef]
- Hawryluk, G.; Whetstone, W.; Saigal, R.; Ferguson, A.; Talbott, J.; Bresnahan, J.; Dhall, S.; Pan, J.; Beattie, M.; Manley, G. Mean Arterial Blood Pressure Correlates with Neurological Recovery after Human Spinal Cord Injury: Analysis of High Frequency Physiologic Data. J. Neurotrauma 2015, 32, 1958–1967. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Zhang, S.; Wang, M.; Li, Q.; Ren, J.; Luo, Y.; Sun, X. Exosomes in pathogenesis, diagnosis, and therapy of ischemic stroke. Front. Bioeng. Biotechnol. 2022, 10, 980548. [Google Scholar] [CrossRef]
- Kucuk, N.; Primozic, M.; Knez, Z.; Leitgeb, M. Exosomes Engineering and Their Roles as Therapy Delivery Tools, Therapeutic Targets, and Biomarkers. Int. J. Mol. Sci. 2021, 22, 9543. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, S.; Du, C.; Zhang, Y.; Li, Y.; Chu, L.; Han, X.; Galons, H.; Zhang, Y.; Sun, H.; et al. Exosomes from different cells: Characteristics, modifications, and therapeutic applications. Eur. J. Med. Chem. 2020, 207, 112784. [Google Scholar] [CrossRef]
- Yang, X.X.; Sun, C.; Wang, L.; Guo, X.L. New insight into isolation, identification techniques and medical applications of exosomes. J. Control. Release 2019, 308, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhang, W.; Zhang, H.; Zhang, F.; Chen, L.; Ma, L.; Larcher, L.M.; Chen, S.; Liu, N.; Zhao, Q.; et al. Progress, opportunity, and perspective on exosome isolation—Efforts for efficient exosome-based theranostics. Theranostics 2020, 10, 3684–3707. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Antimisiaris, S.G.; Mourtas, S.; Marazioti, A. Exosomes and Exosome-Inspired Vesicles for Targeted Drug Delivery. Pharmaceutics 2018, 10, 218. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.H.; You, S.; Taghizadeh, A.; Taghizadeh, M.; Kim, H.S. Cell Membrane-Cloaked Nanotherapeutics for Targeted Drug Delivery. Int. J. Mol. Sci. 2022, 23, 2223. [Google Scholar] [CrossRef]
- Ding, Y.; Luo, Q.; Que, H.; Wang, N.; Gong, P.; Gu, J. Mesenchymal Stem Cell-Derived Exosomes: A Promising Therapeutic Agent for the Treatment of Liver Diseases. Int. J. Mol. Sci. 2022, 23, 972. [Google Scholar] [CrossRef]
- Juan, T.; Furthauer, M. Biogenesis and function of ESCRT-dependent extracellular vesicles. Semin. Cell Dev. Biol. 2018, 74, 66–77. [Google Scholar] [CrossRef]
- Keller, S.; Sanderson, M.P.; Stoeck, A.; Altevogt, P. Exosomes: From biogenesis and secretion to biological function. Immunol. Lett. 2006, 107, 102–108. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; Porto-Carreiro, I.; Simoes, S.; Raposo, G. Exosomes: A common pathway for a specialized function. J. Biochem. 2006, 140, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Muthu, S.; Bapat, A.; Jain, R.; Jeyaraman, N.; Jeyaraman, M. Exosomal therapy-a new frontier in regenerative medicine. Stem Cell 2021, 8, 7. [Google Scholar] [CrossRef]
- Fitzner, D.; Schnaars, M.; van Rossum, D.; Krishnamoorthy, G.; Dibaj, P.; Bakhti, M.; Regen, T.; Hanisch, U.K.; Simons, M. Selective transfer of exosomes from oligodendrocytes to microglia by macropinocytosis. J. Cell Sci. 2011, 124, 447–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, D.; Zhao, W.L.; Ye, Y.Y.; Bai, X.C.; Liu, R.Q.; Chang, L.F.; Zhou, Q.; Sui, S.F. Cellular internalization of exosomes occurs through phagocytosis. Traffic 2010, 11, 675–687. [Google Scholar] [CrossRef]
- Riazifar, M.; Mohammadi, M.R.; Pone, E.J.; Yeri, A.; Lasser, C.; Segaliny, A.I.; McIntyre, L.L.; Shelke, G.V.; Hutchins, E.; Hamamoto, A.; et al. Stem Cell-Derived Exosomes as Nanotherapeutics for Autoimmune and Neurodegenerative Disorders. ACS Nano 2019, 13, 6670–6688. [Google Scholar] [CrossRef]
- Cui, G.H.; Wu, J.; Mou, F.F.; Xie, W.H.; Wang, F.B.; Wang, Q.L.; Fang, J.; Xu, Y.W.; Dong, Y.R.; Liu, J.R.; et al. Exosomes derived from hypoxia-preconditioned mesenchymal stromal cells ameliorate cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice. FASEB J. 2018, 32, 654–668. [Google Scholar] [CrossRef] [Green Version]
