
Opposite Effects of mRNA-Based and Adenovirus-Vectored SARS-CoV-2 Vaccines on Regulatory T Cells: A Pilot Study

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
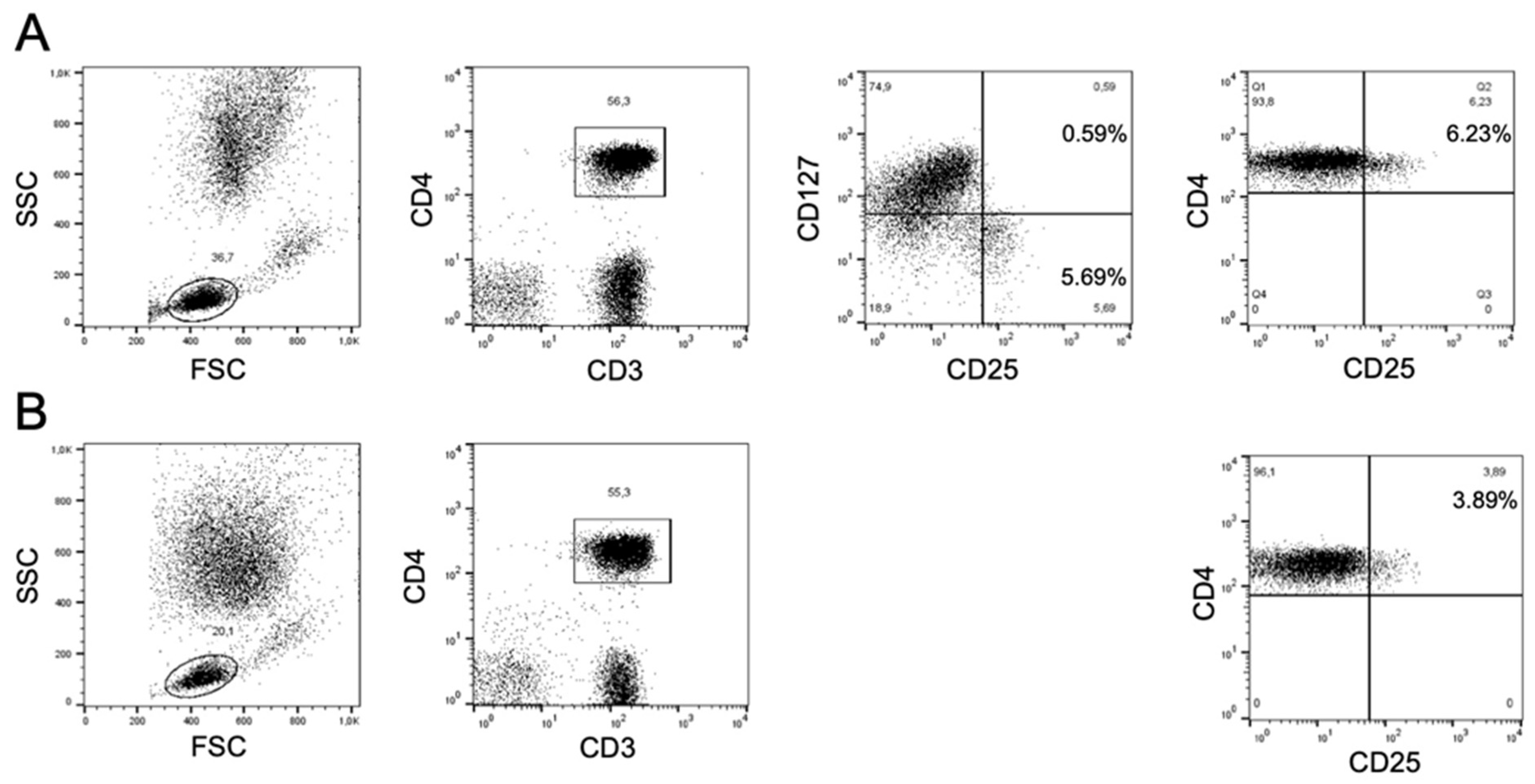
2.2. Flow Cytometry and Gating Strategy
2.3. Anti-SARS-CoV-2 Serology
2.4. Statistical Analysis
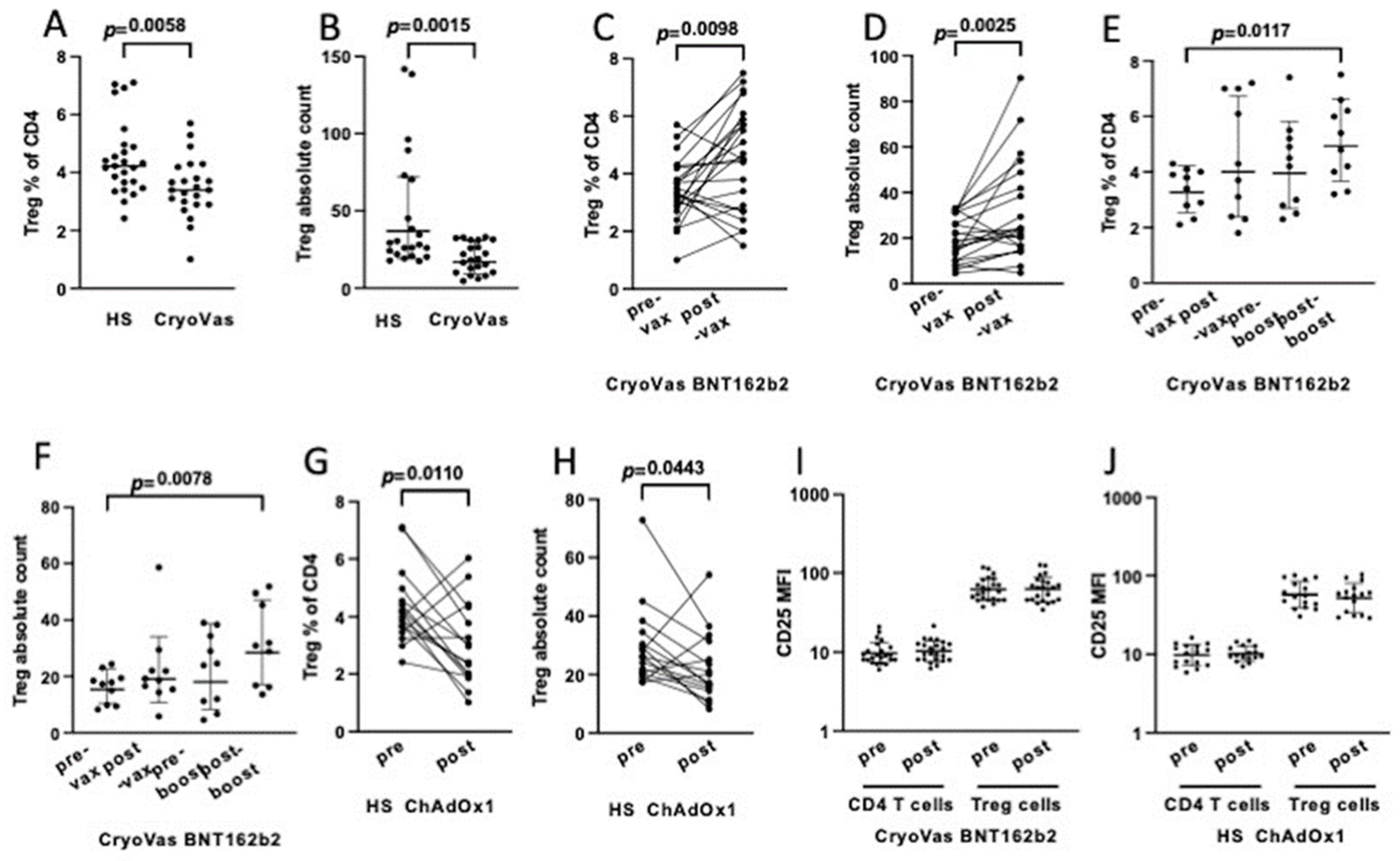
3. Results
4. Discussion
5. Limitations of the Study
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 COVID-19 Vaccine over 6 Months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef] [PubMed]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Milne, G.; Hames, T.; Scotton, C.; Gent, N.; Johnsen, A.; Anderson, R.M.; Ward, T. Does infection with or vaccination against SARS-CoV-2 lead to lasting immunity? Lancet. Respir. Med. 2021, 9, 1450–1466. [Google Scholar] [CrossRef] [PubMed]
- Feikin, D.R.; Higdon, M.M.; Abu-Raddad, L.J.; Andrews, N.; Araos, R.; Goldberg, Y.; Groome, M.J.; Huppert, A.; O’Brien, K.L.; Smith, P.G.; et al. Duration of effectiveness of vaccines against SARS-CoV-2 infection and COVID-19 disease: Results of a systematic review and meta-regression. Lancet 2022, 399, 924–944. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Sachdeva, R.; Gower, C.; Ramsay, M.; Lopez Bernal, J. Effectiveness of COVID-19 booster vaccines against COVID-19-related symptoms, hospitalization and death in England. Nat. Med. 2022, 28, 831–837. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Schmidt, T.; Klemis, V.; Schub, D.; Mihm, J.; Hielscher, F.; Marx, S.; Abu-Omar, A.; Ziegler, L.; Guckelmus, C.; Urschel, R.; et al. Immunogenicity and reactogenicity of heterologous ChAdOx1 nCoV-19/mRNA vaccination. Nat. Med. 2021, 27, 1530–1535. [Google Scholar] [CrossRef]
- Atmar, R.L.; Lyke, K.E.; Deming, M.E.; Jackson, L.A.; Branche, A.R.; El Sahly, H.M.; Rostad, C.A.; Martin, J.M.; Johnston, C.; Rupp, R.E.; et al. Homologous and Heterologous COVID-19 Booster Vaccinations. N. Engl. J. Med. 2022, 386, 1046–1057. [Google Scholar] [CrossRef]
