An Overview of Methods for Cardiac Rhythm Detection in Zebrafish



 ,
, 
 ,
, 

Abstract
:1. Why Zebrafish Behavior Is a Good in vivo Model to Address Cardiac Physiology and Toxicology?
2. Overview of Embryo Cardiac Rhythm Detection
3. Commercial or Third-Party Software
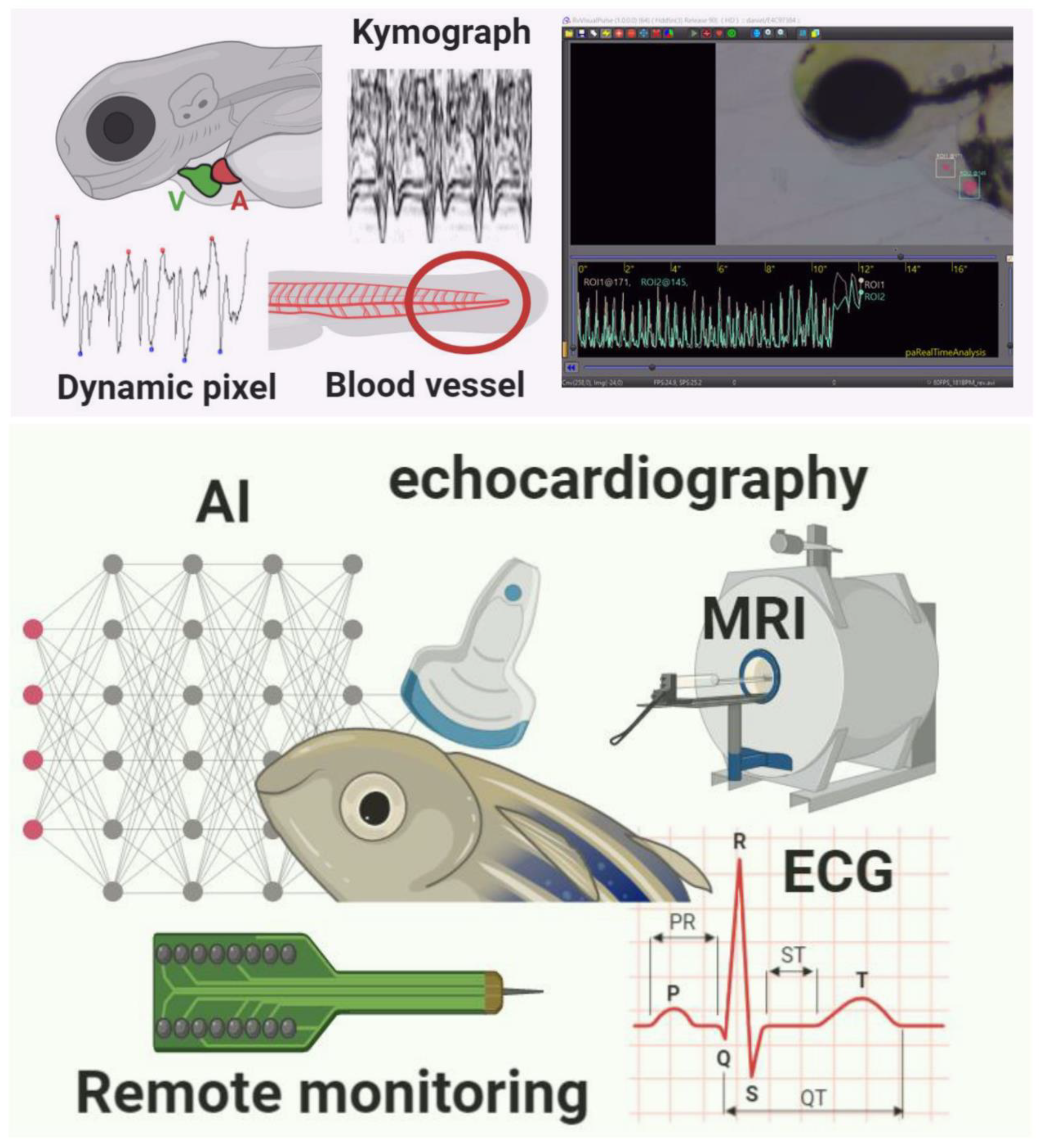
4. Dynamic Pixel Changes Method
5. Indirect Measurement from Blood Vessels
6. Kymograph-Based Method
7. Laser Confocal Scan Method
8. AI Deep Learning-Based Method
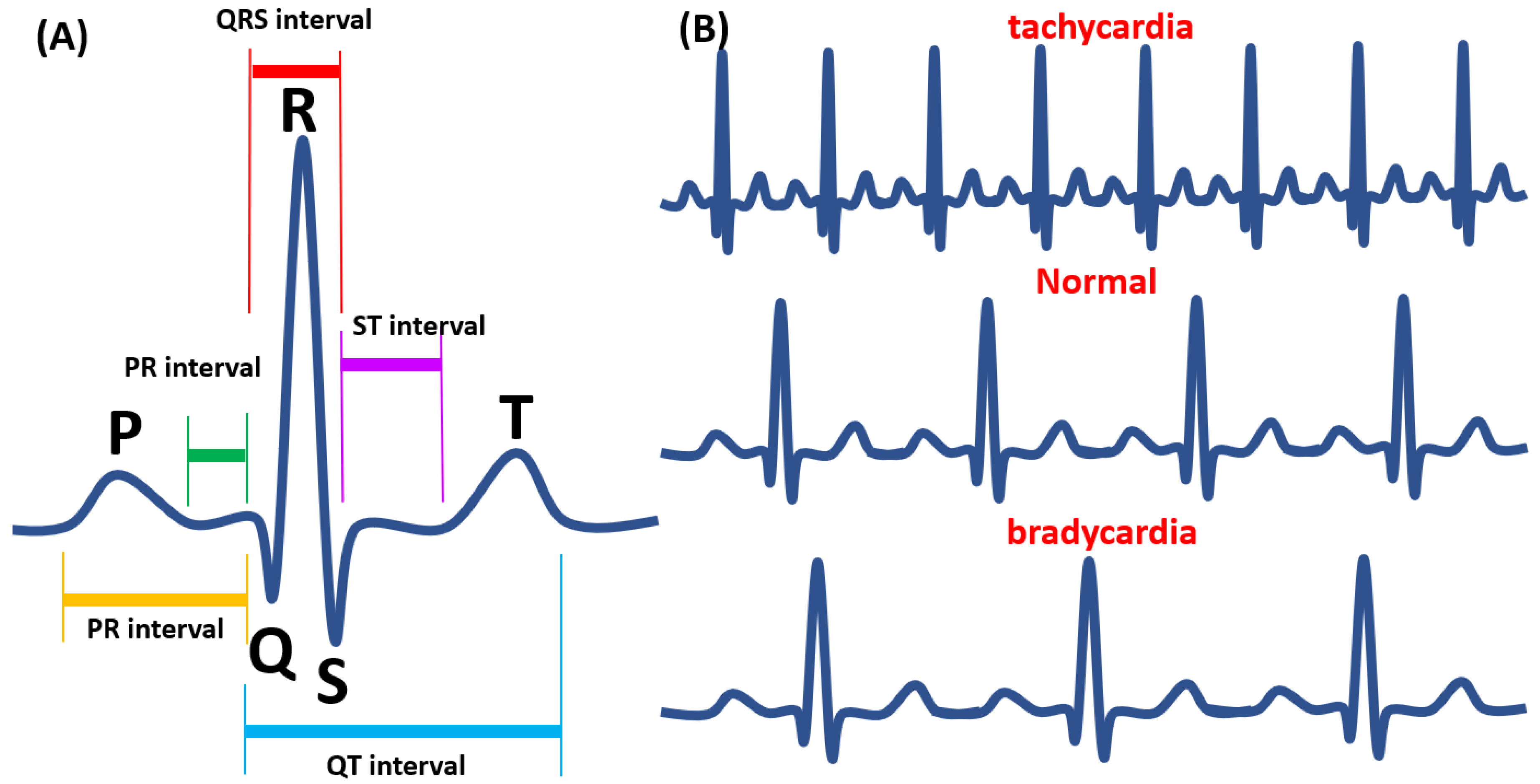
9. Electrocardiography Method (ECG)
10. Overview of Adult Cardiac Rhythm Detection
11. ECG-Based Detection Methods
11.1. Noise Interfaces for ECG
11.2. Comparison of Human ECG with Zebrafish ECG and Application for Toxicity Assessment
12. Light-Cardiogram Methods
13. Echocardiography-Based Detection Methods
14. Magnetic Resonance Imaging (MRI)
15. Remote Monitoring Methods
16. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Redfern, W.; Carlsson, L.; Davis, A.; Lynch, W.; MacKenzie, I.; Palethorpe, S.; Siegl, P.; Strang, I.; Sullivan, A.; Wallis, R. Relationships between preclinical cardiac electrophysiology, clinical QT interval prolongation and torsade de pointes for a broad range of drugs: Evidence for a provisional safety margin in drug development. Cardiovasc. Res. 2003, 58, 32–45. [Google Scholar] [CrossRef]
- Caballero, M.V.; Candiracci, M. Zebrafish as screening model for detecting toxicity and drugs efficacy. J. Unexplored Med. Data 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Giardoglou, P.; Beis, D. On zebrafish disease models and matters of the heart. Biomedicines 2019, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of zebrafish in drug discovery toxicology. Chem. Res. Toxicol. 2019, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Incardona, J.P.; Collier, T.K.; Scholz, N.L. Defects in cardiac function precede morphological abnormalities in fish embryos exposed to polycyclic aromatic hydrocarbons. Toxicol. Appl. Pharmacol. 2004, 196, 191–205. [Google Scholar] [CrossRef]
- Chen, J.-N.; Haffter, P.; Odenthal, J.; Vogelsang, E.; Brand, M.; Van Eeden, F.; Furutani-Seiki, M.; Granato, M.; Hammerschmidt, M.; Heisenberg, C.-P. Mutations affecting the cardiovascular system and other internal organs in zebrafish. Development 1996, 123, 293–302. [Google Scholar]
- Stainier, D.Y. Zebrafish genetics and vertebrate heart formation. Nat. Rev. Genet. 2001, 2, 39–48. [Google Scholar] [CrossRef]