- Pashoutan Sarvar, D.; Shamsasenjan, K.; Akbarzadehlaleh, P. Mesenchymal Stem Cell-Derived Exosomes: New Opportunity in Cell-Free Therapy. Adv. Pharm. Bull. 2016, 6, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Fayazi, N.; Sheykhhasan, M.; Soleimani Asl, S.; Najafi, R. Stem Cell-Derived Exosomes: A New Strategy of Neurodegenerative Disease Treatment. Mol. Neurobiol. 2021, 58, 3494–3514. [Google Scholar] [CrossRef]
- Mathivanan, S.; Simpson, R.J. ExoCarta: A compendium of exosomal proteins and RNA. Proteomics 2009, 9, 4997–5000. [Google Scholar] [CrossRef]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Li, Q.; Zhao, B.; Wang, Y. Stem Cell-Derived Extracellular Vesicles as a Novel Potential Therapeutic Tool for Tissue Repair. Stem Cells Transl. Med. 2017, 6, 1753–1758. [Google Scholar] [CrossRef]
- Guy, R.; Offen, D. Promising Opportunities for Treating Neurodegenerative Diseases with Mesenchymal Stem Cell-Derived Exosomes. Biomolecules 2020, 10, 1320. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Fu, C.H.; Xu, Y.; Yin, X.M.; Cao, Y.; Lin, F.Y. Extracellular Vesicles Derived from Epidural Fat-Mesenchymal Stem Cells Attenuate NLRP3 Inflammasome Activation and Improve Functional Recovery After Spinal Cord Injury. Neurochem. Res. 2020, 45, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhou, Y.; Zhang, R.; Wen, L.; Wu, K.; Li, Y.; Yao, Y.; Duan, R.; Jia, Y. Bone Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Recovery Following Spinal Cord Injury via Improvement of the Integrity of the Blood-Spinal Cord Barrier. Front. Neurosci. 2019, 13, 209. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Zhao, C.; Hou, S.; Zhou, W.; Wang, B.; Chen, Y. Exosomes secreted from miRNA-29b-modified mesenchymal stem cells repaired spinal cord injury in rats. Braz. J. Med. Biol. Res. 2019, 52, e8735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biglari, B.; Swing, T.; Child, C.; Buchler, A.; Westhauser, F.; Bruckner, T.; Ferbert, T.; Jurgen Gerner, H.; Moghaddam, A. A pilot study on temporal changes in IL-1beta and TNF-alpha serum levels after spinal cord injury: The serum level of TNF-alpha in acute SCI patients as a possible marker for neurological remission. Spinal Cord 2015, 53, 510–514. [Google Scholar] [CrossRef]
- Kwon, B.K.; Streijger, F.; Fallah, N.; Noonan, V.K.; Belanger, L.M.; Ritchie, L.; Paquette, S.J.; Ailon, T.; Boyd, M.C.; Street, J.; et al. Cerebrospinal Fluid Biomarkers To Stratify Injury Severity and Predict Outcome in Human Traumatic Spinal Cord Injury. J. Neurotrauma 2017, 34, 567–580. [Google Scholar] [CrossRef]
- Romanelli, P.; Bieler, L.; Scharler, C.; Pachler, K.; Kreutzer, C.; Zaunmair, P.; Jakubecova, D.; Mrowetz, H.; Benedetti, B.; Rivera, F.J.; et al. Extracellular Vesicles Can Deliver Anti-inflammatory and Anti-scarring Activities of Mesenchymal Stromal Cells After Spinal Cord Injury. Front. Neurol. 2019, 10, 1225. [Google Scholar] [CrossRef]
- Davis, B.K.; Wen, H.; Ting, J.P. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu. Rev. Immunol. 2011, 29, 707–735. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Li, M.; He, F.; Zhou, S.; Zhu, L. Targeting the NLRP3 inflammasome to attenuate spinal cord injury in mice. J. Neuroinflamm. 2017, 14, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Huang, Y.; He, F.; Liu, J.; Li, M.; Sun, T.; Ren, W.; Hou, J.; Zhu, L. Dopamine D1 Receptor Agonist A-68930 Inhibits NLRP3 Inflammasome Activation, Controls Inflammation, and Alleviates Histopathology in a Rat Model of Spinal Cord Injury. Spine 2016, 41, E330–E334. [Google Scholar] [CrossRef]
