
Delayed Contralateral Nephrectomy Halted Post-Ischemic Renal Fibrosis Progression and Inhibited the Ischemia-Induced Fibromir Upregulation in Mice

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Studies
2.2. Renal Ischemia-Reperfusion and Nephrectomy
2.3. Organ Harvest
2.4. Plasma Urea Determination
2.5. RNA Preparation
2.6. qPCR Analysis of Gene Expression
2.7. MicroRNA Microarray of 1195 Targets
2.8. MicroRNA qPCR
2.9. MicroRNA Target Network Analysis
2.10. Western Blot
2.11. Statistical Analysis
3. Results
3.1. Effects of Delayed Contralateral Nephrectomy on Kidney Function
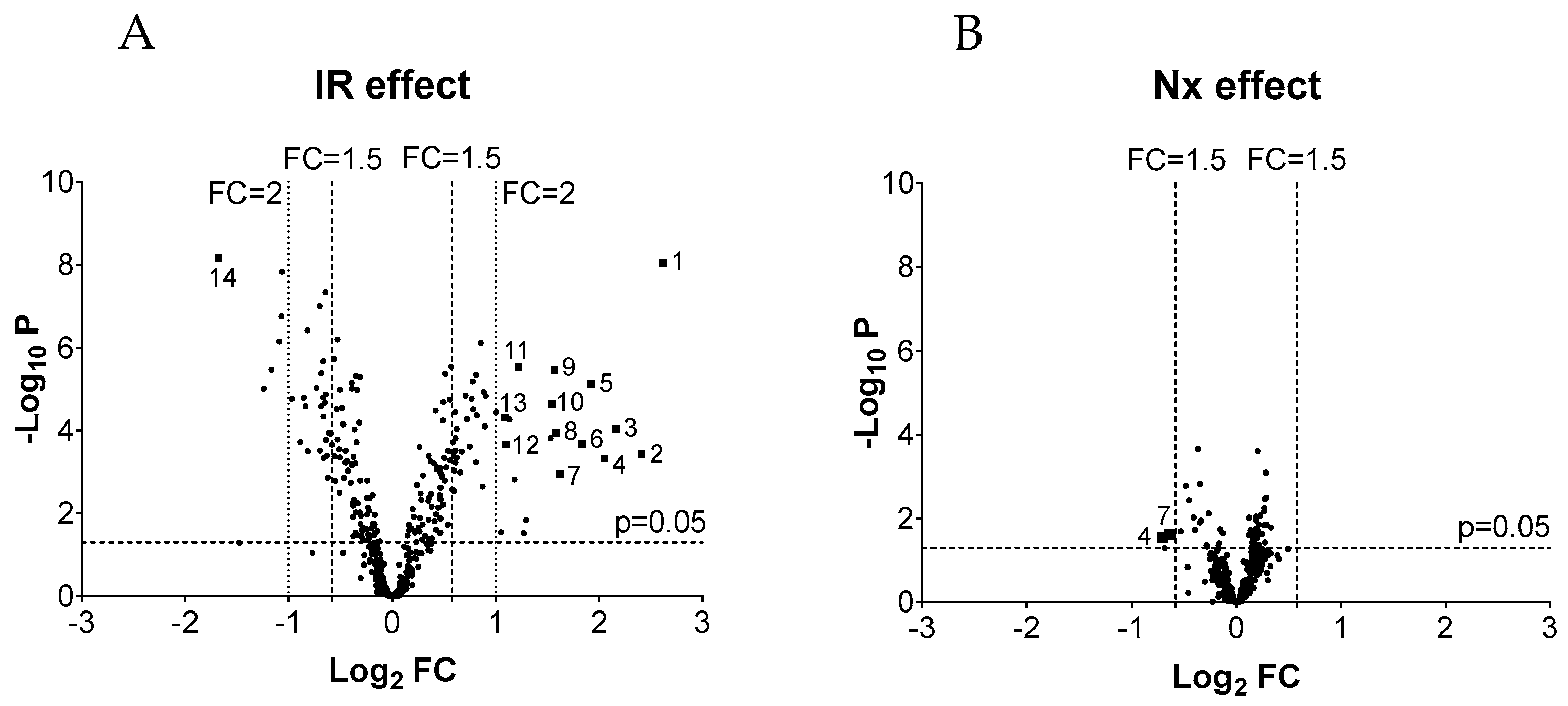
3.2. MicroRNA Validation
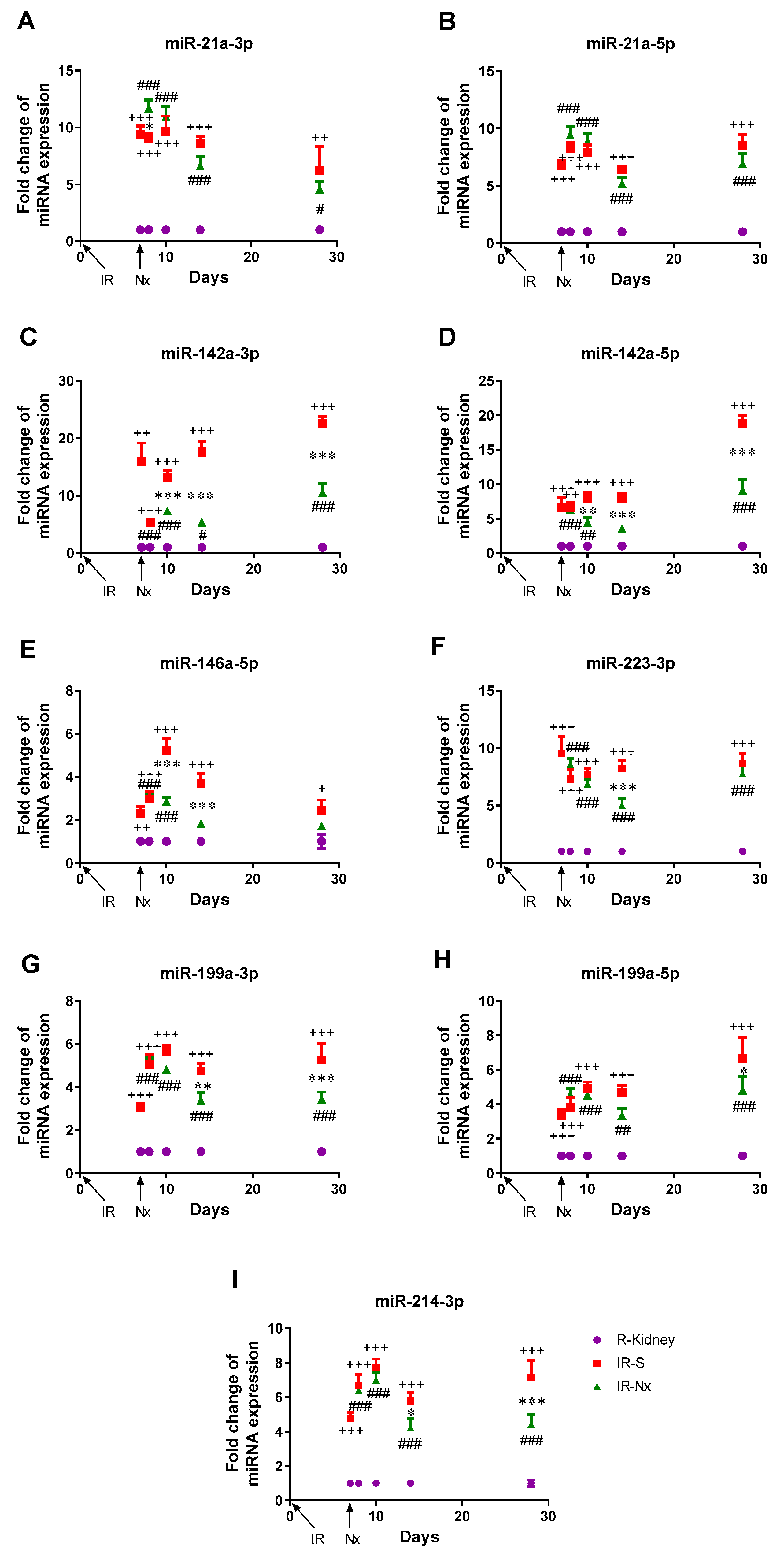
3.3. Temporal Changes of MicroRNA Expression
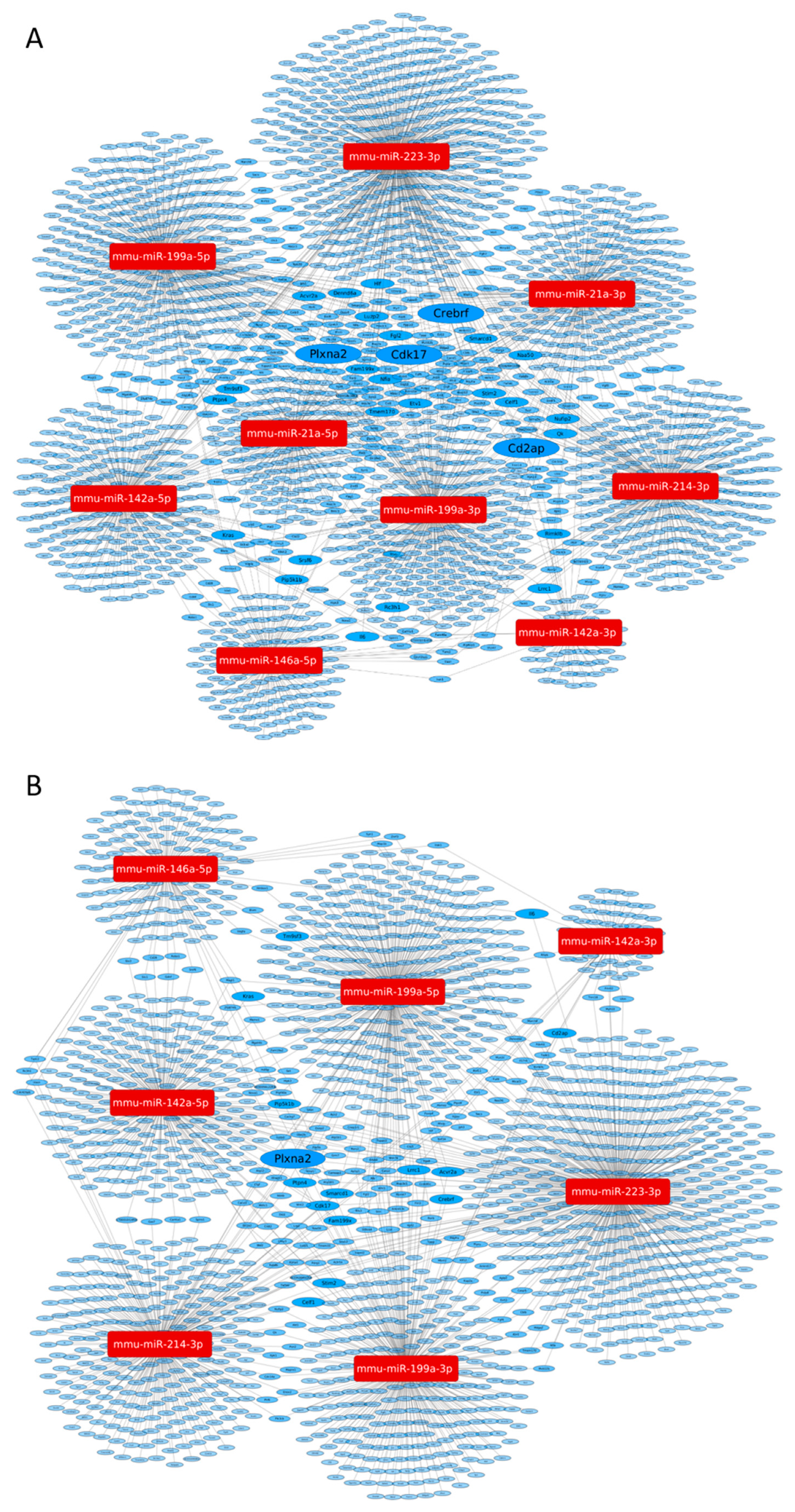
3.4. MicroRNA Target Network Analysis
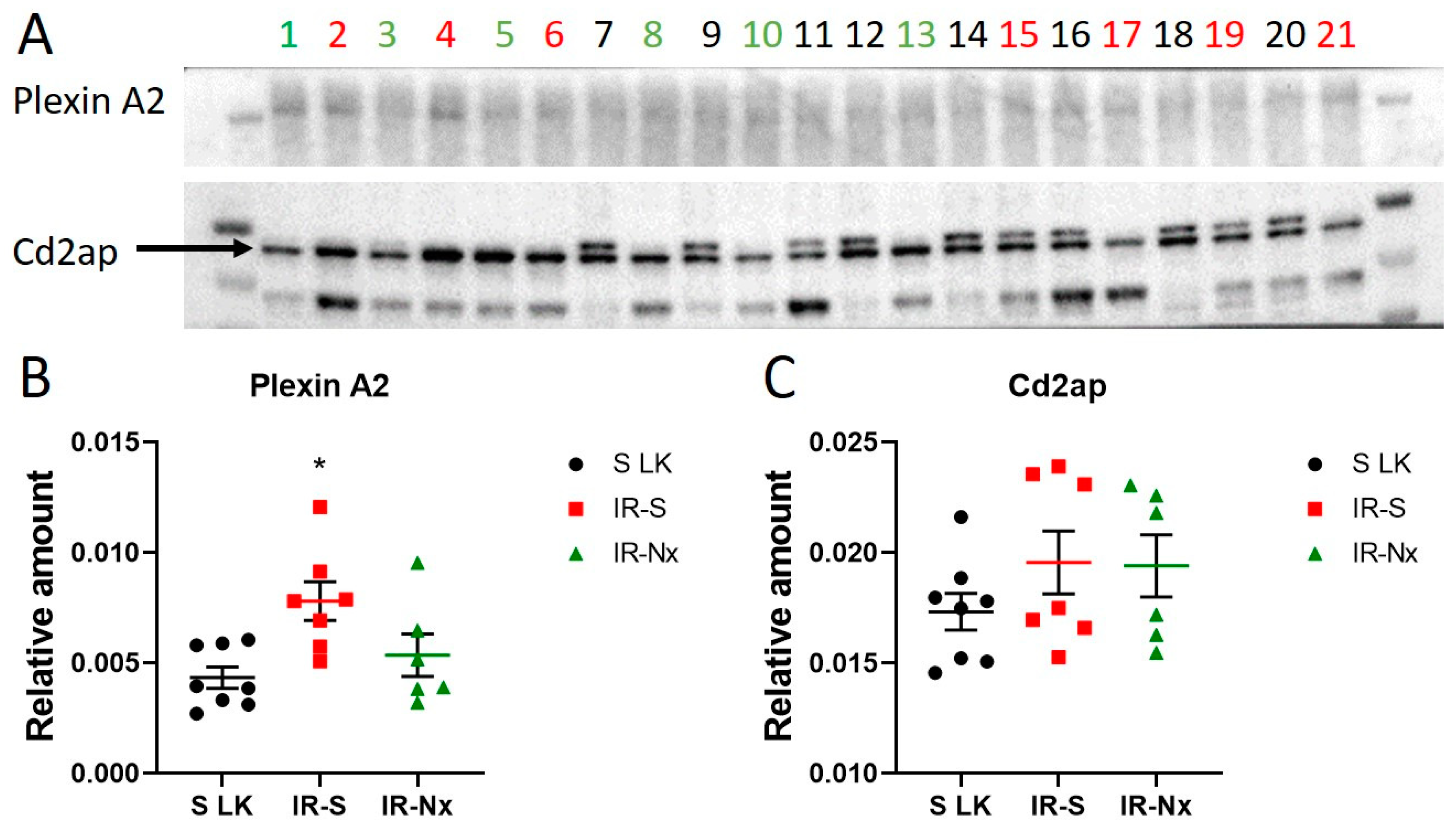
3.5. MicroRNA Target Verification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonsalez, S.R.; Cortês, A.L.; da Silva, R.C.; Lowe, J.; Prieto, M.C.; da Silva Lara, L. Acute kidney injury overview: From basic findings to new prevention and therapy strategies. Pharmacol. Ther. 2019, 200, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.C.; Tang, T.T.; Lv, L.L.; Lan, H.Y. Renal tubule injury: A driving force toward chronic kidney disease. Kidney Int. 2018, 93, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Feliers, D.; Barnes, J.L.; Oh, S.; Choudhury, G.G.; Diaz, V.; Galvan, V.; Strong, R.; Nelson, J.; Salmon, A.; et al. Hydrogen sulfide ameliorates aging-associated changes in the kidney. GeroScience 2018, 40, 163–176. [Google Scholar] [CrossRef]
- Hamar, P.; Kerjaschki, D. Blood capillary rarefaction and lymphatic capillary neoangiogenesis are key contributors to renal allograft fibrosis in an ACE inhibition rat model. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H981–H990. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, Z.; Kokeny, G.; Godo, M.; Mózes, M.; Rosivall, L.; Gross, M.-L.; Ritz, E.; Hamar, P. Increased renoprotection with ACE inhibitor plus aldosterone antagonist as compared to monotherapies--the effect on podocytes. Nephrol. Dial. Transpl. 2009, 24, 3640–3651. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Chittimalli, K.; Jahan, J.; Vasam, G.; Jarajapu, Y.P. ACE2/ACE imbalance and impaired vasoreparative functions of stem/progenitor cells in aging. GeroScience 2021, 43, 1423–1436. [Google Scholar] [CrossRef]
- Skrypnyk, N.I.; Harris, R.C.; de Caestecker, M.P. Ischemia-reperfusion model of acute kidney injury and post injury fibrosis in mice. J. Vis. Exp. 2013, 78, 6–11. [Google Scholar] [CrossRef]
- Zager, R.A.; Johnson, A.C.M.; Becker, K. Acute unilateral ischemic renal injury induces progressive renal inflammation, lipid accumulation, histone modification, and “end-stage” kidney disease. Am. J. Physiol. Physiol. 2011, 301, F1334–F1345. [Google Scholar] [CrossRef] [Green Version]
- Tod, P.; Bukosza, E.N.; Róka, B.; Kaucsár, T.; Fintha, A.; Krenács, T.; Szénási, G.; Hamar, P. Post-Ischemic Renal Fibrosis Progression Is Halted by Delayed Contralateral Nephrectomy: The Involvement of Macrophage Activation. Int. J. Mol. Sci. 2020, 21, 3825. [Google Scholar] [CrossRef]
- Kierulf-Lassen, C.; Nielsen, P.M.; Qi, H.; Damgaard, M.; Laustsen, C.; Pedersen, M.; Krag, S.; Birn, H.; Nørregaard, R.; Jespersen, B. Unilateral nephrectomy diminishes ischemic acute kidney injury through enhanced perfusion and reduced pro-inflammatory and pro-fibrotic responses. PLoS ONE 2017, 12, e0190009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Chen, Z.; Zou, Y.; Wan, X. Roles of Non-Coding RNAs in Acute Kidney Injury. Kidney Blood Press. Res. 2016, 41, 757–769. [Google Scholar] [CrossRef]
