
Preventive Action of Beta-Carotene against the Indoxyl Sulfate-Induced Renal Dysfunction in Male Adult Zebrafish via Regulations of Mitochondrial Inflammatory and β-Carotene Oxygenase-2 Actions

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs and Chemicals
2.3. Induction of Renal Dysfunction by IS Exposure
2.4. Experimental Protocol
- ⮚
- Group 1: This group was employed as a normal control group without exposure to any IS, BC, and CoQ10 agents.
- ⮚
- Group 2: This group was employed as an IS-exposed group (as a negative control group). The induction of renal dysfunction was made as described in the previous sections.
- ⮚
- Groups 3 and 4: This group was employed to exposure to BC doses, i.e., 50 and 100 mg/L/hour/day for 4 weeks in IS-exposed animals, respectively.
- ⮚
- Group 5: This group was employed to expose CoQ10 (20 mg/L/hour/day) for 4 weeks in IS-exposed animals. CoQ10 is a key component of the mitochondrial respiratory chain, and it acts as an endogenous antioxidant [43]. Experimentally, it prevents renal mitochondrial dysfunction and renoprotective actions [44]. Clinically, CoQ10 improves mitochondrial function and attenuates end-stage renal disease [45]. Hence, it is used as a reference drug in this study.
2.5. Estimation of BUN and Creatinine Concentration
2.6. Estimation of Renal Mitochondrial Biomarkers
2.6.1. Estimation of Renal Mitochondrial BCO2 Activity
2.6.2. Estimation of Renal Mitochondrial SOD2 Activity
2.6.3. Estimation of Renal Mitochondrial GPX1 Activity
2.6.4. Estimation of Renal Mitochondrial GSSG/GSH Ratio
2.6.5. Estimation of Renal Mitochondrial Carbonylated Protein Content
2.6.6. Estimation of Renal Mitochondrial ATP Content
2.6.7. Estimation of Renal Mitochondrial Complex I Activity
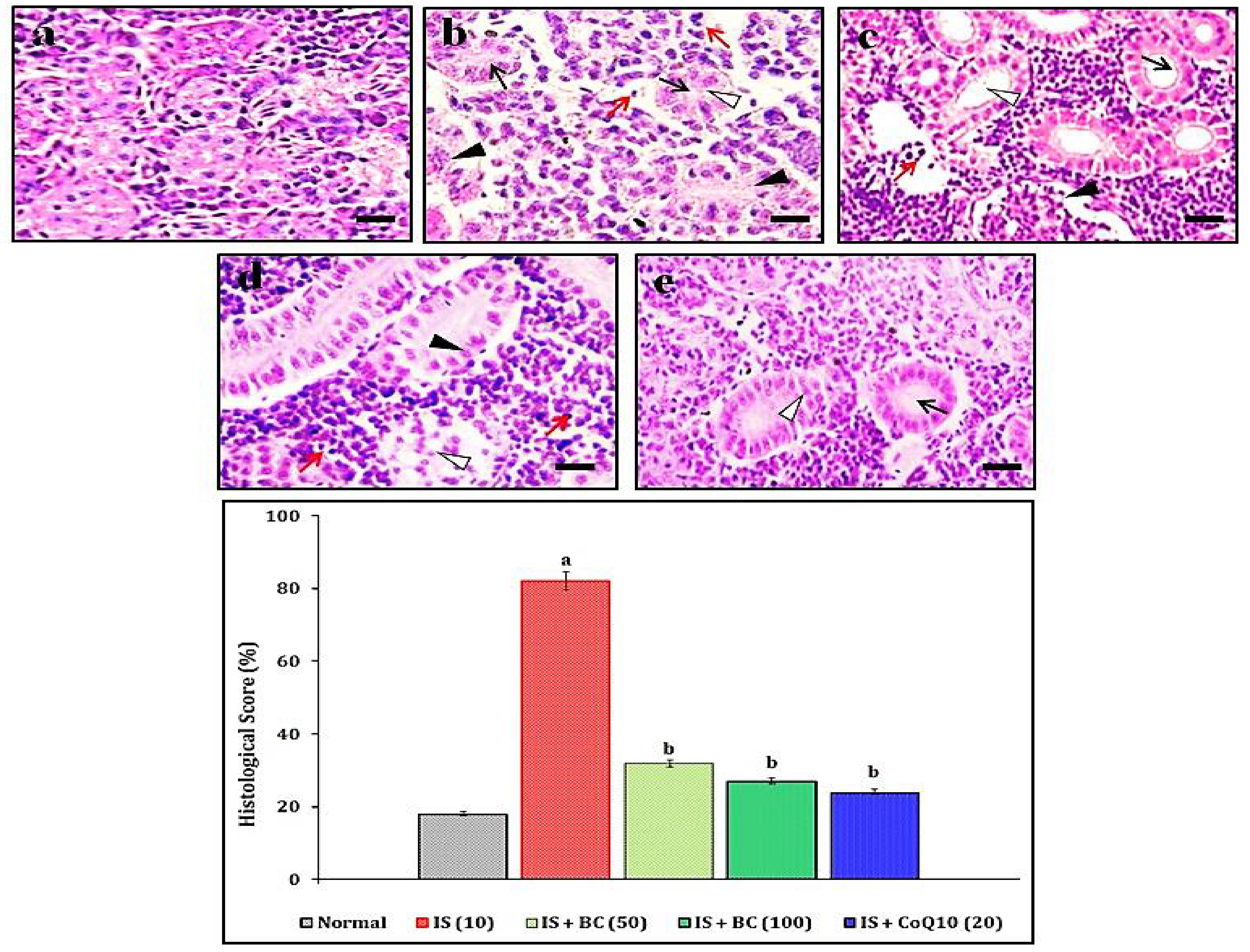
2.7. Assessment of IS-Induced Renal Histopathological Changes
2.8. Statistical Analysis
3. Results
3.1. Effect of BC on IS-Induced Blood Biomarker Changes
3.2. Effect of BC in IS-Induced Renal Tissue Biomarker Changes
3.3. Effect of BC in IS-Induced Histopathological Changes
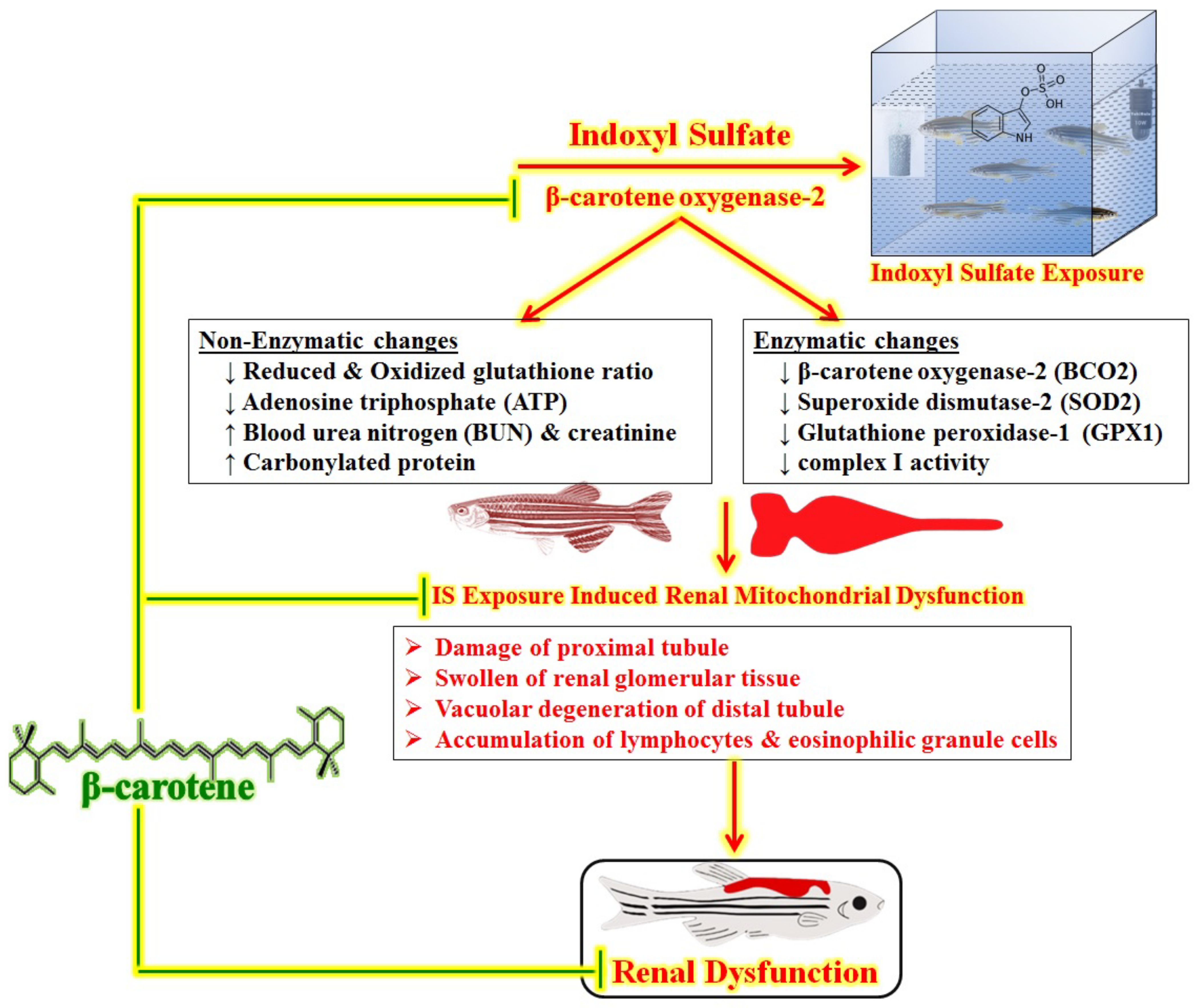
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banoglu, E.; Jha, G.G.; King, R.S. Hepatic Microsomal Metabolism of Indole to Indoxyl, a Precursor of Indoxyl Sulfate. Eur. J. Drug Metab. Pharmacokinet. 2001, 26, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Bouranis, J.A.; Beaver, L.M.; Jiang, D.; Choi, J.; Wong, C.P.; Davis, E.W.; Williams, D.E.; Sharpton, T.J.; Stevens, J.F.; Ho, E. Interplay between Cruciferous Vegetables and the Gut Microbiome: A Multi-Omic Approach. Nutrients 2023, 15, 42. [Google Scholar] [CrossRef] [PubMed]
- Hendrikx, T.; Schnabl, B. Indoles: Metabolites Produced by Intestinal Bacteria Capable of Controlling Liver Disease Manifestation. J. Intern. Med. 2019, 286, 32–40. [Google Scholar] [CrossRef]
- Lauriola, M.; Farré, R.; Evenepoel, P.; Overbeek, S.A.; Meijers, B. Food-Derived Uremic Toxins in Chronic Kidney Disease. Toxins 2023, 15, 116. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Liu, J.; Yu, W.; Li, C.; Huang, L.; Mao, W.; Lu, Z. Tryptophan Intake, Not Always the More the Better. Front. Nutr. 2023, 10, 1140054. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.; Saldanha, J.F.; Stockler-Pinto, M.B.; Fouque, D.; Soulage, C.O.; Mafra, D. Effects of Resveratrol on Inflammation and Oxidative Stress Induced by the Uremic Toxin Indoxyl Sulfate in Murine Macrophage-like RAW 264.7. Biochimie 2023, 213, 22–29. [Google Scholar] [CrossRef]