- Anderson, A.J.; Robert, S.; Huang, W.; Young, W.; Cotman, C.W. Activation of complement pathways after contusion-induced spinal cord injury. J. Neurotrauma 2004, 21, 1831–1846. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.A.; Arora, M.; Bohlson, S.S.; Lozano, E.; Tenner, A.J. Generation of inhibitory NFkappaB complexes and phosphorylated cAMP response element-binding protein correlates with the anti-inflammatory activity of complement protein C1q in human monocytes. J. Biol. Chem. 2007, 282, 7360–7367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brambilla, R.; Bracchi-Ricard, V.; Hu, W.H.; Frydel, B.; Bramwell, A.; Karmally, S.; Green, E.J.; Bethea, J.R. Inhibition of astroglial nuclear factor kappaB reduces inflammation and improves functional recovery after spinal cord injury. J. Exp. Med. 2005, 202, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Pei, S.; Han, L.; Guo, B.; Li, Y.; Duan, R.; Yao, Y.; Xue, B.; Chen, X.; Jia, Y. Mesenchymal Stem Cell-Derived Exosomes Reduce A1 Astrocytes via Downregulation of Phosphorylated NFkappaB P65 Subunit in Spinal Cord Injury. Cell. Physiol. Biochem. 2018, 50, 1535–1559. [Google Scholar] [CrossRef]
- Guerrero, A.R.; Uchida, K.; Nakajima, H.; Watanabe, S.; Nakamura, M.; Johnson, W.E.; Baba, H. Blockade of interleukin-6 signaling inhibits the classic pathway and promotes an alternative pathway of macrophage activation after spinal cord injury in mice. J. Neuroinflamm. 2012, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef]
- David, S.; Kroner, A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Li, G.; Li, D.; Huang, W.; Zhang, R.; Zhang, H.; Duan, Y.; Wang, B. hucMSC derived exosomes promote functional recovery in spinal cord injury mice via attenuating inflammation. Mater. Sci. Eng. C 2018, 89, 194–204. [Google Scholar] [CrossRef]
- Lankford, K.L.; Arroyo, E.J.; Nazimek, K.; Bryniarski, K.; Askenase, P.W.; Kocsis, J.D. Intravenously delivered mesenchymal stem cell-derived exosomes target M2-type macrophages in the injured spinal cord. PLoS ONE 2018, 13, e0190358. [Google Scholar] [CrossRef] [Green Version]
- Reinhold, A.K.; Rittner, H.L. Barrier function in the peripheral and central nervous system-a review. Pflug. Arch. 2017, 469, 123–134. [Google Scholar] [CrossRef]
- Hu, J.; Yu, Q.; Xie, L.; Zhu, H. Targeting the blood-spinal cord barrier: A therapeutic approach to spinal cord protection against ischemia-reperfusion injury. Life Sci. 2016, 158, 1–6. [Google Scholar] [CrossRef]
- Jo, D.H.; Kim, J.H.; Heo, J.I.; Kim, J.H.; Cho, C.H. Interaction between pericytes and endothelial cells leads to formation of tight junction in hyaloid vessels. Mol. Cells 2013, 36, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Lizana, J.; Aliaga, N.; Marani, W.; Escribano, A.; Montemurro, N. Spinal Vascular Shunts: Single-Center Series and Review of the Literature of Their Classification. Neurol. Int. 2022, 14, 581–599. [Google Scholar] [CrossRef]
- Ota, T.; Komiyama, M. Pathogenesis of non-hereditary brain arteriovenous malformation and therapeutic implications. Interv. Neuroradiol. 2020, 26, 244–253. [Google Scholar] [CrossRef]
- Yuan, X.; Wu, Q.; Wang, P.; Jing, Y.; Yao, H.; Tang, Y.; Li, Z.; Zhang, H.; Xiu, R. Exosomes Derived From Pericytes Improve Microcirculation and Protect Blood-Spinal Cord Barrier After Spinal Cord Injury in Mice. Front. Neurosci. 2019, 13, 319. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.Q.; Chen, J.; Wang, S.N.; Duan, F.X.; Chen, Y.Q.; Shi, Y.J.; Hu, J.G.; Lu, H.Z. Identification of serum exosomal microRNAs in acute spinal cord injured rats. Exp. Biol. Med. 2019, 244, 1149–1161. [Google Scholar] [CrossRef]
- Buller, B.; Liu, X.; Wang, X.; Zhang, R.L.; Zhang, L.; Hozeska-Solgot, A.; Chopp, M.; Zhang, Z.G. MicroRNA-21 protects neurons from ischemic death. FEBS J. 2010, 277, 4299–4307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Chen, F.; Ge, X.; Tan, J.; Lei, P.; Zhang, J. miR-21 alleviated apoptosis of cortical neurons through promoting PTEN-Akt signaling pathway in vitro after experimental traumatic brain injury. Brain Res. 2014, 1582, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.M.; Gibbs, K.M.; Davila, J.; Campbell, N.; Sung, S.; Todorova, T.I.; Otsuka, S.; Sabaawy, H.E.; Hart, R.P.; Schachner, M. MicroRNA miR-133b is essential for functional recovery after spinal cord injury in adult zebrafish. Eur. J. Neurosci. 