- Fan, P.-C.; Chen, C.-C.; Chen, Y.-C.; Chang, Y.-S.; Chu, P.-H. MicroRNAs in acute kidney injury. Hum. Genom. 2016, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Banaei, S. Novel role of microRNAs in renal ischemia reperfusion injury. Ren. Fail. 2015, 37, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gholaminejad, A.; Abdul Tehrani, H.; Gholami Fesharaki, M. Identification of candidate microRNA biomarkers in renal fibrosis: A meta-analysis of profiling studies. Biomarkers 2018, 23, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Chung, A.C.-K.; Lan, H.Y. MicroRNAs in renal fibrosis. Front. Physiol. 2015, 6, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, T.; Giles, C.B.; Tarantini, S.; Yabluchanskiy, A.; Balasubramanian, P.; Gautam, T.; Csipo, T.; Nyúl-Tóth, Á.; Lipecz, A.; Szabo, C.; et al. Nicotinamide mononucleotide (NMN) supplementation promotes anti-aging miRNA expression profile in the aorta of aged mice, predicting epigenetic rejuvenation and anti-atherogenic effects. GeroScience 2019, 41, 419–439. [Google Scholar] [CrossRef]
- Godwin, J.G.; Ge, X.; Stephan, K.; Jurisch, A.; Tullius, S.G.; Iacomini, J. Identification of a microRNA signature of renal ischemia reperfusion injury. Proc. Natl. Acad. Sci. USA 2010, 107, 14339–14344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Jiang, W.; Xi, X.; Zou, C.; Ye, Z. MicroRNA-21 attenuates renal ischemia reperfusion injury via targeting caspase signaling in mice. Am. J. Nephrol. 2014, 40, 215–223. [Google Scholar] [CrossRef]
- Li, Z.; Deng, X.; Kang, Z.; Wang, Y.; Xia, T.; Ding, N.; Yin, Y. Elevation of miR-21, through targeting MKK3, may be involved in ischemia pretreatment protection from ischemia–reperfusion induced kidney injury. J. Nephrol. 2016, 29, 27–36. [Google Scholar] [CrossRef]
- Kaucsár, T.; Révész, C.; Godó, M.; Krenács, T.; Albert, M.; Szalay, C.I.; Rosivall, L.; Benyó, Z.; Bátkai, S.; Thum, T.; et al. Activation of the miR-17 Family and miR-21 During Murine Kidney Ischemia-Reperfusion Injury. Nucleic Acid Ther. 2013, 23, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Song, N.; Zhang, T.; Xu, X.L.; Lu, Z.; Yu, X.; Fang, Y.; Hu, J.; Jia, P.; Teng, J.; Ding, X. miR-21 protects against ischemia/reperfusion-induced acute kidney injury by preventing epithelial cell apoptosis and inhibiting dendritic cell maturation. Front. Physiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Kriegel, A.J.; Liu, Y.; Usa, K.; Mladinov, D.; Liu, H.; Fang, Y.; Ding, X.; Liang, M. Delayed ischemic preconditioning contributes to renal protection by upregulation of miR-21. Kidney Int. 2012, 82, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Saikumar, J.; Hoffmann, D.; Kim, T.-M.; Gonzalez, V.R.; Zhang, Q.; Goering, P.L.; Brown, R.P.; Bijol, V.; Park, P.J.; Waikar, S.S.; et al. Expression, circulation, and excretion profile of microRNA-21, -155, and -18a following acute kidney injury. Toxicol. Sci. 2012, 129, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, K.L.; Gerlach, C.V.; Craciun, F.L.; Ramachandran, K.; Bijol, V.; Kissick, H.T.; Vaidya, V.S. Application of small RNA sequencing to identify microRNAs in acute kidney injury and fibrosis. Toxicol. Appl. Pharmacol. 2016, 312, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hong, Q.; Wang, Z.; Yu, Y.; Zou, X.; Xu, L. MiR-21 inhibits autophagy by targeting Rab11a in renal ischemia/reperfusion. Exp. Cell Res. 2015, 338, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Song, N.; Zhang, X.; Jiao, X.; Hu, J.; Liang, M.; Teng, J.; Ding, X. Renal Protection Mediated by Hypoxia Inducible Factor-1α Depends on Proangiogenesis Function of miR-21 by Targeting Thrombospondin 1. Transplantation 2017, 101, 1811–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarjou, A.; Yang, S.; Abraham, E.; Agarwal, A.; Liu, G. Identification of a microRNA signature in renal fibrosis: Role of miR-21. Am. J. Physiol. Renal Physiol. 2011, 301, F793–F801. [Google Scholar] [CrossRef] [Green Version]