- Niwa, T.; Aoyama, I.; Takayama, F.; Tsukushi, S.; Miyazaki, T.; Owada, A.; Shiigai, T. Uremic Toxicity: Urinary Indoxyl Sulfate Is a Clinical Factor That Affects the Progression of Renal Failure. Miner. Electrolyte Metab. 1999, 25, 118–122. [Google Scholar] [CrossRef]
- Namba, S.; Okuda, Y.; Morimoto, A.; Kojima, T.; Morita, T. Serum Indoxyl Sulfate Is a Useful Predictor for Progression of Chronic Kidney Disease. Rinsho Byori. Jpn. J. Clin. Pathol. 2010, 58, 448–453. [Google Scholar]
- Popkov, V.A.; Silachev, D.N.; Zalevsky, A.O.; Zorov, D.B.; Plotnikov, E.Y. Mitochondria as a Source and a Target for Uremic Toxins. Int. J. Mol. Sci. 2019, 20, 3094. [Google Scholar] [CrossRef]
- Strychalski, J.; Gugołek, A.; Brym, P.; Antoszkiewicz, Z. Effect of the Β-carotene Oxygenase 2 Genotype on the Content of Carotenoids, Retinol and A-tocopherol in the Liver, Fat and Milk of Rabbit Does, Reproduction Parameters and Kitten Growth. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1585–1593. [Google Scholar] [CrossRef]
- Bradley, M.; Arballo, J.; Black, M.; Erdman, J.; Amengual, J. Tissue Lycopene Accumulation in Transgenic Mice Lacking One or Both Carotenoid Cleaving Enzymes. Curr. Dev. Nutr. 2022, 6, 55. [Google Scholar] [CrossRef]
- Yabuuchi, N.; Hou, H.; Gunda, N.; Narita, Y.; Jono, H.; Saito, H. Suppressed Hepatic Production of Indoxyl Sulfate Attenuates Cisplatin-Induced Acute Kidney Injury in Sulfotransferase 1a1-Deficient Mice. Int. J. Mol. Sci. 2021, 22, 1764. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.-H.; Ma, M.-C.; Liao, M.-T.; Zheng, C.-M.; Lu, K.-C.; Liao, C.-H.; Hou, Y.-C.; Liu, W.-C.; Lu, C.-L. Indoxyl Sulfate, a Tubular Toxin, Contributes to the Development of Chronic Kidney Disease. Toxins 2020, 12, 684. [Google Scholar] [CrossRef] [PubMed]
- Lano, G.; Burtey, S.; Sallée, M. Indoxyl Sulfate, a Uremic Endotheliotoxin. Toxins 2020, 12, 229. [Google Scholar] [CrossRef]
- Hobson, S.; Arefin, S.; Rahman, A.; Hernandez, L.; Ebert, T.; de Loor, H.; Evenepoel, P.; Stenvinkel, P.; Kublickiene, K. Indoxyl Sulphate Retention Is Associated with Microvascular Endothelial Dysfunction after Kidney Transplantation. Int. J. Mol. Sci. 2023, 24, 3640. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.W.-H.; Wu, P.-H.; Lin, Y.-T.; Chiu, C.-H.; Cheng, T.-L.; Guan, W.-H.; Lin, H.Y.-H.; Lee, K.-T.; Chen, Y.-H.; Chiu, C.-C. Zebrafish Model-Based Assessment of Indoxyl Sulfate-Induced Oxidative Stress and Its Impact on Renal and Cardiac Development. Antioxidants 2022, 11, 400. [Google Scholar] [CrossRef]
- Zgoda-Pols, J.R.; Chowdhury, S.; Wirth, M.; Milburn, M.V.; Alexander, D.C.; Alton, K.B. Metabolomics Analysis Reveals Elevation of 3-Indoxyl Sulfate in Plasma and Brain during Chemically-Induced Acute Kidney Injury in Mice: Investigation of Nicotinic Acid Receptor Agonists. Toxicol. Appl. Pharmacol. 2011, 255, 48–56. [Google Scholar] [CrossRef]
- Di Paola, R.; De, A.; Izhar, R.; Abate, M.; Zappavigna, S.; Capasso, A.; Perna, A.F.; La Russa, A.; Capasso, G.; Caraglia, M. Possible Effects of Uremic Toxins P-Cresol, Indoxyl Sulfate, P-Cresyl Sulfate on the Development and Progression of Colon Cancer in Patients with Chronic Renal Failure. Genes 2023, 14, 1257. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, S.; Wu, Q.-J.; Xiao, J.; Wang, Z.-H.; Mu, X.-W.; Zhang, Y.; Wang, X.-N.; You, L.-L.; Wang, S.-N.; et al. Serum Total Indoxyl Sulfate Levels and All-Cause and Cardiovascular Mortality in Maintenance Hemodialysis Patients: A Prospective Cohort Study. BMC Nephrol. 2022, 23, 231. [Google Scholar] [CrossRef]