- Liu, X.; Shaw, R.H.; Stuart, A.S.V.; Greenland, M.; Aley, P.K.; Andrews, N.J.; Cameron, J.C.; Charlton, S.; Clutterbuck, E.A.; Collins, A.M.; et al. Safety and immunogenicity of heterologous versus homologous prime-boost schedules with an adenoviral vectored and mRNA COVID-19 vaccine (Com-COV): A single-blind, randomised, non-inferiority trial. Lancet 2021, 398, 856–869. [Google Scholar] [CrossRef]
- Stuart, A.S.V.; Shaw, R.H.; Liu, X.; Greenland, M.; Aley, P.K.; Andrews, N.J.; Cameron, J.C.; Charlton, S.; Clutterbuck, E.A.; Collins, A.M.; et al. Immunogenicity, safety, and reactogenicity of heterologous COVID-19 primary vaccination incorporating mRNA, viral-vector, and protein-adjuvant vaccines in the UK (Com-COV2): A single-blind, randomised, phase 2, non-inferiority trial. Lancet 2021, 399, 36–49. [Google Scholar] [CrossRef]
- Chapin-Bardales, J.; Gee, J.; Myers, T. Reactogenicity Following Receipt of mRNA-Based COVID-19 Vaccines. JAMA 2021, 325, 2201. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The mRNA-LNP platform’s lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. iScience 2021, 24, 103479. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, X.; Yang, Y. Innate immune response to adenovirus vectors is mediated by both Toll-like receptor-dependent and -independent pathways. J. Virol. 2007, 81, 3170–3180. [Google Scholar] [CrossRef]
- Mahallawi, W.H.; Mumena, W.A. Reactogenicity and immunogenicity of the Pfizer and AstraZeneca COVID-19 vaccines. Front. Immunol. 2021, 12, 794642. [Google Scholar] [CrossRef]
- Teijaro, J.R.; Farber, D.L. COVID-19 vaccines: Modes of immune activation and future challenges. Nat. Rev. Immunol. 2021, 21, 195–197. [Google Scholar] [CrossRef]
- Sartorius, R.; Trovato, M.; Manco, R.; D’Apice, L.; De Berardinis, P. Exploiting viral sensing mediated by Toll-like receptors to design innovative vaccines, npj. Vaccines. 2021, 6, 1–15. [Google Scholar] [CrossRef]
- Mannie, M.D.; DeOca, K.B.; Bastian, A.G.; Moorman, C.D. Tolerogenic vaccines: Targeting the antigenic and cytokine niches of FOXP3+ regulatory T cells. Cell. Immunol. 2020, 355, 104173. [Google Scholar] [CrossRef]
- Krienke, C.; Kolb, L.; Diken, E.; Streuber, M.; Kirchhoff, S.; Bukur, T.; Akilli-Öztürk, Ö.; Kranz, L.M.; Berger, H.; Petschenka, J.; et al. A noninflammatory mRNA vaccine for treatment of experimental autoimmune encephalomyelitis. Science 2021, 371, 145–153. [Google Scholar] [CrossRef]