- Chau, B.N.; Xin, C.; Hartner, J.; Ren, S.; Castano, A.P.; Linn, G.; Li, J.; Tran, P.T.; Kaimal, V.; Huang, X.; et al. MicroRNA-21 promotes fibrosis of the kidney by silencing metabolic pathways. Sci. Transl. Med. 2012, 4, 121ra18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.-B.; Zheng, Y.; Jin, L.-W.; Zhou, Z.-H.; Li, Z.-Y. Microvesicles containing microRNA-21 secreted by proximal tubular epithelial cells are involved in renal interstitial fibrosis by activating AKT pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 707–714. [Google Scholar] [PubMed]
- Liu, X.J.; Hong, Q.; Wang, Z.; Yu, Y.-Y.; Zou, X.; Xu, L.-H. MicroRNA21 promotes interstitial fibrosis via targeting DDAH1: A potential role in renal fibrosis. Mol. Cell. Biochem. 2016, 411, 181–189. [Google Scholar] [CrossRef]
- Liu, X.; Hong, Q.; Wang, Z.; Yu, Y.; Zou, X.; Xu, L. Transforming growth factor-β-sphingosine kinase 1/S1P signaling upregulates microRNA-21 to promote fibrosis in renal tubular epithelial cells. Exp. Biol. Med. 2016, 241, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, A.C.K.; Dong, Y.; Yang, W.; Zhong, X.; Li, R.; Lan, H.Y. Smad7 suppresses renal fibrosis via altering expression of TGF-β/Smad3-regulated microRNAs. Mol. Ther. 2013, 21, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Chung, A.C.K.; Chen, H.-Y.; Meng, X.-M.; Lan, H.Y. Smad3-mediated upregulation of miR-21 promotes renal fibrosis. J. Am. Soc. Nephrol. 2011, 22, 1668–1681. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Miao, J.; Luo, J.; Yuan, Q.; Cao, H.; Su, W.; Zhou, Y.; Jiang, L.; Fang, L.; Dai, C.; et al. The feedback loop between miR-21, PDCD4 and AP-1 functions as a driving force for renal fibrogenesis. J. Cell Sci. 2018, 131, jcs202317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denby, L.; Ramdas, V.; McBride, M.W.; Wang, J.; Robinson, H.; McClure, J.; Crawford, W.; Lu, R.; Hillyard, D.Z.; Khanin, R.; et al. miR-21 and miR-214 are consistently modulated during renal injury in rodent models. Am. J. Pathol. 2011, 179, 661–672. [Google Scholar] [CrossRef]
- Tang, C.-R.; Luo, S.-G.; Lin, X.; Wang, J.; Liu, Y. Silenced miR-21 inhibits renal interstitial fibrosis via targeting ERK1/2 signaling pathway in mice. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 110–116. [Google Scholar] [PubMed]
- Amrouche, L.; Desbuissons, G.; Rabant, M.; Sauvaget, V.; Nguyen, C.; Benon, A.; Barre, P.; Rabaté, C.; Lebreton, X.; Gallazzini, M.; et al. MicroRNA-146a in Human and Experimental Ischemic AKI: CXCL8-Dependent Mechanism of Action. J. Am. Soc. Nephrol. 2017, 28, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Morishita, Y.; Imai, T.; Yoshizawa, H.; Watanabe, M.; Ishibashi, K.; Muto, S.; Nagata, D. Delivery of microRNA-146a with polyethylenimine nanoparticles inhibits renal fibrosis in vivo. Int. J. Nanomed. 2015, 10, 3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Wang, H.; Wang, Y.; Peng, X.; Li, J.; Gu, W.; He, T.; Chen, M. Regulation and mechanism of miR-146 on renal ischemia reperfusion injury. Pharmazie 2018, 73, 29–34. [Google Scholar]
- Lv, W.; Fan, F.; Wang, Y.; Gonzalez-Fernandez, E.; Wang, C.; Yang, L.; Booz, G.W.; Roman, R.J. Therapeutic potential of microRNAs for the treatment of renal fibrosis and CKD. Physiol. Genom. 2018, 50, 20–34. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ágg, B.; Baranyai, T.; Makkos, A.; Vető, B.; Faragó, N.; Zvara, Á.; Giricz, Z.; Veres, D.V.; Csermely, P.; Arányi, T.; et al. MicroRNA interactome analysis predicts post-transcriptional regulation of ADRB2 and PPP3R1 in the hypercholesterolemic myocardium. Sci. Rep. 2018, 8, 10134. [Google Scholar] [CrossRef] [PubMed]
- Bencsik, P.; Kiss, K.; Ágg, B.; Baán, J.A.; Ágoston, G.; Varga, A.; Gömöri, K.; Mendler, L.; Faragó, N.; Zvara, Á.; et al. Sensory Neuropathy Affects Cardiac miRNA Expression Network Targeting IGF-1, SLC2a-12, EIF-4e, and ULK-2 mRNAs. Int. J. Mol. Sci. 2019, 20, 991. [Google Scholar] [CrossRef] [Green Version]