- Caggiano, G.; Amodio, L.; Stasi, A.; Colabufo, N.A.; Colangiulo, S.; Pesce, F.; Gesualdo, L. Gut-Derived Uremic Toxins in CKD: An Improved Approach for the Evaluation of Serum Indoxyl Sulfate in Clinical Practice. Int. J. Mol. Sci. 2023, 24, 5142. [Google Scholar] [CrossRef]
- Berg, A.H.; Kumar, S.; Karumanchi, S.A. Indoxyl Sulfate in Uremia: An Old Idea with Updated Concepts. J. Clin. Investig. 2022, 132, e155860. [Google Scholar] [CrossRef] [PubMed]
- Alhassani, R.Y.; Bagadood, R.M.; Balubaid, R.N.; Barno, H.I.; Alahmadi, M.O.; Ayoub, N.A. Drug Therapies Affecting Renal Function: An Overview. Cureus 2021, 13, e19924. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.; Jaeger, S.U.; Filler, G. Drug Use, Dosing, and Toxicity in Kidney Disease. In Pediatric Kidney Disease; Springer: Berlin/Heidelberg, Germany, 2023; pp. 1965–1985. [Google Scholar]
- Aloy, B.; Launay-Vacher, V.; Bleibtreu, A.; Bortolotti, P.; Faure, E.; Filali, A.; Gauzit, R.; Gilbert, M.; Lesprit, P.; Mahieu, R.; et al. Antibiotics and Chronic Kidney Disease: Dose Adjustment Update for Infectious Disease Clinical Practice. Med. Mal. Infect. 2020, 50, 323–331. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, C. Renal Disease. Fluid and Electrolyte Disorders. In Medicine for Finals and Beyond; CRC Press: Boca Raton, FL, USA, 2023; pp. 277–330. ISBN 1003193617. [Google Scholar]
- Wiles, K.; Skinner, L.; Smith, P. Renal Disease in Pregnancy. Medicine 2023, 51, 293–299. [Google Scholar] [CrossRef]
- Corridon, P.R. Still Finding Ways to Augment the Existing Management of Acute and Chronic Kidney Diseases with Targeted Gene and Cell Therapies: Opportunities and Hurdles. Front. Med. 2023, 10, 1143028. [Google Scholar] [CrossRef]
- Santhosh, S.; Pazhani, G.P.; Arathi, M.S.; Manickam, S. Nephroprotective Role of Boerhavia Diffusa in Renal Disorders: A Review. Res. J. Pharm. Technol. 2023, 16, 962–968. [Google Scholar]
- Uroko, R.I.; Uche, M.E.; Nweje-Anyalowu, P.C.; Obiwuru, I.; Aguwamba, C.; Aaron, C.F. Combined Anthocleista Vogelii and Alstonia Boonei Stem Barks Extract Alleviates Hyperlipidaemia and Renal Malfunctions in Benign Prostatic Hyperplasia-Induced Rats. Biol. Med. Nat. Prod. Chem. 2023, 12, 69–78. [Google Scholar] [CrossRef]
- Peng, A.; Gu, Y.; Lin, S.Y. Herbal Treatment for Renal Diseases. Ann. Acad. Med. Singap. 2005, 34, 44–51. [Google Scholar] [CrossRef]
- Rather, I.A.; Khan, N.; Kushwah, A.S.; Surampalli, G.; Kumar, M. Nephroprotective Effects of Honokiol in a High-Fat Diet-Streptozotocin Rat Model of Diabetic Nephropathy. Life Sci. 2023, 320, 121543. [Google Scholar] [CrossRef]
- Basist, P.; Parveen, B.; Zahiruddin, S.; Gautam, G.; Parveen, R.; Khan, M.A.; Krishnan, A.; Shahid, M.; Ahmad, S. Potential Nephroprotective Phytochemicals: Mechanism and Future Prospects. J. Ethnopharmacol. 2022, 283, 114743. [Google Scholar] [CrossRef]
- Atteia, H.H.; Alamri, E.S.; Sirag, N.; Zidan, N.S.; Aljohani, R.H.; Alzahrani, S.; Arafa, M.H.; Mohammad, N.S.; Asker, M.E.; Zaitone, S.A. Soluble Guanylate Cyclase Agonist, Isoliquiritigenin Attenuates Renal Damage and Aortic Calcification in a Rat Model of Chronic Kidney Failure. Life Sci. 2023, 317, 121460. [Google Scholar] [CrossRef] [PubMed]
- Pieniazek, A.; Bernasinska-Slomczewska, J.; Gwozdzinski, L. Uremic Toxins and Their Relation with Oxidative Stress Induced in Patients with CKD. Int. J. Mol. Sci. 2021, 22, 6196. [Google Scholar] [CrossRef] [PubMed]