- Sarasamma, S.; Audira, G.; Juniardi, S.; Sampurna, B.; Liang, S.-T.; Hao, E.; Lai, Y.-H.; Hsiao, C.-D. Zinc Chloride Exposure Inhibits Brain Acetylcholine Levels, Produces Neurotoxic Signatures, and Diminishes Memory and Motor Activities in Adult Zebrafish. Int. J. Mol. Sci. 2018, 19, 3195. [Google Scholar] [CrossRef] [Green Version]
- Sarmah, S.; Marrs, J.A. Zebrafish as a vertebrate model system to evaluate effects of environmental toxicants on cardiac development and function. Int. J. Mol. Sci. 2016, 17, 2123. [Google Scholar] [CrossRef] [Green Version]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef] [PubMed]
- Gut, P.; Reischauer, S.; Stainier, D.Y.; Arnaout, R. Little fish, big data: Zebrafish as a model for cardiovascular and metabolic disease. Physiol. Rev. 2017, 97, 889–938. [Google Scholar] [CrossRef] [PubMed]
- Burggren, W.W.; Gore, M. Cardiac and metabolic physiology of early larval zebrafish (Danio rerio) reflects parental swimming stamina. Front. Physiol. 2012, 3, 35. [Google Scholar]
- Fishman, M.C.; Stainier, D.Y.; Breitbart, R.E.; Westerfield, M. Zebrafish: Genetic and Embryological Methods in a Transparent Vertebrate Embryo. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 1997; Volume 52, pp. 67–82. [Google Scholar]
- Burns, C.G.; Milan, D.J.; Grande, E.J.; Rottbauer, W.; MacRae, C.A.; Fishman, M.C. High-throughput assay for small molecules that modulate zebrafish embryonic heart rate. Nat. Chem. Biol. 2005, 1, 263–264. [Google Scholar] [CrossRef]
- Staudt, D.; Stainier, D. Uncovering the molecular and cellular mechanisms of heart development using the zebrafish. Annu. Rev. Genet. 2012, 46, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Samsa, L.A.; Qian, L.; Liu, J. Advances in the study of heart development and disease using zebrafish. J. Cardiovasc. Dev. Dis. 2016, 3, 13. [Google Scholar] [CrossRef]
- De Luca, E.; Zaccaria, G.M.; Hadhoud, M.; Rizzo, G.; Ponzini, R.; Morbiducci, U.; Santoro, M.M. ZebraBeat: A flexible platform for the analysis of the cardiac rate in zebrafish embryos. Sci. Rep. 2014, 4, 4898. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.-P.; Tu, H.-C.; Fu, T.-F.; Wu, J.-M.; Chu, P.-H.; Chang, D.T.-H. An automatic method to calculate heart rate from zebrafish larval cardiac videos. BMC Bioinform. 2018, 19, 169. [Google Scholar] [CrossRef] [Green Version]
- Gaur, H.; Pullaguri, N.; Nema, S.; Purushothaman, S.; Bhargava, Y.; Bhargava, A. ZebraPace: An open-source method for cardiac-rhythm estimation in untethered zebrafish larvae. Zebrafish 2018, 15, 254–262. [Google Scholar] [CrossRef]