2011, 33, 1587–1597. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Zeng, L.; Huang, J.; Wang, G.; Lu, H. miR-126 promotes angiogenesis and attenuates inflammation after contusion spinal cord injury in rats. Brain Res. 2015, 1608, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Sung, S.E.; Seo, M.S.; Kim, Y.I.; Kang, K.K.; Choi, J.H.; Lee, S.; Sung, M.; Yim, S.G.; Lim, J.H.; Seok, H.G.; et al. Human Epidural AD-MSC Exosomes Improve Function Recovery after Spinal Cord Injury in Rats. Biomedicines 2022, 10, 678. [Google Scholar] [CrossRef]
- Zhou, Y.; Wen, L.L.; Li, Y.F.; Wu, K.M.; Duan, R.R.; Yao, Y.B.; Jing, L.J.; Gong, Z.; Teng, J.F.; Jia, Y.J. Exosomes derived from bone marrow mesenchymal stem cells protect the injured spinal cord by inhibiting pericyte pyroptosis. Neural Regen. Res. 2022, 17, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, C.; Xu, Y.; Li, C.; Cao, Y.; Li, P. Exosomes derived from human placenta-derived mesenchymal stem cells improve neurologic function by promoting angiogenesis after spinal cord injury. Neurosci. Lett. 2020, 739, 135399. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tian, Z.; He, L.; Liu, C.; Wang, N.; Rong, L.; Liu, B. Exosomes derived from miR-26a-modified MSCs promote axonal regeneration via the PTEN/AKT/mTOR pathway following spinal cord injury. Stem Cell Res. Ther. 2021, 12, 224. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Y.; Mu, J.; Chen, J.; Zhang, C.; Cao, H.; Gao, J. Transplantation of Human Mesenchymal Stem-Cell-Derived Exosomes Immobilized in an Adhesive Hydrogel for Effective Treatment of Spinal Cord Injury. Nano Lett. 2020, 20, 4298–4305. [Google Scholar] [CrossRef]
- Wang, Y.; Lai, X.; Wu, D.; Liu, B.; Wang, N.; Rong, L. Umbilical mesenchymal stem cell-derived exosomes facilitate spinal cord functional recovery through the miR-199a-3p/145-5p-mediated NGF/TrkA signaling pathway in rats. Stem Cell Res. Ther. 2021, 12, 117. [Google Scholar] [CrossRef]
- Ji, W.; Jiang, W.; Li, M.; Li, J.; Li, Z. miR-21 deficiency contributes to the impaired protective effects of obese rat mesenchymal stem cell-derived exosomes against spinal cord injury. Biochimie 2019, 167, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Li, Z.; Zhi, Z.; Wang, S.; Xu, G. MiR-21 derived from the exosomes of MSCs regulates the death and differentiation of neurons in patients with spinal cord injury. Gene Ther. 2019, 26, 491–503. [Google Scholar] [CrossRef]
- Meng, F.; Henson, R.; Wehbe-Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Song, Y.; Han, X.; Qu, P.; Wang, W. Long noncoding RNA PTENP1 affects the recovery of spinal cord injury by regulating the expression of miR-19b and miR-21. J. Cell. Physiol. 2020, 235, 3634–3645. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Ao, R.; Zhi, Z.; Jia, J.; Yu, B. miR-21 and miR-19b delivered by hMSC-derived EVs regulate the apoptosis and differentiation of neurons in patients with spinal cord injury. J. Cell. Physiol. 2019, 234, 10205–10217. [Google Scholar] [CrossRef]
- Huang, J.H.; Xu, Y.; Yin, X.M.; Lin, F.Y. Exosomes Derived from miR-126-modified MSCs Promote Angiogenesis and Neurogenesis and Attenuate Apoptosis after Spinal Cord Injury in Rats. Neuroscience 2020, 424, 133–145. [Google Scholar] [CrossRef]
- Li, D.; Zhang, P.; Yao, X.; Li, H.; Shen, H.; Li, X.; Wu, J.; Lu, X. Exosomes Derived From miR-133b-Modified Mesenchymal Stem Cells Promote Recovery After Spinal Cord Injury. Front. Neurosci. 2018, 12, 845. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Zhang, J. Mesenchymal stem cell-derived exosomes containing miR-145-5p reduce inflammation in spinal cord injury by regulating the TLR4/NF-kappaB signaling pathway. Cell Cycle 2021, 20, 993–1009. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Y.; Gong, F.; Rong, Y.; Luo, Y.; Tang, P.; Zhou, Z.; Zhou, Z.; Xu, T.; Jiang, T.; et al. Exosomes Derived from Bone Mesenchymal Stem Cells Repair Traumatic Spinal Cord Injury by Suppressing the Activation of A1 Neurotoxic Reactive Astrocytes. J. Neurotrauma 2019, 36, 469–484. [Google Scholar] [CrossRef]