- Sághy, É.; Vörös, I.; Ágg, B.; Kiss, B.; Koncsos, G.; Varga, Z.V.; Görbe, A.; Giricz, Z.; Schulz, R.; Ferdinandy, P. Cardiac miRNA Expression and their mRNA Targets in a Rat Model of Prediabetes. Int. J. Mol. Sci. 2020, 21, 2128. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.-H.; Shrestha, S.; Yang, C.-D.; Chang, N.-W.; Lin, Y.-L.; Liao, K.-W.; Huang, W.-C.; Sun, T.-H.; Tu, S.-J.; Lee, W.-H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Ágg, B.; Császár, A.; Szalay-Bekő, M.; Veres, D.V.; Mizsei, R.; Ferdinandy, P.; Csermely, P.; Kovács, I.A. The EntOptLayout Cytoscape plug-in for the efficient visualization of major protein complexes in protein-protein interaction and signalling networks. Bioinformatics 2019, 35, 4490–4492. [Google Scholar] [CrossRef] [PubMed]
- Motulsky, H.J.; Brown, R.E. Detecting outliers when fitting data with nonlinear regression - a new method based on robust nonlinear regression and the false discovery rate. BMC Bioinform. 2006, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Jia, P.; Fang, Y.; Liu, H.; Jiao, X.; He, J.C.; Ding, X. miR-146a is essential for lipopolysaccharide (LPS)-induced cross-tolerance against kidney ischemia/reperfusion injury in mice. Sci. Rep. 2016, 6, 27091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, F.; Tsuchiya, S.; Terasawa, K.; Tsujimoto, G. Intra-platform repeatability and inter-platform comparability of microRNA microarray technology. PLoS ONE 2009, 4, e5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Git, A.; Dvinge, H.; Salmon-Divon, M.; Osborne, M.; Kutter, C.; Hadfield, J.; Bertone, P.; Caldas, C. Systematic comparison of microarray profiling, real-time PCR, and next-generation sequencing technologies for measuring differential microRNA expression. RNA 2010, 16, 991–1006. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Cui, Y.; Zhang, N.; Yao, X.; Wang, Z.; Yang, L. Oleanolic acid attenuated diabetic mesangial cell injury by activation of autophagy via miRNA-142-5p/PTEN signaling. Cytotechnology 2019, 71, 925–933. [Google Scholar] [CrossRef]
- Soltaninejad, E.; Nicknam, M.H.; Nafar, M.; Ahmadpoor, P.; Pourrezagholi, F.; Sharbafi, M.H.; Hosseinzadeh, M.; Foroughi, F.; Yekaninejad, M.S.; Bahrami, T.; et al. Differential expression of microRNAs in renal transplant patients with acute T-cell mediated rejection. Transpl. Immunol. 2015, 33, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dov, I.Z.; Muthukumar, T.; Morozov, P.; Mueller, F.B.; Tuschl, T.; Suthanthiran, M. MicroRNA Sequence Profiles of Human Kidney Allografts With or Without Tubulointerstitial Fibrosis. Transplant. J. 2012, 94, 1086–1094. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Yang, D.K.; Kim, S.; Kang, H. MIR-142-3p Is a Regulator of the TGFβ-Mediated Vascular Smooth Muscle Cell Phenotype. J. Cell. Biochem. 2015, 116, 2325–2333. [Google Scholar] [CrossRef] [PubMed]
- Ichii, O.; Otsuka, S.; Sasaki, N.; Namiki, Y.; Hashimoto, Y.; Kon, Y. Altered expression of microRNA miR-146a correlates with the development of chronic renal inflammation. Kidney Int. 2012, 81, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Pei, L.; Lin, F.; Yin, H.; Li, X.; He, W.; Liu, N.; Gou, X. Exosomes from human-bone-marrow-derived mesenchymal stem cells protect against renal ischemia/reperfusion injury via transferring miR-199a-3p. J. Cell. Physiol. 2019, 234, 23736–23749. [Google Scholar] [CrossRef]
- Delić, D.; Wiech, F.; Urquhart, R.; Gabrielyan, O.; Rieber, K.; Rolser, M.; Tsuprykov, O.; Hasan, A.A.; Krämer, B.K.; Baum, P.; et al. Linagliptin and telmisartan induced effects on renal and urinary exosomal miRNA expression in rats with 5/6 nephrectomy. Sci. Rep. 2020, 10, 3373. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhu, G.; He, W.; Yin, H.; Lin, F.; Gou, X.; Li, X. BMSCs protect against renal ischemia-reperfusion injury by secreting exosomes loaded with miR-199a-5p that target BIP to inhibit endoplasmic reticulum stress at the very early reperfusion stages. FASEB J. 2019, 33, 5440–5456. [Google Scholar] [CrossRef]