- Fink, M.; Callol-Massot, C.; Chu, A.; Ruiz-Lozano, P.; Belmonte, J.C.I.; Giles, W.; Bodmer, R.; Ocorr, K. A new method for detection and quantification of heartbeat parameters in Drosophila, zebrafish, and embryonic mouse hearts. Biotechniques 2009, 46, 101–113. [Google Scholar] [CrossRef]
- Ocorr, K.; Fink, M.; Cammarato, A.; Bernstein, S.I.; Bodmer, R. Semi-automated optical heartbeat analysis of small hearts. JOVE J. Vis. Exp. 2009, 31, e1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohn, J.; Liebling, M. In Vivo, High-Throughput Imaging for Functional Characterization of the Embryonic Zebrafish Heart. In Proceedings of the 2011 IEEE International Symposium on Biomedical Imaging: From Nano to Macro, Chicago, IL, USA, 30 March–2 April 2011; pp. 1549–1552. [Google Scholar]
- Gierten, J.; Pylatiuk, C.; Hammouda, O.T.; Schock, C.; Stegmaier, J.; Wittbrodt, J.; Gehrig, J.; Loosli, F. Automated high-throughput heart rate measurement in medaka and zebrafish embryos under physiological conditions. BioRxiv 2019, 548594. [Google Scholar] [CrossRef] [Green Version]
- Krishna, S.; Chatti, K.; Galigekere, R.R. Automatic and robust estimation of heart rate in zebrafish larvae. IEEE Trans. Autom. Sci. Eng. 2017, 15, 1041–1052. [Google Scholar] [CrossRef]
- Pylatiuk, C.; Sanchez, D.; Mikut, R.; Alshut, R.; Reischl, M.; Hirth, S.; Rottbauer, W.; Just, S. Automatic zebrafish heartbeat detection and analysis for zebrafish embryos. Zebrafish 2014, 11, 379–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoso, F.; Sampurna, B.P.; Lai, Y.-H.; Liang, S.-T.; Hao, E.; Chen, J.-R.; Hsiao, C.-D. Development of a Simple ImageJ-Based Method for Dynamic Blood Flow Tracking in Zebrafish Embryos and Its Application in Drug Toxicity Evaluation. Inventions 2019, 4, 65. [Google Scholar] [CrossRef] [Green Version]
- Sampurna, B.; Audira, G.; Juniardi, S.; Lai, Y.-H.; Hsiao, C.-D. A Simple ImageJ-Based Method to Measure Cardiac Rhythm in Zebrafish Embryos. Inventions 2018, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- van Opbergen, C.J.; Koopman, C.D.; Kok, B.J.; Knöpfel, T.; Renninger, S.L.; Orger, M.B.; Vos, M.A.; van Veen, T.A.; Bakkers, J.; de Boer, T.P. Optogenetic sensors in the zebrafish heart: A novel in vivo electrophysiological tool to study cardiac arrhythmogenesis. Theranostics 2018, 8, 4750. [Google Scholar] [CrossRef]
- Akerberg, A.A.; Burns, C.E.; Burns, C.G.; Nguyen, C. Deep learning enables automated volumetric assessments of cardiac function in zebrafish. Dis. Models Mech. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Xing, Q.; Huynh, V.; Parolari, T.G.; Maurer-Morelli, C.V.; Peixoto, N.; Wei, Q. Zebrafish larvae heartbeat detection from body deformation in low resolution and low frequency video. Med Biol. Eng. Comput. 2018, 56, 2353–2365. [Google Scholar] [CrossRef]