- Nakazaki, M.; Morita, T.; Lankford, K.L.; Askenase, P.W.; Kocsis, J.D. Small extracellular vesicles released by infused mesenchymal stromal cells target M2 macrophages and promote TGF-beta upregulation, microvascular stabilization and functional recovery in a rodent model of severe spinal cord injury. J. Extracell. Vesicles 2021, 10, e12137. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Rong, Y.; Wang, J.; Zhou, Z.; Ge, X.; Ji, C.; Jiang, D.; Gong, F.; Li, L.; Chen, J.; et al. Exosome-shuttled miR-216a-5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization. J. Neuroinflamm. 2020, 17, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.R.; Kyung, J.W.; Kumar, H.; Kwon, S.P.; Song, S.Y.; Han, I.B.; Kim, B.S. Targeted Delivery of Mesenchymal Stem Cell-Derived Nanovesicles for Spinal Cord Injury Treatment. Int. J. Mol. Sci. 2020, 21, 4185. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cell Therapy | Dose | Rout of Administration | Intervention | Results | Type of Study | Ref. |
|---|---|---|---|---|---|---|
| hEpi ADMSCs-EXOs | 1 × 109 and 5 × 109 particles in 0.2 mL PBS | Intravenous administration | Immediately after SCI was induced, the hEpi ADMSCs-EXOs were injected into the animals and the same amount was administered again after 3 days. | The hEpi ADMSC-EXOs injection improved SCI and reduced the inflammatory response of spinal cord injury through the regulation of various cytokines and targeting immune response and neurogenesis-related genes in the spinal cord tissue. Conversely, Increased the expression of neurotrophin factors such as BDNF and VEGF. | In vivo | [96] |
| BMMSCs-EXOs | 200 μg/mL, ~1 × 106 | Intravenous administration | BMMSCs-EXOs were administered 30 min and 1 day after SCI | The BMSC-EXOs counteracted neuronal cell death, and reduced myelin loss, improving myelin disposition. Furthermore, the treatment increased pericytic/endothelial cell coverage on the vascular wall, inhibited caspase 1 and IL-1β expression, decreased blood-spinal barrier leakage, and promoted accelerated functional recovery in rats. with SCI. In addition, in vivo exposure to BMSC-EXOs it reduced pericyte pyroptosis and increased its survival rate. | In vivo | [97] |
| 100 μg/mL | Pericytes co-incubation and exposure to BMSCs-EXOs | Pericytes were co-incubated with or without BMSCs-EXOs for 8 h before exposure to a compound stimulus of IFN-γ + TNF-α + LPS + Lipofectamine + ATP | In vitro | |||
| hPMSCs-EXOs | 200 μg/μL | Intrathecal injection | Exosomes were directly injected by stereotactic injection into the epicenter of the SCI after damage | The hPMSCs-Exos have proangiogenic effects on endothelial cells inducing tube formation. In addition, in vivo, the hPMSCs-Exos treatment enhanced angiogenesis in the SCI rats and promoted functional recovery. | In vivo | [98] |
| 100 μg/mL | HUVECs SCI model exposed to hPMSCs-EXOs | HUVECs have been cultivated in OGD conditions and undergo scratch in order to induce the SCI model and subsequently exposed to hPMSCs-EXOs | In vitro | |||
| BMMSCs-EXOs-miR-26a | 20 μg/ml | Intravenous administration | BM-MSCs were transfected with the mimics of miR-26a, and the exosomes were collected. Subsequently, PC12 cells were incubated with BMMSCs-EXOs-miR-26a for 48 h. | BMMSCs-Exos-miR-26a induced neurofilament generation in vitro reducing PTEN expression and increasing the PI3K, AKT, and mTOR proteins phosphorylation. In vivo, treatment enhanced axonal regeneration, and neurogenesis, conversely, it reduced glial scarring and improved functional recovery through PTEN/AKT/mTOR signaling cascades. | In vitro | [99] |