- Paramaswaran, Y.; Subramanian, A.; Paramakrishnan, N.; Ramesh, M.; Muthuraman, A. Therapeutic Investigation of Palm Oil Mill Effluent-Derived Beta-Carotene in Streptozotocin-Induced Diabetic Retinopathy via the Regulation of Blood–Retina Barrier Functions. Pharmaceuticals 2023, 16, 647. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.-Q.; Liu, C.; Ernst, H.; Krinsky, N.I.; Russell, R.M.; Wang, X.-D. The Biochemical Characterization of Ferret Carotene-9′, 10′-Monooxygenase Catalyzing Cleavage of Carotenoids in Vitro and in Vivo. J. Biol. Chem. 2006, 281, 19327–19338. [Google Scholar] [CrossRef]
- Jin, Y.; Yu, Y.; Zhang, C.; Li, S.; Zhang, X.; Li, F. Characterization and Function Analysis of the Beta-Carotene Oxygenase-like Genes in Carotenoids Metabolism of the Ridgetail White Prawn Exopalaemon Carinicauda. Front. Physiol. 2020, 11, 745. [Google Scholar] [CrossRef]
- Wu, L.; Lu, P.; Guo, X.; Song, K.; Lyu, Y.; Bothwell, J.; Wu, J.; Hawkins, O.; Clarke, S.L.; Lucas, E.A.; et al. β-Carotene Oxygenase 2 Deficiency-Triggered Mitochondrial Oxidative Stress Promotes Low-Grade Inflammation and Metabolic Dysfunction. Free Radic. Biol. Med. 2021, 164, 271–284. [Google Scholar] [CrossRef]
- Hosseini, F.; Gharib Naseri, M.K.; Badavi, M.; Ghaffari, M.A.; Shahbazian, H.; Rashidi, I. Protective Effect of Beta Carotene Pretreatment on Renal Ischemia/reperfusion Injury in Rat. Pak. J. Biol. Sci. 2009, 12, 1140–1145. [Google Scholar] [CrossRef]
- Nakano, T.; Tanaka, S.; Tsuruya, K.; Kitazono, T. Low Intake of β Carotene and Dietary Fiber from Vegetables and Fruits in Patients with Chronic Kidney Disease. Sci. Rep. 2022, 12, 19953. [Google Scholar] [CrossRef]
- Cleveland, K.H.; Schnellmann, R.G. Pharmacological Targeting of Mitochondria in Diabetic Kidney Disease. Pharmacol. Rev. 2023, 75, 250–262. [Google Scholar] [CrossRef]
- Lee, W.C.; Li, L.C.; Chen, J.B.; Chang, H.W. Indoxyl Sulfate-Induced Oxidative Stress, Mitochondrial Dysfunction, and Impaired Biogenesis Are Partly Protected by Vitamin C and N-Acetylcysteine. Sci. World J. 2015, 2015, 620826. [Google Scholar] [CrossRef]
- Hidalgo-Gutiérrez, A.; González-García, P.; Díaz-Casado, M.E.; Barriocanal-Casado, E.; López-Herrador, S.; Quinzii, C.M.; López, L.C. Metabolic Targets of Coenzyme Q10 in Mitochondria. Antioxidants 2021, 10, 520. [Google Scholar] [CrossRef] [PubMed]
- Sourris, K.C.; Harcourt, B.E.; Tang, P.H.; Morley, A.L.; Huynh, K.; Penfold, S.A.; Coughlan, M.T.; Cooper, M.E.; Nguyen, T.-V.; Ritchie, R.H.; et al. Ubiquinone (Coenzyme Q10) Prevents Renal Mitochondrial Dysfunction in an Experimental Model of Type 2 Diabetes. Free Radic. Biol. Med. 2012, 52, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, J.; Han, E.; Wang, Y.; Gao, J. Efficacy of Coenzyme Q10 in Patients with Chronic Kidney Disease: Protocol for a Systematic Review. BMJ Open 2019, 9, e029053. [Google Scholar] [CrossRef] [PubMed]
- Babaei, F.; Ramalingam, R.; Tavendale, A.; Liang, Y.; Yan, L.S.; Ajuh, P.; Cheng, S.H.; Lam, Y.W. Novel Blood Collection Method Allows Plasma Proteome Analysis from Single Zebrafish. J. Proteome Res. 2013, 12, 1580–1590. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, G.F.; Schrader, L.N.; Wingert, R.A. Dissection of the Adult Zebrafish Kidney. J. Vis. Exp. JoVE 2011, 54, e2839. [Google Scholar] [CrossRef]
- Gross, V.S.; Greenberg, H.K.; Baranov, S.V.; Carlson, G.M.; Stavrovskaya, I.G.; Lazarev, A.V.; Kristal, B.S. Isolation of Functional Mitochondria from Rat Kidney and Skeletal Muscle without Manual homogenization. Anal. Biochem. 2011, 418, 213–223. [Google Scholar] [CrossRef]