- Martin, W.K.; Tennant, A.H.; Conolly, R.B.; Prince, K.; Stevens, J.S.; DeMarini, D.M.; Martin, B.L.; Thompson, L.C.; Gilmour, M.I.; Cascio, W.E. High-Throughput Video Processing of Heart Rate Responses in Multiple Wild-type Embryonic Zebrafish per Imaging Field. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Yozzo, K.L.; Isales, G.M.; Raftery, T.D.; Volz, D.C. High-content screening assay for identification of chemicals impacting cardiovascular function in zebrafish embryos. Environ. Sci. Technol. 2013, 47, 11302–11310. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.K.; Lin, C.C.; Cheng, S.H. Noninvasive technique for measurement of heartbeat regularity in zebrafish (Danio rerio) embryos. BMC Biotechnol. 2009, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-J. Role of Endoplasmic Reticulum Calcium Pump in Alternation of Heart Failure Membrane Potential Heartbeat in Zebrafish; Department of Medicine, National Taiwan University Taiwan: Taipei, Taiwan, 2016. [Google Scholar]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Grimm, W.; Steder, U.; Menz, V.; Hoffmann, J.; Maisch, B. QT dispersion and arrhythmic events in idiopathic dilated cardiomyopathy. Am. J. Cardiol. 1996, 78, 458–461. [Google Scholar] [CrossRef]
- Nitzsche, B.; Bormuth, V.; Bräuer, C.; Howard, J.; Ionov, L.; Kerssemakers, J.; Korten, T.; Leduc, C.; Ruhnow, F.; Diez, S. Studying Kinesin Motors by Optical 3D-Nanometry in Gliding Motility Assays. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 95, pp. 247–271. [Google Scholar]
- Marra, M.H.; Tobias, Z.J.; Cohen, H.R.; Glover, G.; Weissman, T.A. In vivo time-lapse imaging in the zebrafish lateral line: A flexible, open-ended research project for an undergraduate neurobiology laboratory course. J. Undergrad. Neurosci. Educ. 2015, 13, A215. [Google Scholar] [PubMed]
- Tessadori, F.; van Weerd, J.H.; Burkhard, S.B.; Verkerk, A.O.; de Pater, E.; Boukens, B.J.; Vink, A.; Christoffels, V.M.; Bakkers, J. Identification and functional characterization of cardiac pacemaker cells in zebrafish. PLoS ONE 2012, 7, e47644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waziri, B.; Shahzad, A. Direct effects of glucose administration on heart rate, myocardial contraction, and duration of cardiac cycle in frog’s heart. J. Pract. Cardiovasc. Sci. 2018, 4, 29. [Google Scholar] [CrossRef]
- Ferreira, T.; Miura, K.; Chef, B.; Eglinger, J. Scripts: BAR 1.1.6. Available online: https://zenodo.org/record/28838#.X1CaPnkzYuU (accessed on 3 August 2020).