| 200 μg in 200 μL PBS | PC12 cells incubated with BMMSCs-EXOs-miR-26a | Immediately following SCI, the rats received an injection of BMMSCs-EXOs-miR-26a via tail vein injection. | In vivo | |||
| BMMSCs-EXOs containing miR-544 | 100 μg diluted in 0.5 mL PBS | Intravenous administration | 24 h after SCI, the rat model was induced, the animals received an injection of BMMSCs-EXOs containing miR-544 via tail vein injection. | BMMSCs-EXOs containing miR-544 reduced inflammation and improved both neuronal survival and consequently promoted functional recovery after SCI | In vivo | [100] |
| hUCMSCs-EXOs co-transfected with miR-199a-3p/145-5p | 0, 1, 3, 5, 7, 10, 13, 15, 17, 20, 25 μg/mL | PC12 cells incubated with hUCMSCs-EXOs co-transfected with miR-199a-3p/145-5p | After being pretreated with hUCMSCs-EXOs or with hUCMSCs-EXOs co-transfected with miR-199a-3p/145-5p, the PC12 cells were treated with LPS (11 μg/mL) in order to establish an injury model. | Administration of hUCMSCs-EXOs co-transfected with miR-199a-3p/145-5p to neurons of SCI rats upregulated TrkA expression at the site of injury. Consequently, the downstream pathways of NGF/TrkA and Akt were inactivated. Thus, the treatment promoted locomotor recovery in SCI rats indicating that hUC-MSC-EXOs may be a promising treatment strategy for SCI. | In vitro | [101] |
| 200 μg | Intravenous administration | SD rats were induced with laminectomy and subsequently underwent artery clamp to trigger acute trauma. After the SCI model was induced, the animals were injected via the tail vein with hUCMSCs-EXOs or hUCMSCs-EXOs co-transfected with miR-199a-3p/145-5p | In vivo | |||
| MSC-EXOs isolated from obese rats transfected with miR-21 | 100 mg in 0.5 mL of PBS | Intravenous administration | The animals were treated with MSC-EXOs isolated from obese rats transfected with miR-21 mimic or without transfection 24 h after injury | The MSC-EXOs isolated from obese rats, due to a reduction in the levels of miR-21, as a result of insulin resistance, do not exert protective effects against skiing. On the contrary, the MSC-EXOs transfected with miR-21 Mimic showed an increase in the level of miR-21 in MSC-EXOs isolated from obese rats decreased cell apoptosis and area of injury, thus recovering their protective effects against this pathology. | In vivo | [97] |
| MSC-EXOs transfected with miR-21 | - | Intravenous administration | Immediately following SCI, the rats received an injection of MSC-EXOs transfected with miR-21 via tail vein injection. | The MSC-EXOs transfected with miR-21 protected neuronal cells from SCI-induced apoptosis and improved the functional recovery after injury by the miR-21/PTEN/PDCD4 signaling pathway | In vivo | [98] |
| - | SH-SY5Y and U251 incubated with MSC-EXOs transfected with miR-21 or PTEN siRNA | SH-SY5Y and U251 cells were firstly transfected with miR-21, or PTEN siRNA using Lipofectamine 2000 and subsequently were treated MSC-EXOs transfected with miR-21 or PTEN siRNA for 48 h | In vitro | |||
| PC12 cells/ /MSC-EXOs co-transfected with miR-21/miR-19b | - | SH-SY5Y and U251 incubated with PC12 cells/MSC-EXOs co-transfected with miR-21/miR-19b | SH-SY5Y and U251 cells were transfected with miR-21, miR-19, or PTEN siRNA using Lipofectamine 2000 and subsequently were treated with PC12 cells/MSC-EXOs transfected with miR-21/miR-19b for 48 h. | PC12 cells/MSC-EXOs transfected with miR-21/miR-19b suppressed the apoptosis of neuron cells by downregulating the PTEN expression. | In vitro | [105] |
| - | Intravenous administration | After SCI, the animals were treated with PC12 cells derived EXOs or with MSC-EXOs transfected with miR-21/miR-19 or PTEN siRNA. | In vivo | |||
| PC12 cells/MSC-EXOs | SH-SY5Y and U251 incubated with PC12 cells/MSC-EXOs | SH-SY5Y and U251 cells were co-transfected with miR-21 and wild type/mutant PTEN mRNA or miR-19b and wild type/mutant PTEN mRNA using Lipofectamine 2000 for 48 h | In EXOs derived from MSCs and PC12 cells, the elevated expression of miR-21 and miR19b reduced PTEN expression and attenuated apoptosis in neuronal cells confirming the therapeutic effects of PC12 cells/MSC-EXOs by downregulating the expression of PTEN. | In vitro | [106] | |