- Babino, D.; Palczewski, G.; Widjaja-Adhi, M.A.K.; Kiser, P.D.; Golczak, M.; von Lintig, J. Characterization of the Role of β-Carotene 9, 10-Dioxygenase in Macular Pigment Metabolism. J. Biol. Chem. 2015, 290, 24844–24857. [Google Scholar] [CrossRef]
- Thomas, L.D.; Bandara, S.; Parmar, V.M.; Srinivasagan, R.; Khadka, N.; Golczak, M.; Kiser, P.D.; von Lintig, J. The Human Mitochondrial Enzyme BCO2 Exhibits Catalytic Activity toward Carotenoids and Apocarotenoids. J. Biol. Chem. 2020, 295, 15553–15565. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Glutathione Peroxidase Activity in Selenium-Deficient Rat Liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Dringen, R.; Hamprecht, B. Glutathione Content as an Indicator for the Presence of Metabolic Pathways of Amino Acids in Astroglial Cultures. J. Neurochem. 1996, 67, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Hays, A.M.; Lantz, R.C.; Witten, M.L. Correlation between in Vivo and in Vitro Pulmonary Responses to Jet Propulsion Fuel-8 Using Precision-Cut Lung Slices and a Dynamic Organ Culture System. Toxicol. Pathol. 2003, 31, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- McCampbell, K.; Springer, K.; Wingert, R. Atlas of Cellular Dynamics during Zebrafish Adult Kidney Regeneration. Stem Cells Int. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- McKee, R.A.; Wingert, R.A. Zebrafish Renal Pathology: Emerging Models of Acute Kidney Injury. Curr. Pathobiol. Rep. 2015, 3, 171–181. [Google Scholar] [CrossRef]
- Yao, X.; Liu, Y.; Yang, Y.; Li, Y.; Hu, N.; Song, F.; Yang, F. Microcystin-LR-Exposure-Induced Kidney Damage by Inhibiting MKK6-Mediated Mitophagy in Mice. Toxins 2023, 15, 404. [Google Scholar] [CrossRef]
- Mantle, D.; Heaton, R.A.; Hargreaves, I.P. Coenzyme Q10 and Immune Function: An Overview. Antioxidants 2021, 10, 759. [Google Scholar] [CrossRef]
- Ferreira-Cravo, M.; Moreira, D.C.; Hermes-Lima, M. Glutathione Depletion Disrupts Redox Homeostasis in an Anoxia-Tolerant Invertebrate. Antioxidants 2023, 12, 1197. [Google Scholar] [CrossRef]
- Nesci, S.; Spagnoletta, A.; Oppedisano, F. Inflammation, Mitochondria and Natural Compounds Together in the Circle of Trust. Int. J. Mol. Sci. 2023, 24, 6106. [Google Scholar] [CrossRef]
- Khan, A.U.H.; Allende-Vega, N.; Gitenay, D.; Garaude, J.; Vo, D.-N.; Belkhala, S.; Gerbal-Chaloin, S.; Gondeau, C.; Daujat-Chavanieu, M.; Delettre, C.; et al. Mitochondrial Complex I Activity Signals Antioxidant Response through ERK5. Sci. Rep. 2018, 8, 7420. [Google Scholar] [CrossRef]
- Trigo, D.; Avelar, C.; Fernandes, M.; Sá, J.; da Cruz e Silva, O. Mitochondria, Energy, and Metabolism in Neuronal Health and Disease. FEBS Lett. 2022, 596, 1095–1110. [Google Scholar] [CrossRef] [PubMed]
- Hammerling, U.; Kim, Y.-K.; Quadro, L. Quantum Chemistry Rules Retinoid Biology. Commun. Biol. 2023, 6, 227. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.E.; Ramkumar, S.; Sun, W.; Colon Ortiz, C.; Kiser, P.D.; Golczak, M.; von Lintig, J. The Biochemical Basis of Vitamin A Production from the Asymmetric Carotenoid β-Cryptoxanthin. ACS Chem. Biol. 2018, 13, 2121–2129. [Google Scholar] [CrossRef] [PubMed]
- Teixé-Roig, J.; Oms-Oliu, G.; Odriozola-Serrano, I.; Martín-Belloso, O. Effect of the Emulsifier Used in Dunaliella salina-Based Nanoemulsions Formulation on the β-Carotene Absorption and Metabolism in Rats. Mol. Nutr. Food Res. 2023, 67, 2200492. [Google Scholar] [CrossRef] [PubMed]
- Johra, F.T.; Bepari, A.K.; Bristy, A.T.; Reza, H.M. A Mechanistic Review of β-Carotene, Lutein, and Zeaxanthin in Eye Health and Disease. Antioxidants 2020, 9, 1046. [Google Scholar] [CrossRef]