- Ishikawa, H.; Marshall, W.F. Efficient Live Fluorescence Imaging of Intraflagellar Transport in Mammalian Primary Cilia. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 127, pp. 189–201. [Google Scholar]
- Ott, C.; Lippincott-Schwartz, J. Visualization of live primary cilia dynamics using fluorescence microscopy. Curr. Protoc. Cell Biol. 2012, 57, 4–26. [Google Scholar] [CrossRef] [Green Version]
- Malone, M.H.; Sciaky, N.; Stalheim, L.; Hahn, K.M.; Linney, E.; Johnson, G.L. Laser-scanning velocimetry: A confocal microscopy method for quantitative measurement of cardiovascular performance in zebrafish embryos and larvae. BMC Biotechnol. 2007, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Rogers, M.A.; Aikawa, E. Cardiovascular calcification: Artificial intelligence and big data accelerate mechanistic discovery. Nat. Rev. Cardiol. 2019, 16, 261–274. [Google Scholar] [CrossRef]
- Teixidó, E.; Kießling, T.R.; Krupp, E.; Quevedo, C.; Muriana, A.; Scholz, S. Automated morphological feature assessment for zebrafish embryo developmental toxicity screens. Toxicol. Sci. 2019, 167, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, S.S.; Dóró, É.; Magyary, I.; Egginton, S.; Sík, A.; Müller, F. Optimisation of embryonic and larval ECG measurement in zebrafish for quantifying the effect of QT prolonging drugs. PLoS ONE 2013, 8, e60552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendon-Morales, E.; Prance, R.; Prance, H.; Aviles-Espinosa, R. Non-invasive electrocardiogram detection of in vivo zebrafish embryos using electric potential sensors. Appl. Phys. Lett. 2015, 107, 193701. [Google Scholar] [CrossRef]
- Cooke, S.; Chandroo, K.; Beddow, T.; Moccia, R.; McKinley, R. Swimming activity and energetic expenditure of captive rainbow trout Oncorhynchus mykiss (Walbaum) estimated by electromyogram telemetry. Aquac. Res. 2000, 31, 495–505. [Google Scholar] [CrossRef]
- Lenning, M.; Fortunato, J.; Le, T.; Clark, I.; Sherpa, A.; Yi, S.; Hofsteen, P.; Thamilarasu, G.; Yang, J.; Xu, X. Real-time monitoring and analysis of zebrafish electrocardiogram with anomaly detection. Sensors 2018, 18, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, L.; Genge, C.E.; Cua, M.; Sheng, X.; Rayani, K.; Beg, M.F.; Sarunic, M.V.; Tibbits, G.F. Functional assessment of cardiac responses of adult zebrafish (Danio rerio) to acute and chronic temperature change using high-resolution echocardiography. PLoS ONE 2016, 11, e0145163. [Google Scholar] [CrossRef] [Green Version]
- Milan, D.J.; Jones, I.L.; Ellinor, P.T.; MacRae, C.A. In vivo recording of adult zebrafish electrocardiogram and assessment of drug-induced QT prolongation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H269–H273. [Google Scholar] [CrossRef]
- Liu, C.C.; Li, L.; Lam, Y.W.; Siu, C.W.; Cheng, S.H. Improvement of surface ECG recording in adult zebrafish reveals that the value of this model exceeds our expectation. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Ho-Chiang, C.; Huang, H.; Huang, C.-C. High-frequency ultrasound deformation imaging for adult zebrafish during heart regeneration. Quant. Imaging Med. Surg. 2020, 10, 66. [Google Scholar] [CrossRef]
- Koth, J.; Maguire, M.L.; McClymont, D.; Diffley, L.; Thornton, V.L.; Beech, J.; Patient, R.K.; Riley, P.R.; Schneider, J.E. High-resolution magnetic resonance imaging of the regenerating adult zebrafish heart. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kabli, S.; Alia, A.; Spaink, H.P.; Verbeek, F.J.; De Groot, H.J. Magnetic resonance microscopy of the adult zebrafish. Zebrafish 2006, 3, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Norling, T.A. Remotely Monitor Heart-Rate and Feeding Behaviour of Fish by Using Electronic Sensor-Tags. Available online: https://stud.epsilon.slu.se/10662/1/arvennorling_t_170831.pdf (accessed on 3 August 2020).
- Brijs, J.; Sandblom, E.; Axelsson, M.; Sundell, K.; Sundh, H.; Kiessling, A.; Berg, C.; Gräns, A. Remote physiological monitoring provides unique insights on the cardiovascular performance and stress responses of freely swimming rainbow trout in aquaculture. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ho, D.; Zhao, X.; Gao, S.; Hong, C.; Vatner, D.E.; Vatner, S.F. Heart rate and electrocardiography monitoring in mice. Curr. Protoc. Mouse Biol. 2011, 1, 123–139. [Google Scholar]
- Mousavi, S.E.; Patil, J.G. Light-cardiogram, a simple technique for heart rate determination in adult zebrafish, Danio rerio. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 246, 110705. [Google Scholar] [CrossRef] [PubMed]
- da Silva, V.V.; Napoleão, P.; Geraldes, V.; Rocha, I. Non-invasive ECG recording for zebrafish. In Proceedings of the 2017 IEEE 5th Portuguese Meeting on Bioengineering (ENBENG), Coimbra, Portugal, 16–18 February 2017; pp. 1–4. [Google Scholar]
