
Biological Role of the 3β-Corner Structural Motif in Proteins

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
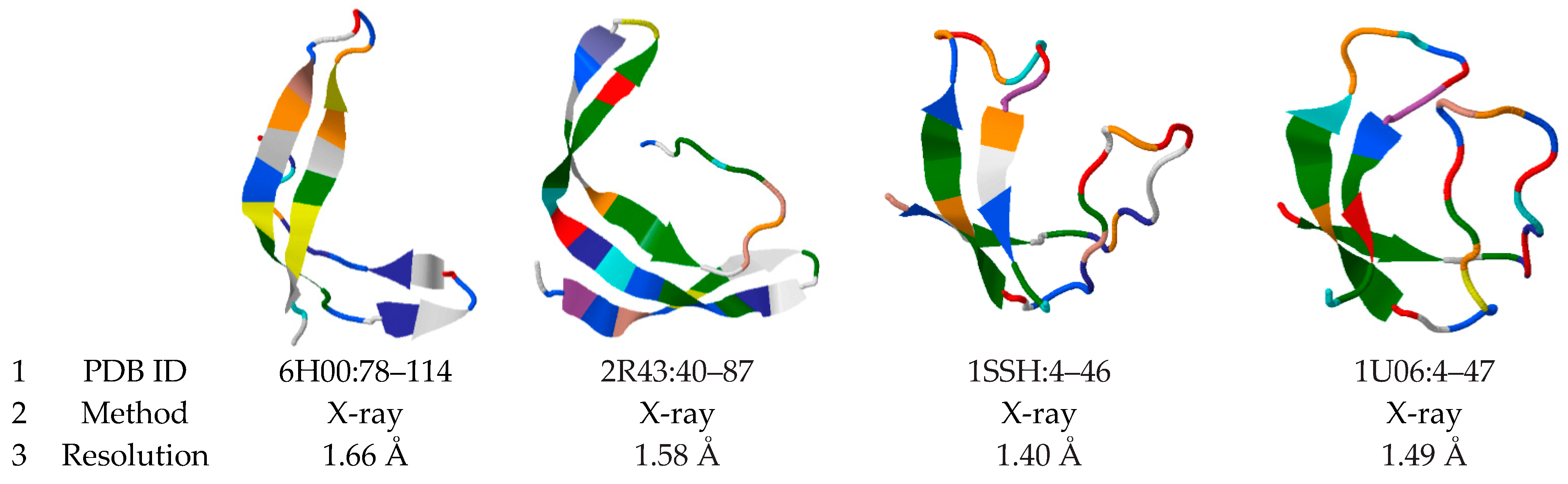
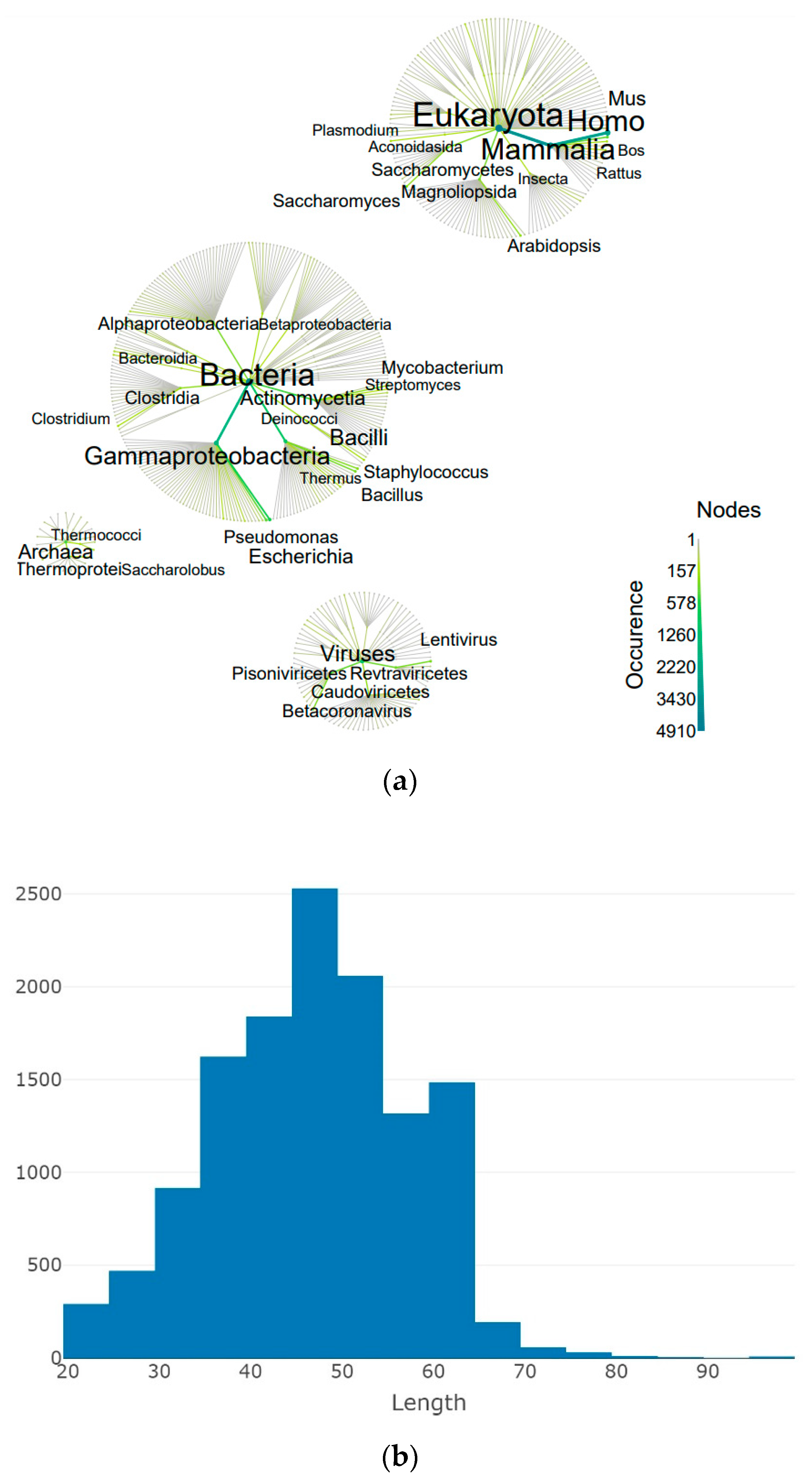
3.1. Occurrence of the 3β-Corner Motif in Proteins
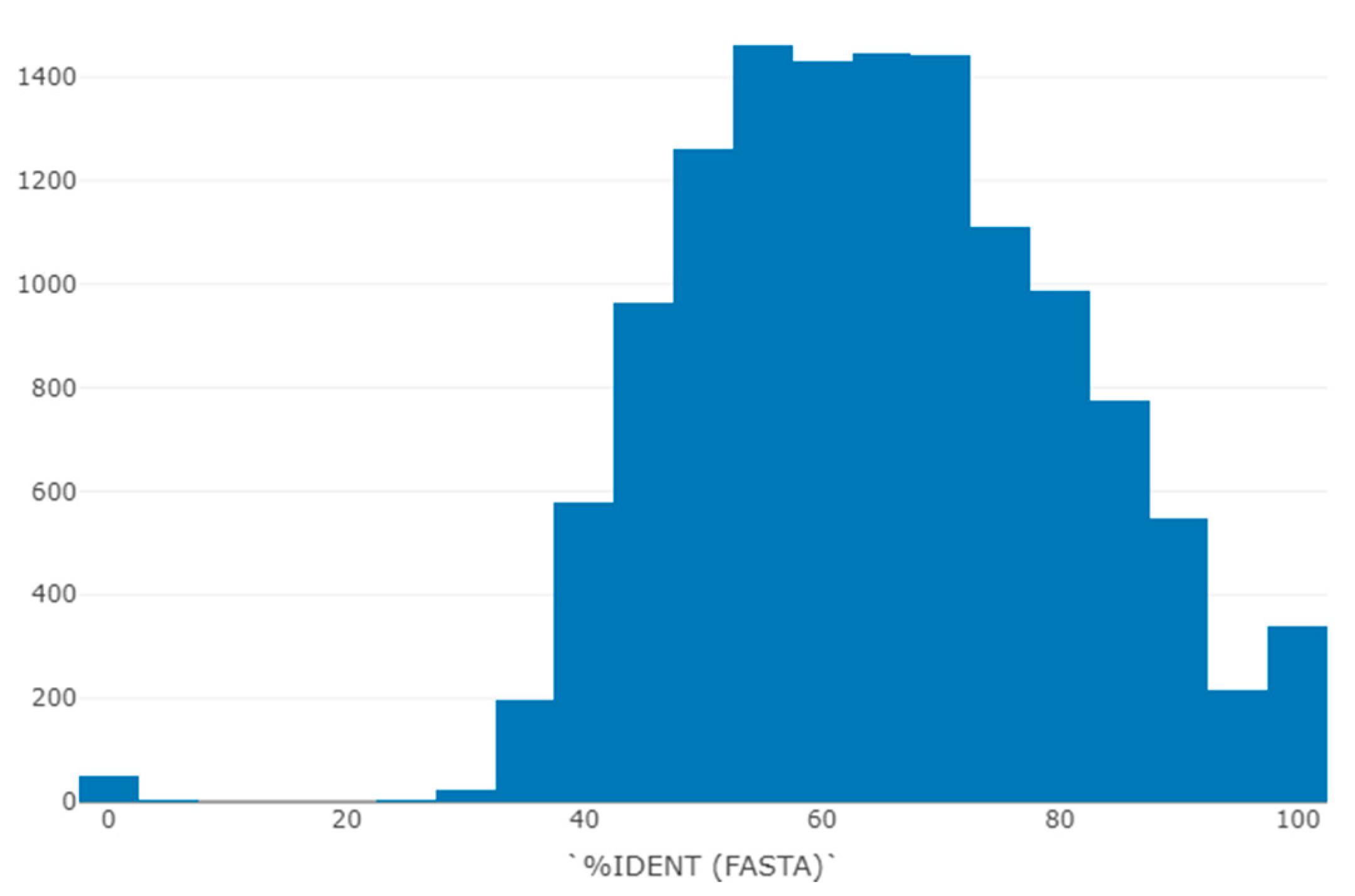
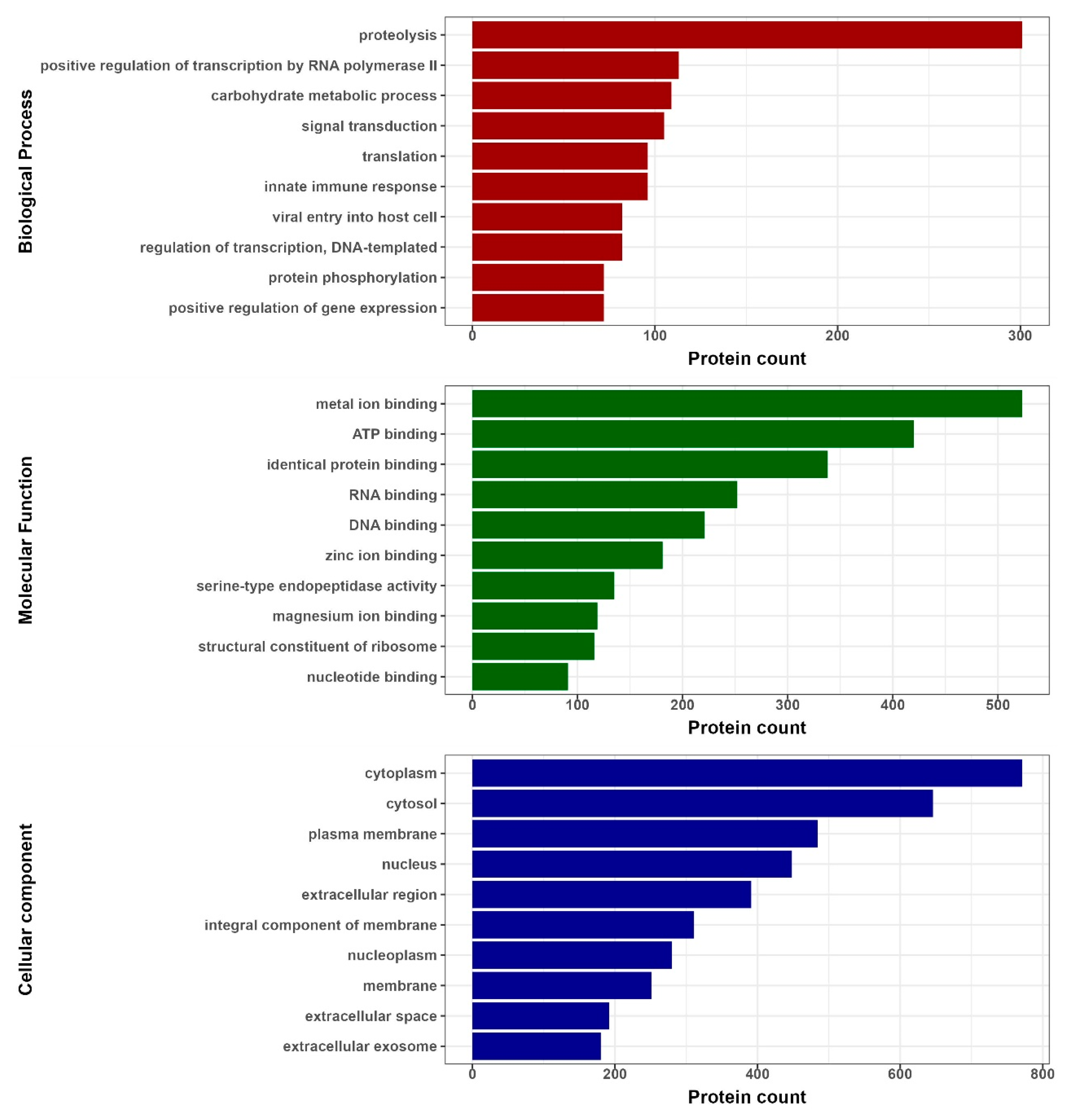
3.2. Protein Type and Fold Diversity Containing 3β-Corners
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Efimov, A.V. A Novel Super-Secondary Structure of Beta-Proteins. A Triple-Strand Corner. FEBS Lett. 1992, 298, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Remmert, M.; Biegert, A.; Linke, D.; Lupas, A.N.; Söding, J. Evolution of Outer Membrane Beta-Barrels from an Ancestral Beta Beta Hairpin. Mol. Biol. Evol. 2010, 27, 1348–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairman, J.W.; Noinaj, N.; Buchanan, S.K. The Structural Biology of β-Barrel Membrane Proteins: A Summary of