- Våge, D.I.; Boman, I.A. A Nonsense Mutation in the Beta-Carotene Oxygenase 2 (BCO2) Gene Is Tightly Associated with Accumulation of Carotenoids in Adipose Tissue in Sheep (Ovis Aries). BMC Genet. 2010, 11, 10. [Google Scholar] [CrossRef]
- Lim, J.Y.; Liu, C.; Hu, K.-Q.; Smith, D.E.; Wang, X.-D. Ablation of Carotenoid Cleavage Enzymes (BCO1 and BCO2) Induced Hepatic Steatosis by Altering the Farnesoid X receptor/miR-34a/sirtuin 1 Pathway. Arch. Biochem. Biophys. 2018, 654, 1–9. [Google Scholar] [CrossRef]
- Chiaverelli, R.A.; Hu, K.-Q.; Liu, C.; Lim, J.Y.; Daniels, M.S.; Xia, H.; Mein, J.; von Lintig, J.; Wang, X.-D. β-Cryptoxanthin Attenuates Cigarette-Smoke-Induced Lung Lesions in the Absence of Carotenoid Cleavage Enzymes (BCO1/BCO2) in Mice. Molecules 2023, 28, 1383. [Google Scholar] [CrossRef]
- Dulińska-Litewka, J.; Sharoni, Y.; Hałubiec, P.; Łazarczyk, A.; Szafrański, O.; McCubrey, J.A.; Gąsiorkiewicz, B.; Laidler, P.; Bohn, T. Recent Progress in Discovering the Role of Carotenoids and Their Metabolites in Prostatic Physiology and Pathology with a Focus on Prostate Cancer—A Review—Part I: Molecular Mechanisms of Carotenoid Action. Antioxidants 2021, 10, 585. [Google Scholar] [CrossRef]
- Lindqvist, A.; He, Y.-G.; Andersson, S. Cell Type-Specific Expression of β-Carotene 9′, 10′-monooxygenase in Human Tissues. J. Histochem. Cytochem. 2005, 53, 1403–1412. [Google Scholar] [CrossRef]
- Ismaiel, M.M.S.; Piercey-Normore, M.D. Antioxidant Enzymes of Pseudochlorella Pringsheimii under Two Stressors: Variation of SOD Isoforms Activity. J. Plant Res. 2023, 136, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, A.; Lalanne, K.; Vianna, F.; Perrot, Y.; Richaud, M.; SenGupta, T.; Cardot-Martin, M.; Pedini, P.; Picard, C.; Nilsen, H. Targeted Central Nervous System Irradiation with Proton Microbeam Induces Mitochondrial Changes in Caenorhabditis Elegans. Biology 2023, 12, 839. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.A.; Park, C.W. Catalytic Antioxidants in the Kidney. Antioxidants 2021, 10, 130. [Google Scholar] [CrossRef]
- Esposito, L.A.; Kokoszka, J.E.; Waymire, K.G.; Cottrell, B.; MacGregor, G.R.; Wallace, D.C. Mitochondrial Oxidative Stress in Mice Lacking the Glutathione Peroxidase-1 Gene. Free Radic. Biol. Med. 2000, 28, 754–766. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, L.J.H.C.; Riemer, J. Maintenance of Small Molecule Redox Homeostasis in Mitochondria. FEBS Lett. 2023, 597, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Dragicevic, B.; Suvakov, S.; Jerotic, D.; Reljic, Z.; Djukanović, L.; Zelen, I.; Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Simic, T.; Dragicevic, D.; et al. Association of SOD2 (rs4880) and GPX1 (rs1050450) Gene Polymorphisms with Risk of Balkan Endemic Nephropathy and Its Related Tumors. Medicina 2019, 55, 435. [Google Scholar] [CrossRef]
- Fu, H.; Liu, H.; Ge, Y.; Chen, Y.; Tan, P.; Bai, J.; Dai, Z.; Yang, Y.; Wu, Z. Chitosan Oligosaccharide Alleviates and Removes the Toxicological Effects of Organophosphorus Pesticide Chlorpyrifos Residues. J. Hazard. Mater. 2023, 446, 130669. [Google Scholar] [CrossRef]
- Kyriakidis, N.C.; Cobo, G.; Dai, L.; Lindholm, B.; Stenvinkel, P. Role of Uremic Toxins in Early Vascular Ageing and Calcification. Toxins 2021, 13, 26. [Google Scholar] [CrossRef]
- Zanella, L.; Vianello, F. Potential of Microalgae as Functional Foods Applied to Mitochondria Protection and Healthy Aging Promotion. Nutraceuticals 2023, 3, 119–152. [Google Scholar] [CrossRef]
- Kalani, A.; Chaturvedi, P.; Brunetti, D.; Kalani, K. Mitochondrial Therapy in Neurodegeneration. Front. Pharmacol. 2023, 14, 1144093. [Google Scholar] [CrossRef] [PubMed]