- Chaudhari, G.H.; Chennubhotla, K.S.; Chatti, K.; Kulkarni, P. Optimization of the adult zebrafish ECG method for assessment of drug-induced QTc prolongation. J. Pharmacol. Toxicol. Methods 2013, 67, 115–120. [Google Scholar] [CrossRef]
- Yeo, S.; Yoon, C.; Lien, C.-L.; Song, T.-K.; Shung, K.K. Monitoring of Adult Zebrafish Heart Regeneration Using High-Frequency Ultrasound Spectral Doppler and Nakagami Imaging. Sensors 2019, 19, 4094. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.W.; Huttner, I.G.; Santiago, C.F.; Kesteven, S.H.; Yu, Z.-Y.; Feneley, M.P.; Fatkin, D. Standardized echocardiographic assessment of cardiac function in normal adult zebrafish and heart disease models. Dis. Models Mech. 2017, 10, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Leong, I.U.S.; Skinner, J.; Shelling, A.; Love, D. Zebrafish as a model for long QT syndrome: The evidence and the means of manipulating zebrafish gene expression. Acta Physiol. 2010, 199, 257–276. [Google Scholar] [CrossRef]
- Aehlert, B.J. ECGs Made Easy-E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Haverkamp, W.; Breithardt, G.; Camm, A.J.; Janse, M.J.; Rosen, M.R.; Antzelevitch, C.; Escande, D.; Franz, M.; Malik, M.; Moss, A. The potential for QT prolongation and pro-arrhythmia by non-anti-arrhythmic drugs: Clinical and regulatory implications: Report on a Policy Conference of the European Society of Cardiology. Cardiovasc. Res. 2000, 47, 219–233. [Google Scholar] [CrossRef]
- Leor-Librach, R.J.; Bobrovsky, B.-Z.; Eliash, S.; Kaplinsky, E. Computer-controlled heart rate increase by isoproterenol infusion: Mathematical modeling of the system. Am. J. Physiol. Heart Circ. Physiol. 1999, 277, H1478–H1483. [Google Scholar] [CrossRef]
- Langheinrich, U.; Vacun, G.; Wagner, T. Zebrafish embryos express an orthologue of HERG and are sensitive toward a range of QT-prolonging drugs inducing severe arrhythmia. Toxicol. Appl. Pharmacol. 2003, 193, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Bournele, D.; Beis, D. Zebrafish models of cardiovascular disease. Heart Fail. Rev. 2016, 21, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Milan, D.J.; MacRae, C.A. Animal models for arrhythmias. Cardiovasc. Res. 2005, 67, 426–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biomedical, D. Introductory Guide to Identifiying ECG Irregularities. Available online: https://www.dcbiomed.com/proimages/materials/Brochures_and_related_Articles/Introductory_guide_to_ECG_E2_0-950820.pdf (accessed on 3 August 2020).
- Yu, F.; Huang, J.; Adlerz, K.; Jadvar, H.; Hamdan, M.H.; Chi, N.; Chen, J.-N.; Hsiai, T.K.J.B.m. Evolving cardiac conduction phenotypes in developing zebrafish larvae: Implications to drug sensitivity. Zebrafish 2010, 7, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Baer, H.U.; Hendrawan, S.; The, S.; Lelosutan, S.A.; Salim, G.; Lindl, T.; Mathes, S.; Graf-Hausner, U.; Weber, U.; Watson, R.; et al. The Intracorporeal Autologous Hepatocyte Matrix Implant for the Treatment of Chronic Liver Disease: A Modified Clinical Phase I Study. World J. Surg. Surg. Res. 2018, 1, 1067. [Google Scholar]
- Forouhar, A.; Hove, J.; Calvert, C.; Flores, J.; Jadvar, H.; Gharib, M. Electrocardiographic Characterization of Embryonic Zebrafish. In Proceedings of the 26th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, San Francisco, CA, USA, 1–4 September 2004; pp. 3615–3617. [Google Scholar]
- Van der Linde, H.; Van Deuren, B.; Teisman, A.; Towart, R.; Gallacher, D. The effect of changes in core body temperature on the QT interval in beagle dogs: A previously ignored phenomenon, with a method for correction. Br. J. Pharmacol. 2008, 154, 1474–1481. [Google Scholar] [CrossRef]
- Parker, T.; Libourel, P.-A.; Hetheridge, M.J.; Cumming, R.I.; Sutcliffe, T.P.; Goonesinghe, A.C.; Ball, J.S.; Owen, S.F.; Chomis, Y.; Winter, M.J. A multi-endpoint in vivo larval zebrafish (Danio rerio) model for the assessment of integrated cardiovascular function. J. Pharmacol. Toxicol. Methods 2014, 69, 30–38. [Google Scholar] [CrossRef]
- Vornanen, M.; Hassinen, M.J.C. Zebrafish heart as a model for human cardiac electrophysiology. Channels 2016, 10, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Lauridsen, H.; Gonzales, S.; Hedwig, D.; Perrin, K.L.; Williams, C.J.; Wrege, P.H.; Bertelsen, M.F.; Pedersen, M.; Butcher, J.T. Extracting physiological information in experimental biology via Eulerian video magnification. BMC Biol. 2019, 17, 103. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yun, M.; Nguyen, S.A.; Tran, M.; Nguyen, T.P. In Vivo Surface Electrocardiography for Adult Zebrafish. JOVE J. Vis. Exp. 2019, 150, e60011. [Google Scholar] [CrossRef]