- Lino Cardenas, C.L.; Henaoui, I.S.; Courcot, E.; Roderburg, C.; Cauffiez, C.; Aubert, S.; Copin, M.-C.; Wallaert, B.; Glowacki, F.; Dewaeles, E.; et al. miR-199a-5p Is upregulated during fibrogenic response to tissue injury and mediates TGFbeta-induced lung fibroblast activation by targeting caveolin-1. PLoS Genet. 2013, 9, e1003291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, M.D.; Furr, J.W.; Munshi, Y.; Roy-O’Reilly, M.A.; Maniskas, M.E.; Koellhoffer, E.C.; D’Aigle, J.; Sansing, L.H.; McCullough, L.D.; Urayama, A. Transforming growth factor-β promotes basement membrane fibrosis, alters perivascular cerebrospinal fluid distribution, and worsens neurological recovery in the aged brain after stroke. GeroScience 2019, 41, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Lv, C.; Chen, F.; Ma, X.; Shao, Y.; Wang, Q. The function of miR-199a-5p/Klotho regulating TLR4/NF-κB p65/NGAL pathways in rat mesangial cells cultured with high glucose and the mechanism. Mol. Cell. Endocrinol. 2015, 417, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, W.; Li, H. miR-214 ameliorates acute kidney injury via targeting DKK3 and activating of Wnt/ β-catenin signaling pathway. Biol. Res. 2018, 51, 1–10. [Google Scholar] [CrossRef]
- Liu, M.; Liu, L.; Bai, M.; Zhang, L.; Ma, F.; Yang, X.; Sun, S. Hypoxia-induced activation of Twist/miR-214/E-cadherin axis promotes renal tubular epithelial cell mesenchymal transition and renal fibrosis. Biochem. Biophys. Res. Commun. 2018, 495, 2324–2330. [Google Scholar] [CrossRef]
- Denby, L.; Ramdas, V.; Lu, R.; Conway, B.R.; Grant, J.S.; Dickinson, B.; Aurora, A.B.; McClure, J.D.; Kipgen, D.; Delles, C.; et al. MicroRNA-214 antagonism protects against renal fibrosis. J. Am. Soc. Nephrol. 2014, 25, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Chen, H.; Ding, D.; Song, R.; Lin, J.; Zhang, Y.; Guo, Y.; Chen, S.; Ding, G.; Zhang, Y.; et al. MicroRNA-214 promotes chronic kidney disease by disrupting mitochondrial oxidative phosphorylation. Kidney Int. 2019, 95, 1389–1404. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, X.; Chen, C.; Zhou, J.; Han, M. Bone mesenchymal stem cells ameliorate ischemia/reperfusion-induced damage in renal epithelial cells via microRNA-223. Stem Cell Res. Ther. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Go, V.; Bowley, B.G.E.; Pessina, M.A.; Zhang, Z.G.; Chopp, M.; Finklestein, S.P.; Rosene, D.L.; Medalla, M.; Buller, B.; Moore, T.L. Extracellular vesicles from mesenchymal stem cells reduce microglial-mediated neuroinflammation after cortical injury in aged Rhesus monkeys. GeroScience 2020, 42, 1–17. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Y.; Deng, Y.; Li, H. MicroRNA-223 Regulates Cardiac Fibrosis after Myocardial Infarction by Targeting RASA1. Cell. Physiol. Biochem. 2018, 46, 1439–1454. [Google Scholar] [CrossRef]
- Meschiari, C.A.; Ero, O.K.; Pan, H.; Finkel, T.; Lindsey, M.L. The impact of aging on cardiac extracellular matrix. GeroScience 2017, 39, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Shih, N.Y.; Li, J.; Karpitskii, V.; Nguyen, A.; Dustin, M.L.; Kanagawa, O.; Miner, J.H.; Shaw, A.S. Congenital nephrotic syndrome in mice lacking CD2-associated protein. Science 1999, 286, 312–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Wu, H.; Green, G.; Winkler, C.A.; Kopp, J.B.; Miner, J.H.; Unanue, E.R.; Shaw, A.S. CD2-associated protein haploinsufficiency is linked to glomerular disease susceptibility. Science 2003, 300, 1298–1300. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Worzfeld, T. Semaphorins and Plexins in Kidney Disease. Nephron 2016, 132, 93–100. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, L.; Tao, X.; Song, Y.; Cui, J.; Wan, J. The role of podocyte damage in the etiology of ischemia-reperfusion acute kidney injury and post-injury fibrosis. BMC Nephrol. 2019, 20, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, F.; Villegas, G.; Teichman, J.; Mundel, P.; Tufro, A. Autocrine class 3 semaphorin system regulates slit diaphragm proteins and podocyte survival. Kidney Int. 2006, 69, 1564–1569. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, P.; Jayakumar, C.; Mohamed, R.; Weintraub, N.L.; Ramesh, G. Semaphorin 3A inactivation suppresses ischemia-reperfusion-induced inflammation and acute kidney injury. Am. J. Physiol. Renal Physiol. 2014, 307, F183–F194. [Google Scholar] [CrossRef]