Recent Reports. Curr. Opin. Struct. Biol. 2011, 21, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.H.; Aulakh, S.; Paetzel, M. The Bacterial Outer Membrane β-Barrel Assembly Machinery. Protein Sci. Publ. Protein Soc. 2012, 21, 751–768. [Google Scholar] [CrossRef] [Green Version]
- Rollauer, S.E.; Sooreshjani, M.A.; Noinaj, N.; Buchanan, S.K. Outer Membrane Protein Biogenesis in Gram-Negative Bacteria. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20150023. [Google Scholar] [CrossRef] [Green Version]
- Noinaj, N.; Gumbart, J.C.; Buchanan, S.K. The β-Barrel Assembly Machinery in Motion. Nat. Rev. Microbiol. 2017, 15, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Tsirigos, K.D.; Elofsson, A.; Bagos, P.G. PRED-TMBB2: Improved Topology Prediction and Detection of Beta-Barrel Outer Membrane Proteins. Bioinforma. Oxf. Engl. 2016, 32, i665–i671. [Google Scholar] [CrossRef] [Green Version]
- Serra-Batiste, M.; Ninot-Pedrosa, M.; Puig, E.; Ciudad, S.; Gairí, M.; Carulla, N. Preparation of a Well-Defined and Stable β-Barrel Pore-Forming Aβ42 Oligomer. Methods Mol. Biol. Clifton NJ 2018, 1779, 13–22. [Google Scholar] [CrossRef]
- Durell, S.R.; Guy, H.R. The Amyloid Concentric β-Barrel Hypothesis: Models of Synuclein Oligomers, Annular Protofibrils, Lipoproteins, and Transmembrane Channels. Proteins 2022, 90, 512–542. [Google Scholar] [CrossRef]
- Züst, R.; Cervantes-Barragán, L.; Kuri, T.; Blakqori, G.; Weber, F.; Ludewig, B.; Thiel, V. Coronavirus Non-Structural Protein 1 Is a Major Pathogenicity Factor: Implications for the Rational Design of Coronavirus Vaccines. PLoS Pathog. 2007, 3, e109. [Google Scholar] [CrossRef]
- Wathelet, M.G.; Orr, M.; Frieman, M.B.; Baric, R.S. Severe Acute Respiratory Syndrome Coronavirus Evades Antiviral Signaling: Role of Nsp1 and Rational Design of an Attenuated Strain. J. Virol. 