| Intravenous administration | After SCI, the animals were treated with PC12 cells derived EXOs or with MSC-EXOs. | In vivo | ||||
| MSC-EXOs transfected with miR-126 | 100 μg, about 1 × 1010 particles diluted in 0.5 mL of PBS | Intravenous administration | Approximately 30 min after injury, the rats were treated with MSC-EXOs transfected with miR-126 or miR-con EXOs via tail vein injection | MSC-EXOs transfected with miR-126 efficiently transferred miR-126 to the site of injury, induced angiogenesis by suppressing SPRED1 and PIK3R2, stimulated neurogenesis, and protected neuronal cell to apoptosis SCI induced. | In vivo | [107] |
| 10 μg | HUVECs OGD model exposed to hPMSCs-EXOs | First, HUVECs were incubated with miR-con EXOs or miR-126 EXOs for 6 h and subjected to OGD. Then, the cells were incubated with MSC-EXOs for 2 hours | In vitro | |||
| MSC-EXOs transfected with miR-133b | 100 μg in 0.5 mL of PBS | Intravenous administration | The animals were treated with MSC-EXOs transfected with miR-133b, 24 h after injury | Systemic administration of miR-133b-transfected EXOs upregulated miR-133b expression at the site of injury and protected neuronal cells from apoptosis and promoted axon regeneration. Part of these effects was mediated by the activation of the ERK1/2, STAT3, and CREB pathways, targeting RhoA which is instead inhibited. | In vivo | [102] |
| MSC-EXOs transfected with miR-145-5p | 100 μg in 0.5 mL of PBS, equivalent to 1 × 1010 particles | Intravenous administration | The animals were treated with MSC-EXOs transfected with miR-145-5p 30 min after injury | MSC-EXOs transfected with miR-145-5p can be useful to reduce the inflammation in SCI by the involvement of TLR4/NF-κB pathway modulation. | In vivo | [109] |
| 10 μg | PC12 cells incubated with MSCs-EXOs transfected with miR-145-5p | After 24 h, PC12 cells treated with LPS were incubated with MSCs-EXOs containing miR-145-5p for 48 h | In vitro | |||
| BMMSCs-EVs | 200 μg/mL | Intravenous administration | BMMSCs-Evs were injected into the tail vein 30 min post-SCI and 1-day post-injury | BMMSC-Evs reduced apoptosis in neuronal cells, promote regeneration, improved motor function, and attenuated the disruption of BSCB and pericyte coverage via NF-κB p65. | In vivo | [63] |
| 100 μg/mL | Pericytes co-incubation and exposure to BMSCs-Evs | Pericytes were seeded 100 μg/mL of BMMSC-Evs and after were exposed to OGD/reperfusion exposure | In vitro | |||
| BMMSC-EXOs | 200 µg/mL, derived from ∼1 × 106 MSCs | Intravenous administration | 30-min post-injury and 1 day after, the animals were infused by tail vain with BMMSCs or with BMMSC-EXOs | Both MSCs and MSC-EXOs administration exerted anti-inflammatory and neuroprotective properties attenuating SCI-induced A1 astrocytes activation via inhibiting nuclear translocation of NFκB p65 | In vivo | [77] |
| 5 × 104 | Astrocyte co-culture with BMMSCs or BMMSC-EXOs | Astrocytes collected from SCI rats co-cultured with BMMSCs or MSC-EXOs for 48 h | In vitro | |||
| BMMSC-EXOs | 200 μg of BMSCs-EXOs precipitated in 200 μL of PBS | Intravenous administration | After SCI, the rats were treated with BMSCs-EXOs via tail vain | The BMMSC-EXOs treatment induced functional behavioral recovery, enhancing blood vessel formation, reducing glial scars, protecting the neuronal cells against apoptosis, increasing axonal regeneration, and decreasing inflammatory response by suppression of the activation of A1 neurotoxic reactive astrocytes. | In vivo | [110] |
| 100 μg/mL | HUVECs exposed to BMMSCs-EXOs | 2 × 104 HUVEC cells were seeded and after co-incubated with BMMSC-EXOs for 24 h. It has been tested the effects of BMMSCs-EXOs on nitric oxide production, HUVEC cells, 1 hour before stimulation with 5 ng/mL LPS, microglia 2 × 105 cells/mL, have been pretreated with or without BMSCs-EXOs 100 μg/mL | In vitro | |||