- Park, M.; Kwon, C.H.; Ha, H.K.; Han, M.; Song, S.H. RNA-Seq identifies condition-specific biological signatures of ischemia-reperfusion injury in the human kidney. BMC Nephrol. 2020, 21, 398. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Animals | ||||
|---|---|---|---|---|
| Day of Termination | IR-S Group | IR-Nx Group | S-S Group | S-Nx Group |
| Day 7 | 8 | - | - | - |
| Day 8 | 9 | 10 | 6 | 6 |
| Day 10 | 9 | 9 | 8 | 7 |
| Day 14 | 8 | 7 | - | - |
| Day 28 | 9 | 8 | - | - |
| Target | Forward Primer | Reverse Primer |
|---|---|---|
| Lcn-2 | AGGTGGTACGTTGTGGGC | CTGTACCTGAGGATACCTGTG |
| Tnf-α | AAATGGCCTCCCTCTCATCA | AGATAGCAAATCGGCTGACG |
| Tgf-β | CAACAATTCCTGGCGTTACCTTGG | GAAAGCCCTGTATTCCGTCTCCTT |
| 18S | CCAGAATGAGGATCCCAGAA | ACCACCTGAAACATGCAACA |
| miRNA | Fold Change (S-S vs. IR-S, Microarray) | p Value ANOVA (Microarray) | Fold Change (S-S vs. IR-S, qPCR) | p Value ANOVA (qPCR) | Result of Validation | |
|---|---|---|---|---|---|---|
| miRNAs upregulated by IR | ||||||
| 1 | mmu-miR-21a-5p | 6.13 ± 0.63 | 0.0000 | 8.09 ± 1.35 | 0.000 | verified |
| 2 | mmu-miR-2137 | 5.31 ± 1.88 | 0.0004 | 4.27 ± 2.51 | 0.011 | verified |
| 3 | mmu-miR-142-3p | 4.48 ± 1.31 | 0.0001 | 7.38 ± 2.10 | 0.000 | verified |
| 4 | mmu-miR-762 | 4.15 ± 1.46 | 0.0005 | 0.96 ± 0.36 | 0.900 | failed |
| 5 | mmu-miR-223-3p | 3.79 ± 0.79 | 0.0000 | 5.42 ± 1.65 | 0.000 | verified |
| 6 | mmu-miR-142-5p | 3.58 ± 0.81 | 0.0002 | 9.63 ± 3.08 | 0.000 | verified |
| 7 | mmu-miR-2861 | 3.08 ± 0.94 | 0.0011 | - | - | failed |
| 8 | mmu-miR-3102-5p | 2.99 ± 0.77 | 0.0001 | 0.64 ± 0.23 | 0.136 | failed |
| 9 | mmu-miR-199a-5p | 2.97 ± 0.39 | 0.0000 | 3.45 ± 1.34 | 0.000 | verified |
| 10 | mmu-miR-199a-3p/ mmu-miR-199b-3p | 2.92 ± 0.53 | 0.0000 | 4.03 ± 1.05 | 0.000 | verified |
| 11 | mmu-miR-214-3p | 2.33 ± 0.16 | 0.0000 | 4.19 ± 1.01 | 0.000 | verified |
| 12 | mmu-miR-146a-5p | 2.15 ± 0.45 | 0.0002 | 2.46 ± 0.75 | 0.000 | verified |
| 13 | mmu-miR-21a-3p | 2.13 ± 0.36 | 0.0000 | 12.91 ± 1.98 | 0.000 | partially verified |
| miRNAs downregulated by IR | ||||||
| 1 | mmu-miR-129-1-3p | 0.31 ± 0.06 | 0.0000 | 0.33 ± 0.19 | 0.174 | failed |
| A | Gene Symbol | NCBI Gene Description | miRNA Target Strength | Associated miRNAs | miRTarBase 8.0 | miRDB v5.0 Score | TargetScan Mouse 7.2 Context++ Score |
|---|---|---|---|---|---|---|---|
| 1 | CD2AP | CD2-associated protein | 4 | mmu-miR-21a-3p | − | 93.3 | - |
| mmu-miR-142a-3p | + | - | - | ||||
| mmu-miR-199a-3p | − | 94.5 | −0.603 | ||||
| mmu-miR-223-3p | − | - | −0.299 | ||||
| 2 | Cdk17 | cyclin-dependent kinase 17 | 4 | mmu-miR-21a-3p | − | 95.4 | −0.210 |
| mmu-miR-142a-5p | − | 93.2 | - | ||||
| mmu-miR-199a-3p | − | 88.3 | −0.342 | ||||
| mmu-miR-223-3p | − | 98.8 | −0.834 | ||||
| 3 | Crebrf | CREB3 regulatory factor | 4 | mmu-miR-21a-3p | − | 90.4 | - |
| mmu-miR-199a-3p | − | - | −0.220 | ||||
| mmu-miR-199a-5p | − | - | −0.213 | ||||
| mmu-miR-223-3p | − | 80.4 | - | ||||
| 4 | PlxnA2 | plexin A2 | 4 | mmu-miR-142a-5p | − | 91.1 | - |
| mmu-miR-199a-5p | − | 95.5 | −0.369 | ||||
| mmu-miR-214-3p | − | 82.4 | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 5 | Acvr2a | activin receptor IIA | 3 | mmu-miR-199a-3p | − | 98.6 | −0.614 |
| mmu-miR-199a-5p | − | - | −0.306 | ||||
| mmu-miR-223-3p | − | 94.2 | −0.436 | ||||
| 6 | Celf1 | CUGBP, Elav-like family member 1 | 3 | mmu-miR-199a-3p | − | - | −0.253 |
| mmu-miR-214-3p | − | 81.0 | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 7 | Dennd6a | DENN/MADD domain containing 6A | 3 | mmu-miR-21a-3p | − | 94.6 | - |
| mmu-miR-199a-5p | − | - | −0.236 | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 8 | Etv1 | ets variant 1 | 3 | mmu-miR-21a-3p | − | 99.5 | - |