- Del Padre, M.; Todi, L.; Mitrevski, M.; Marrapodi, R.; Colantuono, S.; Fiorilli, M.; Casato, M.; Visentini, M. Reversion of anergy signatures in clonal CD21low B cells of mixed cryoglobulinemia after clearance of HCV viremia. Blood 2017, 130, 35–38. [Google Scholar] [CrossRef]
- Liu, W.; Putnam, A.L.; Xu-Yu, Z.; Szot, G.L.; Lee, M.R.; Zhu, S.; Gottlieb, P.A.; Kapranov, P.; Gingeras, T.R.; Fazekas de St Groth, B.; et al. CD127 expression inversely correlates with FoxP3 and suppressive function of human CD4+ Treg cells. J. Exp. Med. 2006, 203, 1701–1711. [Google Scholar] [CrossRef] [Green Version]
- Boyer, O.; Saadoun, D.; Abriol, J.; Dodille, M.; Piette, J.C.; Cacoub, P.; Klatzmann, D. CD4+CD25+ regulatory T-cell deficiency in patients with hepatitis C-mixed cryoglobulinemia vasculitis. Blood 2004, 103, 3428–3430. [Google Scholar] [CrossRef] [PubMed]
- Neidleman, J.; Luo, X.; McGregor, M.; Xie, G.; Murray, V.; Greene, W.C.; Lee, S.A.; Roan, N.R. mRNA vaccine-induced T cells respond identically to SARS-CoV-2 variants of concern but differ in longevity and homing properties depending on prior infection status. eLife 2021, 10, e72619. [Google Scholar] [CrossRef] [PubMed]
- Gil-Manso, S.; Carbonell, D.; López-Fernández, L.; Miguens, I.; Alonso, R.; Buño, I.; Muñoz, P.; Ochando, J.; Pion, M.; Correa-Rocha, R. Induction of high levels of specific humoral and cellular responses to SARS-CoV-2 after the administration of COVID-19 mRNA vaccines requires several days. Front. Immunol. 2021, 12, 726960. [Google Scholar] [CrossRef] [PubMed]
- de Wolf, A.C.M.T.; van Aalst, S.; Ludwig, I.S.; Bodinham, C.L.; Lewis, D.J.; van der Zee, R.; van Eden, W.; Broere, F. Regulatory T cell frequencies and phenotypes following anti-viral vaccination. PLoS ONE 2017, 12, e0179942. [Google Scholar] [CrossRef]
- Salemi, S.; Picchianti-Diamanti, A.; Germano, V.; Donatelli, I.; Di Martino, A.; Facchini, M.; Nisini, R.; Biselli, R.; Ferlito, C.; Podestà, E.; et al. Influenza vaccine administration in rheumatoid arthritis patients under treatment with TNFalpha blockers: Safety and immunogenicity. Clin. Immunol. 2010, 134, 113–120. [Google Scholar] [CrossRef]