- Bovik, A.C. Handbook of Image and Video Processing; Academic Press: Burlington, VT, USA, 2010. [Google Scholar]
- Relić, R.R.; Hristov, S.V.; Vučinić, M.M.; Poleksić, V.D.; Marković, Z.Z. Principles of fish welfare assessment in farm rearing conditions. J. Agric. Sci. 2010, 55, 273–282. [Google Scholar]
- Baras, E.; Lagardère, J.-P. Fish telemetry in aquaculture: Review and perspectives. Aquac. Int. 1995, 3, 77–102. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Author and Published Year | Require a Transgenic Fish Line? | Require a Special Script to Run the Software? | Major Platform to Calculate Heartbeat Regularity | Major Facility to Capture Heartbeat Images | Region of Interests (ROI) | What Kind of Message Can Be Obtained? | Automated Calculation? | References |
|---|---|---|---|---|---|---|---|---|
| ImageJ-Based Methods | ||||||||
| (Santoso, Sampurna et al., 2019) | No | No | ImageJ (dynamic pixel changes method) | Inverted microscope mounted with high-speed CCD camera | Dorsal Aorta | Atrium rhythm and heartbeat frequency | No | [27] |
| (Sampurna, Audira et al. 2018) | No | No | ImageJ (dynamic pixel changes method) | CCD mount onto dissecting microscope | Heart | Atrium and ventricle rhythm, whole heartbeat frequency | No | [28] |
| (Gaur, Pullaguri et al. 2018) | No | No | ImageJ (pixel intensity changes method) | An inverted microscope (Olympus IX73 series) equipped with a 10-MP camera (ProCAM HS-10 MP) | Heart | Whole heartbeat frequency | No | [20] |
| (van Opbergen, Koopman et al., 2018) | Yes | Not mentioned | ImageJ and MATLAB (method not mentioned) | Upright widefield microscope (Cairn research, Kent, UK) with a high-speed camera (AndorZyla 4.2 plus sCMOS) | Heart | Whole heartbeat frequency | Yes | [29] |
| Matlab-Based Methods | ||||||||
| (Gierten, Pylatiuk et al., 2019) | Yes | Yes | MATLAB (dynamic pixel changes method) | An ACQUIFER wide-field high content screening microscope equipped with a white LED array | Heart | Whole heartbeat frequency | Yes | [24] |
| (Akerberg, Burns, et al., 2019) | No | Yes | Matlab (deep learning platform) | ZEISS Lightsheet Z.1 microscope and optical microscope | Heart | Whole heartbeat frequency, Cardiac Function Parameters | Yes | [30] |
| (Xing, Huynh, et al., 2018) | No | Yes | Matlab | Electrophysiological recordings by an Opticam attached to the microscope | Heart | Whole heartbeat frequency | Yes | [31] |
| (Krishna, Chatti, et al., 2017) | No | Yes | Matlab | Zeiss Stereo Discovery V8 zoom stereo microscope with a ProRes color camera C3 (3 Megapixels) | Heart | Whole heartbeat frequency | Yes | [25] |
| (De Luca, Zaccaria et al., 2014) | Yes | Yes | MATLAB (threshold value method) | Leica TCS SP5X II confocal laser-scanning inverted microscope equipped with a tandem scanning system | Heart | Atrium and ventricle rhythm, whole heartbeat frequency | Yes | [18] |
| (Pylatiuk, Sanchez, et al., 2014) | Yes | Yes | MATLAB (threshold value method) | An inverted microscope (Leica DMIL LED) with a digital camera (Leica DFC 400) | Heart | Whole heartbeat frequency | Yes | [26] |
| (Fink, Callol-Massot et al., 2009) | No | Yes | MATLAB | Hamamatsu EM-CCD digital camera mounted on Leica DM-LFSA microscope | Heart | Atrium and ventricle rhythm, whole heartbeat frequency | Yes | [21] |
| Commercial Software-Based Methods | ||||||||
| (Martin, Tennant, et al., 2019) | No | Yes | FishRateZ software (commercial) (pixel intensity changes method) | AndorZyla 4.2 sCMOS (Andor Technologies, Belfast, NI, UK) camera mounted to a Nikon Ti microscope (Nikon Instruments, Melville, NY, USA) | Heart | Whole heartbeat frequency | Yes | [32] |
| (Yozzo, Isales, et al., 2013) | Yes | Yes | MetaXpress 4.0.0.24 software (commercial) | ImageXpress Micro (IXM) Widefield High-Content Screening | Heart | Whole heartbeat frequency | Yes | [33] |
| (Chan, Lin, et al., 2009) | No | Yes | A custom-made program which developed in C# language was used for digital motion analysis | Stereo-microscope (Olympus) equipped with a 3-color CCD camera | Caudal blood vessel | Whole heartbeat frequency | Yes | [34] |
| (Lin, S.J. 2016) | No | Yes | SoftEdge™ (IonOptix Corporation) | Light microscope (Axiovert 100 V microscope, Carl Zeiss, Jena, Germany) | Heart | Whole heartbeat frequency | Yes | [35] |
| (Ocorr, Fink, et al., 2009) | No | Yes | Semi-automatic Optical Heartbeat Analysis (SOHA) | Not mentioned | Heart | Whole heartbeat frequency, heart diameter measurements | Yes | [22] |
| Zgenebio and RasVector Technology | No | No | Rv Visual Pulse Analysis | CCD mount onto dissecting microscope | Heart | Atrium and ventricle rhythm | Yes | (https://www.zgenebio.com.tw) |