| mmu-miR-21a-5p | − | - | −0.238 | ||||
| mmu-miR-142a-5p | − | 92.8 | - | ||||
| 9 | Fam199x | family with sequence similarity 199, X-linked | 3 | mmu-miR-142a-5p | − | 84.4 | - |
| mmu-miR-199a-3p | − | 96.0 | −0.485 | ||||
| mmu-miR-223-3p | − | 93.5 | - | ||||
| 10 | Fgl2 | fibrinogen-like protein 2 | 3 | mmu-miR-21a-3p | − | 93.8 | - |
| mmu-miR-199a-3p | − | - | −0.208 | ||||
| mmu-miR-199a-5p | + | - | - | ||||
| 11 | Hlf | hepatic leukemia factor | 3 | mmu-miR-21a-3p | − | 99.1 | - |
| mmu-miR-199a-5p | − | - | −0.299 | ||||
| mmu-miR-223-3p | + | - | −0.584 | ||||
| 12 | Il6 | interleukin 6 | 3 | mmu-miR-142a-3p | + | - | - |
| mmu-miR-146a-5p | + | - | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 13 | Kras | Kirsten rat sarcoma viral oncogene homolog | 3 | mmu-miR-142a-5p | − | 92.5 | - |
| mmu-miR-146a-5p | − | 88.6 | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 14 | Lrrc1 | leucine rich repeat containing 1 | 3 | mmu-miR-142a-3p | − | 92.9 | - |
| mmu-miR-199a-3p | − | - | −0.272 | ||||
| mmu-miR-214-3p | − | 81.7 | - | ||||
| 15 | Luzp2 | leucine zipper protein 2 | 3 | mmu-miR-21a-3p | − | 97.1 | - |
| mmu-miR-142a-5p | − | - | −0.205 | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 16 | Naa50 | N(alpha)-acetyltransferase 50, NatE catalytic subunit | 3 | mmu-miR-21a-3p | − | 99.7 | - |
| mmu-miR-214-3p | − | 94.6 | - | ||||
| mmu-miR-223-3p | − | 86.6 | −0.284 | ||||
| 17 | Nfia | nuclear factor I/A | 3 | mmu-miR-21a-5p | − | - | −0.321 |
| mmu-miR-199a-3p | − | - | −0.268 | ||||
| mmu-miR-223-3p | − | - | −0.296 | ||||
| 18 | Nufip2 | nuclear fragile X mental retardation protein interacting protein 2 | 3 | mmu-miR-21a-3p | − | 83.0 | - |
| mmu-miR-199a-3p | − | 88.6 | −0.330 | ||||
| mmu-miR-214-3p | − | 98.7 | - | ||||
| 19 | Pip5k1b | phosphatidylinositol-4-phosphate 5-kinase, type 1 beta | 3 | mmu-miR-142a-5p | − | - | −0.202 |
| mmu-miR-146a-5p | − | - | −0.236 | ||||
| mmu-miR-199a-3p | − | 88.7 | −0.319 | ||||
| 20 | Ptpn4 | protein tyrosine phosphatase, non-receptor type 4 | 3 | mmu-miR-142a-5p | − | 97.9 | - |
| mmu-miR-199a-3p | − | - | −0.274 | ||||
| mmu-miR-199a-5p | − | - | −0.253 | ||||
| 21 | Qk | quaking | 3 | mmu-miR-21a-3p | − | 84.5 | - |
| mmu-miR-199a-3p | + | - | - | ||||
| mmu-miR-214-3p | − | 82.9 | - | ||||
| 22 | Rc3h1 | RING CCCH (C3H) domains 1 | 3 | mmu-miR-21a-5p | + | - | - |
| mmu-miR-146a-5p | + | - | - | ||||
| mmu-miR-214-3p | − | 86.3 | - | ||||
| 23 | Rimklb | ribosomal modification protein rimK-like family member B | 3 | mmu-miR-21a-3p | − | 98.7 | - |
| mmu-miR-142a-3p | − | 85.7 | - | ||||
| mmu-miR-199a-3p | − | 95.1 | - | ||||
| 24 | Smarcd1 | SWI/SNF related, matrix associated, actin dependent regulator of chromatin, subfamily d, member 1 | 3 | mmu-miR-199a-5p | − | - | −0.238 |
| mmu-miR-214-3p | − | 99.1 | - | ||||
| mmu-miR-223-3p | − | - | −0.258 | ||||
| 25 | Srsf6 | serine and arginine-rich splicing factor 6 | 3 | mmu-miR-21a-3p | − | 83.4 | −0.287 |
| mmu-miR-142a-5p | − | 98.7 | - | ||||
| mmu-miR-146a-5p | − | 88.2 | - | ||||
| 26 | Stim2 | stromal interaction molecule 2 | 3 | mmu-miR-199a-3p | − | - | −0.288 |
| mmu-miR-214-3p | − | 80.1 | - | ||||
| mmu-miR-223-3p | − | 95.0 | - | ||||
| 27 | Tm9sf3 | transmembrane 9 superfamily member 3 | 3 | mmu-miR-146a-5p | − | 85.4 | - |
| mmu-miR-199a-5p | − | - | −0.256 | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 28 | Tmem170 | transmembrane protein 170 | 3 | mmu-miR-21a-5p | − | - | −0.492 |
| mmu-miR-199a-3p | − | - | −0.293 | ||||
| mmu-miR-223-3p | − | - | −0.693 | ||||
| B | Gene Symbol | NCBI Gene Description | miRNA Target Strength | Associated miRNAs | miRTarBase 8.0 | miRDB v5.0 Score | TargetScan Mouse 7.2 Context++ Score |
| 1 | PlxnA2 | plexin A2 | 4 | mmu-miR-142a-5p | − | 91.1 | - |
| mmu-miR-199a-5p | − | 95.5 | −0.369 | ||||