- Ferlito, C.; Visco, V.; Biselli, .R.R.; Cattaruzza, M.S.; Carreras, G.; Salerno, G.; Lista, F.; Capobianchi, M.R.; Castilletti, C.; Lapa, D.; et al. Safety of multiple vaccinations and durability of vaccine-induced antibodies in an Italian military cohort 5 years after immunization. Biomedicines 2021, 10, 6. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug. Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Wardell, C.M.; Levings, M.K. mRNA vaccines take on immune tolerance. Nat. Biotechnol. 2021, 39, 419–421. [Google Scholar] [CrossRef]
- Kobiyama, K.; Ishii, K.J. Making innate sense of mRNA vaccine adjuvanticity. Nat. Immunol. 2022, 23, 474–476. [Google Scholar] [CrossRef]
- Suzuki, Y.; Ishihara, H. Difference in the lipid nanoparticle technology employed in three approved siRNA (Patisiran) and mRNA (COVID-19 vaccine) drugs. Drug. Metab. Pharmacokinet. 2021, 41, 100424. [Google Scholar] [CrossRef]
- Kranz, L.M.; Diken, M.; Haas, H.; Kreiter, S.; Loquai, C.; Reuter, K.C.; Meng, M.; Fritz, D.; Vascotto, F.; Hefesha, H.; et al. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 2016, 534, 396–401. [Google Scholar] [CrossRef]
- Alameh, M.G.; Tombácz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; Gaudette, B.T.; Soliman, O.Y.; Pine, M.; Hicks, P.; et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity 2021, 54, 2877–2892.e7. [Google Scholar] [CrossRef]
- Li, C.; Lee, A.; Grigoryan, L.; Arunachalam, P.S.; Scott, M.K.D.; Trisal, M.; Wimmers, F.; Sanyal, M.; Weidenbacher, P.A.; Feng, Y.; et al. Mechanisms of innate and adaptive immunity to the Pfizer-BioNTech BNT162b2 vaccine. Nat. Immunol. 2022, 23, 543–555. [Google Scholar] [CrossRef]
- Tahtinen, S.; Tong, A.J.; Himmels, P.; Oh, J.; Paler-Martinez, A.; Kim, L.; Wichner, S.; Oei, Y.; McCarron, M.J.; Freund, E.C.; et al. IL-1 and IL-1ra are key regulators of the inflammatory response to RNA vaccines. Nat. Immunol. 2022, 23, 532–542. [Google Scholar] [CrossRef]