| BMMSC-small EVs | 2.5 × 109 or 8.3 × 108 | Intravenous administration | On day 7 post-SCI, MSCs or MSC-sEVs suspended in 1 mL DMEM were infused via the femoral vein. On 8- and 9-day post-SCI, 0.2 mL DMEM alone or 1/3 dose MSC-sEVs in 0.2 mL DMEM was infused via the saphenous vein. | Both MSC infusion and fractionated MSC-smallEVs target M2 macrophages and augment TGF-β receptors, activating the TGF-β signaling pathway, thus promoting functional recovery in SCI animals. | In vivo | [111] |
| BMMSC-EXOs | 2.5 × 109 in 0.2 mL of PBS | Intravenous administration | One week post-injury, DiR labeled MSC-EXOs were infused into the saphenous veins | Intravenous administration of MSC-EXOs rapidly traffic to the injured and associate specifically with M2 macrophages. | In vivo | [83] |
| BMMSC-EXOs | 200 μg/mL | BV2 exposed to BMMSCs-EXOs under hypoxic or normoxic condition | LPS (1 μg/mL) was co-cultured with BV2 microglia for 24 h followed by the addition of EXOs | Hypoxia preconditioning represents a promising and effective approach to enhancing the therapeutic properties of MSC-EXOs, promoting functional behavioral recovery in the SCI model by shifting microglial polarization from M1 to M2 phenotype in vivo and in vitro. | In vitro | [112] |
| 200 μg of total protein of EXOs precipitated in 200 μL PBS | Intravenous administration | Mice were subjected to SCI, followed by tail vein injection of EXOs, HEXOs, miR-NCOE-HExos, miROE-HExos, miR-NCKD-HExos, and miRKD-HExos. | In vivo | |||
| N-NVs and MF-NVs | 20 μg/mL | PC12 cells and HUVECs were cultured in the media containing 500 μM of H2O2 and lipopolysaccharide in a hypoxic incubator for 24 h. To polarize macrophages into the M1 phenotype, RAW 264.7 cells were cultured in the presence of LPS for 24 h in normoxia. After, the cells were treated with N-NVs and MF-NVs (20 μg/mL) for 24 h under hypoxic and polarization culture conditions. | Compared to normal N-NVs, MF-NVs contained a larger quantity of ischemic area-targeting molecules. The MF-NVs enhanced their accumulation in the injured spinal cord. This increased reduced apoptosis and inflammation prevented the axonal loss, increased blood vessel formation, decreased fibrosis, and consequently, improved spinal cord function. | In vitro | [113] | |
| 25 μg of N-NV in 100 μL PBS, and 25 μg of MF-NV in 100 μL PBS | Intravenous administration | N-NVs and MF-NVs intravenously injected to tail vein 1 h and 7 days post-SCI | In vivo | |||
| Human MSC-EXOs immobilized in pGel | 100 μg suspended in 20 μL PBS | Topical transplantation | After the lesion, the animals were implanted with human MSCs-EXOs and injected into the pGel (60 μL) | The human MSC-EXOs immobilized in pGel induced nerve tissue repair and functional recovery protecting bladder and kidney tissues from SCI induced-neuronal damage. | In vivo | [100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schepici, G.; Silvestro, S.; Mazzon, E. Regenerative Effects of Exosomes-Derived MSCs: An Overview on Spinal Cord Injury Experimental Studies. Biomedicines 2023, 11, 201. https://doi.org/10.3390/biomedicines11010201
Schepici G, Silvestro S, Mazzon E. Regenerative Effects of Exosomes-Derived MSCs: An Overview on Spinal Cord Injury Experimental Studies. Biomedicines. 2023; 11(1):201. https://doi.org/10.3390/biomedicines11010201
Chicago/Turabian StyleSchepici, Giovanni, Serena Silvestro, and Emanuela Mazzon. 2023. "Regenerative Effects of Exosomes-Derived MSCs: An Overview on Spinal Cord Injury Experimental Studies" Biomedicines 11, no. 1: 201. https://doi.org/10.3390/biomedicines11010201
APA StyleSchepici, G., Silvestro, S., & Mazzon, E. (2023). Regenerative Effects of Exosomes-Derived MSCs: An Overview on Spinal Cord Injury Experimental Studies. Biomedicines, 11(1), 201. https://doi.org/10.3390/biomedicines11010201