| mmu-miR-214-3p | − | 82.4 | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 2 | Acvr2a | activin receptor IIA | 3 | mmu-miR-199a-3p | − | 98.6 | −0.614 |
| mmu-miR-199a-5p | − | - | −0.306 | ||||
| mmu-miR-223-3p | − | 94.2 | −0.436 | ||||
| 3 | CD2AP | CD2-associated protein | 3 | mmu-miR-142a-3p | + | - | - |
| mmu-miR-199a-3p | − | 94.5 | −0.603 | ||||
| mmu-miR-223-3p | − | - | −0.299 | ||||
| 4 | Cdk17 | cyclin-dependent kinase 17 | 3 | mmu-miR-142a-5p | − | 93.2 | - |
| mmu-miR-199a-3p | − | 88.3 | −0.342 | ||||
| mmu-miR-223-3p | − | 98.8 | −0.834 | ||||
| 5 | Celf1 | CUGBP, Elav-like family member 1 | 3 | mmu-miR-199a-3p | − | - | −0.253 |
| mmu-miR-214-3p | − | 81.0 | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 6 | Crebrf | CREB3 regulatory factor | 3 | mmu-miR-199a-3p | − | - | −0.220 |
| mmu-miR-199a-5p | − | - | −0.213 | ||||
| mmu-miR-223-3p | − | 80.4 | - | ||||
| 7 | Fam199x | family with sequence similarity 199, X-linked | 3 | mmu-miR-142a-5p | − | 84.4 | - |
| mmu-miR-199a-3p | − | 96.0 | −0.485 | ||||
| mmu-miR-223-3p | − | 93.5 | - | ||||
| 8 | Il6 | interleukin 6 | 3 | mmu-miR-142a-3p | + | - | - |
| mmu-miR-146a-5p | + | - | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 9 | Kras | Kirsten rat sarcoma viral oncogene homolog | 3 | mmu-miR-142a-5p | − | 92.5 | - |
| mmu-miR-146a-5p | − | 88.6 | - | ||||
| mmu-miR-223-3p | + | - | - | ||||
| 10 | Lrrc1 | leucine rich repeat containing 1 | 3 | mmu-miR-142a-3p | − | 92.9 | - |
| mmu-miR-199a-3p | − | - | −0.272 | ||||
| mmu-miR-214-3p | − | 81.7 | - | ||||
| 11 | Pip5k1b | phosphatidylinositol-4-phosphate 5-kinase, type 1 beta | 3 | mmu-miR-142a-5p | − | - | −0.202 |
| mmu-miR-146a-5p | − | - | −0.236 | ||||
| mmu-miR-199a-3p | − | 88.7 | −0.319 | ||||
| 12 | Ptpn4 | protein tyrosine phosphatase, non-receptor type 4 | 3 | mmu-miR-142a-5p | − | 97.9 | - |
| mmu-miR-199a-3p | − | - | −0.274 | ||||
| mmu-miR-199a-5p | − | - | −0.253 | ||||
| 13 | Smarcd1 | SWI/SNF related, matrix associated, actin dependent regulator of chromatin, subfamily d, member 1 | 3 | mmu-miR-199a-5p | − | - | −0.238 |
| mmu-miR-214-3p | − | 99.1 | - | ||||
| mmu-miR-223-3p | − | - | −0.258 | ||||
| 14 | Stim2 | stromal interaction molecule 2 | 3 | mmu-miR-199a-3p | − | - | −0.288 |
| mmu-miR-214-3p | − | 80.1 | - | ||||
| mmu-miR-223-3p | − | 95.0 | - | ||||
| 15 | Tm9sf3 | transmembrane 9 superfamily member 3 | 3 | mmu-miR-146a-5p | − | 85.4 | - |
| mmu-miR-199a-5p | − | - | −0.256 | ||||
| mmu-miR-223-3p | + | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Róka, B.; Tod, P.; Kaucsár, T.; Bukosza, É.N.; Vörös, I.; Varga, Z.V.; Petrovich, B.; Ágg, B.; Ferdinandy, P.; Szénási, G.; et al. Delayed Contralateral Nephrectomy Halted Post-Ischemic Renal Fibrosis Progression and Inhibited the Ischemia-Induced Fibromir Upregulation in Mice. Biomedicines 2021, 9, 815. https://doi.org/10.3390/biomedicines9070815
Róka B, Tod P, Kaucsár T, Bukosza ÉN, Vörös I, Varga ZV, Petrovich B, Ágg B, Ferdinandy P, Szénási G, et al. Delayed Contralateral Nephrectomy Halted Post-Ischemic Renal Fibrosis Progression and Inhibited the Ischemia-Induced Fibromir Upregulation in Mice. Biomedicines. 2021; 9(7):815. https://doi.org/10.3390/biomedicines9070815
Chicago/Turabian StyleRóka, Beáta, Pál Tod, Tamás Kaucsár, Éva Nóra Bukosza, Imre Vörös, Zoltán V. Varga, Balázs Petrovich, Bence Ágg, Péter Ferdinandy, Gábor Szénási, and et al. 2021. "Delayed Contralateral Nephrectomy Halted Post-Ischemic Renal Fibrosis Progression and Inhibited the Ischemia-Induced Fibromir Upregulation in Mice" Biomedicines 9, no. 7: 815. https://doi.org/10.3390/biomedicines9070815
APA StyleRóka, B., Tod, P., Kaucsár, T., Bukosza, É. N., Vörös, I., Varga, Z. V., Petrovich, B., Ágg, B., Ferdinandy, P., Szénási, G., & Hamar, P. (2021). Delayed Contralateral Nephrectomy Halted Post-Ischemic Renal Fibrosis Progression and Inhibited the Ischemia-Induced Fibromir Upregulation in Mice. Biomedicines, 9(7), 815. https://doi.org/10.3390/biomedicines9070815