- Parry, H.; Bruton, R.; Tut, G.; Ali, M.; Stephens, C.; Greenwood, D.; Faustini, S.; Hughes, S.; Huissoon, A.; Meade, R.; et al. Immunogenicity of single vaccination with BNT162b2 or ChAdOx1 nCoV-19 at 5-6 weeks post vaccine in participants aged 80 years or older: An exploratory analysis. Lancet. Healthy Longev. 2021, 2, e554–e560. [Google Scholar] [CrossRef]
- Collier, A.Y.; Yu, J.; McMahan, K.; Liu, J.; Chandrashekar, A.; Maron, J.S.; Atyeo, C.; Martinez, D.R.; Ansel, J.L.; Aguayo, R.; et al. Differential kinetics of immune responses elicited by COVID-19 vaccines. N. Engl. J. Med. 2021, 385, 2010–2012. [Google Scholar] [CrossRef]
- Pozzetto, B.; Legros, V.; Djebali, S.; Barateau, V.; Guibert, N.; Villard, M.; Peyrot, L.; Allatif, O.; Fassier, J.B.; Massardier-Pilonchéry, A. Immunogenicity and efficacy of heterologous ChAdOx1-BNT162b2 vaccination. Nature 2021, 600, 701–706. [Google Scholar] [CrossRef]
- Heine, A.; Juranek, S.; Brossart, P. Clinical and immunological effects of mRNA vaccines in malignant diseases. Mol. Cancer. 2021, 20, 1–20. [Google Scholar] [CrossRef]
- Pen, J.J.; De Keersmaecker, B.; Maenhout, S.K.; Van Nuffel, A.M.; Heirman, C.; Corthals, J.; Escors, D.; Bonehill, A.; Thielemans, K.; Breckpot, K.; et al. Modulation of regulatory T cell function by monocyte-derived dendritic cells matured through electroporation with mRNA encoding CD40 ligand, constitutively active TLR4, and CD70. J. Immunol. 2013, 191, 1976–1983. [Google Scholar] [CrossRef]
- Van Lint, S.; Wilgenhof, S.; Heirman, C.; Corthals, J.; Breckpot, K.; Bonehill, A.; Neyns, B.; Thielemans, K. Optimized dendritic cell-based immunotherapy for melanoma: The TriMix-formula. Cancer Immunol. Immunother. 2014, 63, 959–967. [Google Scholar] [CrossRef]
- Wilgenhof, S.; Corthals, J.; Heirman, C.; van Baren, N.; Lucas, S.; Kvistborg, P.; Thielemans, K.; Neyns, B. Phase II study of autologous monocyte-derived mRNA electroporated dendritic cells (TriMixDC-MEL) plus ipilimumab in patients with pretreated advanced melanoma. J. Clin. Oncol. 2016, 34, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Santegoets, S.J.; Dijkgraaf, E.M.; Battaglia, A.; Beckhove, P.; Britten, C.M.; Gallimore, A.; Godkin, A.; Gouttefangeas, C.; de Gruijl, T.D.; Koenen, H.J.; et al. Monitoring regulatory T cells in clinical samples: Consensus on an essential marker set and gating strategy for regulatory T cell analysis by flow cytometry. Cancer Immunol. Immunother. 2015, 64, 1271–1286. [Google Scholar] [CrossRef] [PubMed]