| Viewpoint Company | No | No | MicroZebraLab | Not mentioned | Heart | Whole heartbeat frequency | Yes | (http://www.viewpoint.fr/en/home) |
| Noldus Company | No | No | Danioscope Software | ZEISS SteREO Discovery.V8 microscope | Heart | Whole heartbeat frequency | Yes | (https://www.noldus.com/danioscope) |
| Author and Published Years | Require Special Transgenic Fish Lines? | Require Special Script to Run the Software? | Major Platform to Calculate Heartbeat Regularity | Major Facility to Capture Heartbeat Images | Region of Interests (ROI) | What Kind of Message Can Be Obtained? | Automated Calculation? | References |
|---|---|---|---|---|---|---|---|---|
| Electrocardiogram-Based Methods | ||||||||
| Mousavi & Patil (2020) | No | No | ImageJ | Stereoscope (MZ12.5, Leica Microsystems, Wetzlar, Germany) and camera (Dino-Eye Edge series AM7025X) | Whole heart | Heart rate, dominant frequency | No | [61] |
| Lenning, et al. (2017) | No | Yes | LabView | 4-channel MEA membranes for ECG acquisition | Whole heart | Whole heartbeat frequency | Yes | [51] |
| Vaz da Silva et al. (2017) | Yes | Yes | LabChart program | ECG signals a NeuroLog System | Whole heart | Whole heartbeat frequency | Yes | [62] |
| Liu et al. (2016) | No | Yes | Clampfit 10.0 software | ECG recording | Whole heart | Heart rate, PR, QRS, and QT intervals | Yes | [54] |
| Chaudari et al. (2013) | No | Yes | Power lab software | ECG recording | Whole heart | Heart rate, QT, PR and RR intervals | Yes | [63] |
| Echocardiography-Based Detection Methods | ||||||||
| Chiang, et al. (2020) | No | Yes | Matlab | A 70-MHz ultrasound imaging system and single-element transducer | Heart and dorsal aorta | Blood flow, tissue velocity, and cardiac deformation measurement | Yes | [55] |
| Yeo, Yoon et al. (2019) | No | Yes | Matlab | Custom-built, 64-channel high-frequency array imaging system and a high-frequency linear array transducer with 256 elements | Heart and dorsal aorta | Blood flow velocity, Heart regeneration | Yes | [64] |
| Wang, Huttneer et al. (2017) | Yes | Yes | Vevo Lab™ analysis software | Vevo2100® Imaging System and Vevo Imaging Station (VisualSonics) equipped with a high frequency transducer | Whole heart | Cardiovascular function parameters | Yes | [65] |
| Lee, Genge et al. (2016) | No | Yes | VisualSonics software | Vevo 2100 ultrasound system (VisualSonics1, Toronto, ON, Canada), with a 70 MHz ultrasound transducer | Whole heart | Heart rate, stroke volume (SV), ejection fraction (EF), fractional shorting (FS) and cardiac output (CO) | Yes | [52] |
| Magnetic Resonance Imaging (MRI) | ||||||||
| Koth, Maguire, et al. (2017) | Yes | Yes | Matlab | MR scanner | Whole heart | Heart regeneration | Yes | [56] |
| Kabli, Alia et al. (2006) | No | Yes | Para Vision 3.02pl running on a Silicon Graphics 02 workstation with the Irix 6.5.3 operating system and using Linux XWinNMR 3.2 | Magnetic Resonance Microscopy (MRM) consist of microimaging probe, MR magnet | Whole body | Developmental process in zebrafish | Yes | [57] |
| Remote Monitoring Methods | ||||||||
| Norling (2017) | No | Yes | Software AcceleRater (python-based web application) | DST micro-HRT logger | Whole heart | Heart rate, Behavior performance | Yes | [58] |
| Brijs, Sandblom et al. (2019) | No | Yes | LabChart Pro software | Custom-built Logger | Heart and coeliacomesenteric artery | Blood flow, Heart rate | Yes | [59] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santoso, F.; Farhan, A.; Castillo, A.L.; Malhotra, N.; Saputra, F.; Kurnia, K.A.; Chen, K.H.-C.; Huang, J.-C.; Chen, J.-R.; Hsiao, C.-D. An Overview of Methods for Cardiac Rhythm Detection in Zebrafish. Biomedicines 2020, 8, 329. https://doi.org/10.3390/biomedicines8090329
Santoso F, Farhan A, Castillo AL, Malhotra N, Saputra F, Kurnia KA, Chen KH-C, Huang J-C, Chen J-R, Hsiao C-D. An Overview of Methods for Cardiac Rhythm Detection in Zebrafish. Biomedicines. 2020; 8(9):329. https://doi.org/10.3390/biomedicines8090329
Chicago/Turabian StyleSantoso, Fiorency, Ali Farhan, Agnes L. Castillo, Nemi Malhotra, Ferry Saputra, Kevin Adi Kurnia, Kelvin H.-C. Chen, Jong-Chin Huang, Jung-Ren Chen, and Chung-Der Hsiao. 2020. "An Overview of Methods for Cardiac Rhythm Detection in Zebrafish" Biomedicines 8, no. 9: 329. https://doi.org/10.3390/biomedicines8090329
APA StyleSantoso, F., Farhan, A., Castillo, A. L., Malhotra, N., Saputra, F., Kurnia, K. A., Chen, K. H. -C., Huang, J. -C., Chen, J. -R., & Hsiao, C. -D. (2020). An Overview of Methods for Cardiac Rhythm Detection in Zebrafish. Biomedicines, 8(9), 329. https://doi.org/10.3390/biomedicines8090329