- Marí, M.; de Gregorio, E.; de Dios, C.; Roca-Agujetas, V.; Cucarull, B.; Tutusaus, A.; Morales, A.; Colell, A. Mitochondrial Glutathione: Recent Insights and Role in Disease. Antioxidants 2020, 9, 909. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Lerman, A.; Lerman, L.O. The Emerging Role of Mitochondrial Targeting in Kidney Disease. Handb. Exp. Pharmacol. 2017, 240, 229–250. [Google Scholar] [CrossRef] [PubMed]
- Thome, T.; Coleman, M.D.; Ryan, T.E. Mitochondrial Bioenergetic and Proteomic Phenotyping Reveals Organ-Specific Consequences of Chronic Kidney Disease in Mice. Cells 2021, 10, 3282. [Google Scholar] [CrossRef]
- Ho, H.-J.; Shirakawa, H. Oxidative Stress and Mitochondrial Dysfunction in Chronic Kidney Disease. Cells 2023, 12, 88. [Google Scholar] [CrossRef]
- Vašková, J.; Kočan, L.; Vaško, L.; Perjési, P. Glutathione-Related Enzymes and Proteins: A Review. Molecules 2023, 28, 1447. [Google Scholar] [CrossRef]
- Jaja-Chimedza, A.; Gantar, M.; Mayer, G.D.; Gibbs, P.D.L.; Berry, J.P. Effects of Cyanobacterial Lipopolysaccharides from Microcystis on Glutathione-Based Detoxification Pathways in the Zebrafish (Danio rerio) Embryo. Toxins 2012, 4, 390–404. [Google Scholar] [CrossRef]
- Yang, L.-L.; Wang, G.-Q.; Yang, L.-M.; Huang, Z.-B.; Zhang, W.-Q.; Yu, L.-Z. Endotoxin Molecule Lipopolysaccharide-Induced Zebrafish Inflammation Model: A Novel Screening Method for Anti-Inflammatory Drugs. Molecules 2014, 19, 2390–2409. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Ferrington, D.A.; Kannan, R. Glutathione Metabolism and the Novel Role of Mitochondrial GSH in Retinal Degeneration. Antioxidants 2021, 10, 661. [Google Scholar] [CrossRef]
- Siems, W.; Sommerburg, O.; Schild, L.; Augustin, W.; Langhans, C.-D.; Wiswedel, I. β-Carotene Cleavage Products Induce Oxidative Stress in Vitro by Impairing Mitochondrial Respiration. FASEB J. 2002, 16, 1289–1291. [Google Scholar] [CrossRef]
- Peoples, J.N.; Saraf, A.; Ghazal, N.; Pham, T.T.; Kwong, J.Q. Mitochondrial Dysfunction and Oxidative Stress in Heart Disease. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The Importance of Antioxidants Which Play the Role in Cellular Response against Oxidative/nitrosative Stress: Current State. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Akaboshi, T.; Yamanishi, R. Certain Carotenoids Enhance the Intracellular Glutathione Level in a Murine Cultured Macrophage Cell Line by Inducing Glutamate-Cysteine-Ligase. Mol. Nutr. Food Res. 2014, 58, 1291–1300. [Google Scholar] [CrossRef]
- Nyariki, J.N.; Thuita, J.K.; Wambugu, A.M.; Nyamweya, N.O.; Rashid, K.; Nyambati, G.K.; Isaac, A.O. Coenzyme Q10 and Endogenous Antioxidants Neuro-Protect Mice Brain against Deleterious Effects of Melarsoprol and Trypanasoma Brucei Rhodesiense. J. Nat. Sci. Res 2018, 8, 61–73. [Google Scholar]
- Yu, J.H.; Lim, S.W.; Luo, K.; Cui, S.; Quan, Y.; Shin, Y.J.; Lee, K.E.; Kim, H.L.; Ko, E.J.; Chung, B.H.; et al. Coenzyme Q(10) Alleviates Tacrolimus-Induced Mitochondrial Dysfunction in Kidney. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 12288–12298. [Google Scholar] [CrossRef] [PubMed]
- Fazal, Y.; Fatima, S.N.; Shahid, S.M.; Mahboob, T. Nephroprotective Effects of B-Carotene on ACE Gene Expression, Oxidative Stress and Antioxidant Status in Thioacetamide Induced Renal Toxicity in Rats. Pak. J. Pharm. Sci. 2016, 29, 1139–1144. [Google Scholar]