- Sjaastad, L.E.; Owen, D.L.; Tracy, S.I.; Farrar, M.A. Phenotypic and Functional Diversity in Regulatory T Cells. Front. Cell. Dev. Biol. 2021, 9, 715901. [Google Scholar] [CrossRef] [PubMed]
- Baecher-Allan, C.; Viglietta, V.; Hafler, D.A. Human CD4+CD25+ regulatory T cells. Semin. Immunol. 2004, 16, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Longhi, M.S.; Hussain, M.J.; Mitry, R.R.; Arora, S.K.; Mieli-Vergani, G.; Vergani, D.; Ma, Y. Functional study of CD4+CD25+ regulatory T cells in health and autoimmune hepatitis. J. Immunol. 2006, 176, 4484–4491. [Google Scholar] [CrossRef]
- Schaefer, C.; Kim, G.G.; Albers, A.; Hoermann, K.; Myers, E.N.; Whiteside, T.L. Characteristics of CD4+CD25+ regulatory T cells in the peripheral circulation of patients with head and neck cancer. Br. J. Cancer. 2005, 92, 913–920. [Google Scholar] [CrossRef]
- Landau, D.A.; Rosenzwajg, M.; Saadoun, D.; Trébeden-Negre, H.; Klatzmann, D.; Cacoub, P. Correlation of clinical and virologic responses to antiviral treatment and regulatory T cell evolution in patients with hepatitis C virus-induced mixed cryoglobulinemia vasculitis. Arthritis. Rheum. 2008, 58, 2897–2907. [Google Scholar] [CrossRef]
- Saadoun, D.; Rosenzwajg, M.; Joly, F.; Six, A.; Carrat, F.; Thibault, V.; Sene, D.; Cacoub, P.; Klatzmann, D. Regulatory T-cell responses to low-dose interleukin-2 in HCV-induced vasculitis. N. Engl. J. Med. 2011, 365, 2067–2077. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Markers | Subset | Before Vaccine | After Vaccine | p-Value |
|---|---|---|---|---|
| B cells | ||||
| CD19+ (% of lymphocytes) | Total B cells | 10.7 ± 17.1 | 11.6 ± 18.5 | NS |
| CD19+IgM+CD27− (% of CD19+) | Naive | 59.1 ± 27 | 61.1 ± 28.4 | NS |
| CD19+IgM+CD27+ (% of CD19+) | IgM memory | 14.8 ± 16.8 | 14.5 ± 17.6 | NS |
| CD19+IgM−CD27+ (% of CD19+) | Switched | 16.9 ± 14.4 | 17.5 ± 12.8 | NS |
| CD19+CD24highCD38high (% of CD19+) | Transitional | 2.2 ± 2 | 2.3 ± 3.1 | NS |
| CD19+CD21low (% of CD19+) | Exhausted | 14.7 ± 9.6 | 14.2 ± 12.9 | NS |
| T cells | ||||
| CD3+ (% of lymphocytes) | Total T cells | 70.6 ± 14 | 72.5 ± 10.7 | NS |
| CD4+ (% of lymphocytes) | Helper/Treg | 40.5 ± 10.4 | 41.1 ± 10.6 | NS |
| CD8+ (% of lymphocytes) | Cytotoxic | 28.3 ± 11.9 | 29.4 ± 10.3 | NS |
| CD3+CD4−CD8− (% of lymphocytes) | Double negative | 2.1 ± 2.6 | 2.3 ± 2 | NS |
| CD3+HLA-DR+ (% of CD3+) | Activated | 19.6 ± 18.3 | 21.5 ± 17.9 | NS |
| CD3+CD56+ (% of CD3+) | Natural killer T | 7 ± 8.6 | 5.5 ± 6.1 | 0.041 |
| CD4+CD25+CD127low (% of CD4+) | Tregs | 3.4 ± 1 | 4.9 ± 2.1 | 0.006 |
| NK cells | Natural killer | 11.1 ± 4.8 | 10.6 ± 5 | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Gualana, F.; Maiorca, F.; Marrapodi, R.; Villani, F.; Miglionico, M.; Santini, S.A.; Pulcinelli, F.; Gragnani, L.; Piconese, S.; Fiorilli, M.; et al. Opposite Effects of mRNA-Based and Adenovirus-Vectored SARS-CoV-2 Vaccines on Regulatory T Cells: A Pilot Study. Biomedicines 2023, 11, 511. https://doi.org/10.3390/biomedicines11020511
La Gualana F, Maiorca F, Marrapodi R, Villani F, Miglionico M, Santini SA, Pulcinelli F, Gragnani L, Piconese S, Fiorilli M, et al. Opposite Effects of mRNA-Based and Adenovirus-Vectored SARS-CoV-2 Vaccines on Regulatory T Cells: A Pilot Study. Biomedicines. 2023; 11(2):511. https://doi.org/10.3390/biomedicines11020511
Chicago/Turabian StyleLa Gualana, Francesca, Francesca Maiorca, Ramona Marrapodi, Francesca Villani, Marzia Miglionico, Stefano Angelo Santini, Fabio Pulcinelli, Laura Gragnani, Silvia Piconese, Massimo Fiorilli, and et al. 2023. "Opposite Effects of mRNA-Based and Adenovirus-Vectored SARS-CoV-2 Vaccines on Regulatory T Cells: A Pilot Study" Biomedicines 11, no. 2: 511. https://doi.org/10.3390/biomedicines11020511
APA StyleLa Gualana, F., Maiorca, F., Marrapodi, R., Villani, F., Miglionico, M., Santini, S. A., Pulcinelli, F., Gragnani, L., Piconese, S., Fiorilli, M., Basili, S., Casato, M., Stefanini, L., & Visentini, M. (2023). Opposite Effects of mRNA-Based and Adenovirus-Vectored SARS-CoV-2 Vaccines on Regulatory T Cells: A Pilot Study. Biomedicines, 11(2), 511. https://doi.org/10.3390/biomedicines11020511