- Santamarina, A.B.; de Souza Mesquita, L.M.; Casagrande, B.P.; Sertorio, M.N.; Vitor de Souza, D.; Mennitti, L.V.; Ribeiro, D.A.; Estadella, D.; Ventura, S.P.M.; de Rosso, V.V.; et al. Supplementation of Carotenoids from Peach Palm Waste (Bactris Gasipaes) Obtained with an Ionic Liquid Mediated Process Displays Kidney Anti-Inflammatory and Antioxidant Outcomes. Food Chem. X 2022, 13, 100245. [Google Scholar] [CrossRef]
- Gwozdzinski, K.; Pieniazek, A.; Gwozdzinski, L. Reactive Oxygen Species and Their Involvement in Red Blood Cell Damage in Chronic Kidney Disease. Oxidative Med. Cell. Longev. 2021, 2021, 6639199. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Groups | BUN (mM/L) | Creatinine (µM/L) |
|---|---|---|
| Normal | 6.24 ± 0.4 | 20.19 ± 0.7 |
| IS (10) | 12.83 ± 0.9 a | 39.72 ± 0.8 a |
| IS + BC (50) | 8.17 ± 0.8 b | 28.81 ± 0.5 b |
| IS + BC (100) | 7.36 ± 0.6 b | 25.71 ± 1.1 b |
| IS + CoQ10 (20) | 6.94 ± 0.3 b | 21.26 ± 0.9 b |
| Groups | BCO2 (nM/mg of Protein) | SOD2 (ng/mg of Protein) | GPX1 (U/mg of Protein) | GSH/GSSG Ratio | Carbonylated Proteins (nM/mg of Protein) |
|---|---|---|---|---|---|
| Normal | 1.98 ± 0.2 | 49.29 ± 2.1 | 3.61 ± 0.8 | 8.23 ± 0.6 | 1.91 ± 0.9 |
| IS (10) | 0.13 ± 0.4 a | 13.18 ± 1.5 a | 1.02 ± 1.2 a | 1.79 ± 1.2 a | 10.54 ± 0.8 a |
| IS + BC (50) | 1.06 ± 0.3 b | 31.47 ± 1.9 b | 2.71 ± 0.7 b | 4.01 ± 0.8 b | 4.59 ± 0.7 b |
| IS + BC (100) | 1.37 ± 0.5 b | 37.82 ± 1.1 b | 3.14 ± 0.9 b | 3.27 ± 0.6 b | 3.87 ± 0.4 b |
| IS + CoQ10 (20) | 1.63 ± 0.6 b | 45.39 ± 1.3 b | 3.45 ± 0.6 b | 2.36 ± 0.9 b | 2.61 ± 0.3 b |
| Groups | ATP (nM/mg of Protein) | Complex I Activity (µM/Minute/mg of Protein) |
|---|---|---|
| Normal | 5.73 ± 0.6 | 47.28 ± 1.2 |
| IS (10) | 1.26 ± 0.3 a | 12.04 ± 1.4 a |
| IS + BC (50) | 3.47 ± 0.8 b | 36.56 ± 0.9 b |
| IS + BC (100) | 4.62 ± 0.5 b | 40.28 ± 1.5 b |
| IS + CoQ10 (20) | 5.41 ± 0.7 b | 43.91 ± 1.3 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muthuraman, A.; Sayem, A.S.M.; Meenakshisundaram, S.; Ali, N.; Ahmad, S.F.; AlAsmari, A.F.; Nishat, S.; Lim, K.G.; Paramaswaran, Y. Preventive Action of Beta-Carotene against the Indoxyl Sulfate-Induced Renal Dysfunction in Male Adult Zebrafish via Regulations of Mitochondrial Inflammatory and β-Carotene Oxygenase-2 Actions. Biomedicines 2023, 11, 2654. https://doi.org/10.3390/biomedicines11102654
Muthuraman A, Sayem ASM, Meenakshisundaram S, Ali N, Ahmad SF, AlAsmari AF, Nishat S, Lim KG, Paramaswaran Y. Preventive Action of Beta-Carotene against the Indoxyl Sulfate-Induced Renal Dysfunction in Male Adult Zebrafish via Regulations of Mitochondrial Inflammatory and β-Carotene Oxygenase-2 Actions. Biomedicines. 2023; 11(10):2654. https://doi.org/10.3390/biomedicines11102654
Chicago/Turabian StyleMuthuraman, Arunachalam, Abu Sadat Md. Sayem, Sakthiganapathi Meenakshisundaram, Nemat Ali, Sheikh F. Ahmad, Abdullah F. AlAsmari, Shamama Nishat, Khian Giap Lim, and Yamunna Paramaswaran. 2023. "Preventive Action of Beta-Carotene against the Indoxyl Sulfate-Induced Renal Dysfunction in Male Adult Zebrafish via Regulations of Mitochondrial Inflammatory and β-Carotene Oxygenase-2 Actions" Biomedicines 11, no. 10: 2654. https://doi.org/10.3390/biomedicines11102654
APA StyleMuthuraman, A., Sayem, A. S. M., Meenakshisundaram, S., Ali, N., Ahmad, S. F., AlAsmari, A. F., Nishat, S., Lim, K. G., & Paramaswaran, Y. (2023). Preventive Action of Beta-Carotene against the Indoxyl Sulfate-Induced Renal Dysfunction in Male Adult Zebrafish via Regulations of Mitochondrial Inflammatory and β-Carotene Oxygenase-2 Actions. Biomedicines, 11(10), 2654. https://doi.org/10.3390/biomedicines11102654