2007, 81, 11620–11633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semper, C.; Watanabe, N.; Savchenko, A. Structural Characterization of Nonstructural Protein 1 from SARS-CoV-2. iScience 2021, 24, 101903. [Google Scholar] [CrossRef] [PubMed]
- Chothia, C.; Janin, J. Orthogonal Packing of Beta-Pleated Sheets in Proteins. Biochemistry 1982, 21, 3955–3965. [Google Scholar] [CrossRef] [PubMed]
- Efimov, A.V. A Structural Tree for Proteins Containing 3beta-Corners. FEBS Lett. 1997, 407, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Boshkova, E.A.; Efimov, A.V. Structures Closed into Cycles in Proteins Containing 3β-Corners. Biochem. Biokhimiia 2010, 75, 1258–1263. [Google Scholar] [CrossRef]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 13 September 2022).
- Foster, Z.S.L.; Sharpton, T.J.; Grünwald, N.J. Metacoder: An R Package for Visualization and Manipulation of Community Taxonomic Diversity Data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [Green Version]
- Efimov, A.V. Structural Similarity between Two-Layer Alpha/Beta and Beta-Proteins. J. Mol. Biol. 1995, 245, 402–415. [Google Scholar] [CrossRef]
- Rudnev, V.R.; Nikolsky, K.S.; Petrovsky, D.V.; Kulikova, L.I.; Kargatov, A.M.; Malsagova, K.A.; Stepanov, A.A.; Kopylov, A.T.; Kaysheva, A.L.; Efimov, A.V. 3β-Corner Stability by Comparative Molecular Dynamics Simulations. Int. J. Mol. Sci. 2022, 23, 11674. [Google Scholar] [CrossRef] [PubMed]
- Höhr, A.I.C.; Straub, S.P.; Warscheid, B.; Becker, T.; Wiedemann, N. Assembly of β-Barrel Proteins in the Mitochondrial Outer Membrane. Biochim. Biophys. Acta BBA Mol. Cell Res. 2015, 1853, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, D.; Mahalakshmi, R. Transmembrane β-Barrels: Evolution, Folding and Energetics. Biochim. Biophys. Acta BBA Biomembr. 2017, 1859, 2467–2482. [Google Scholar] [CrossRef]
- Jores, T.; Rapaport, D. Early Stages in the Biogenesis of Eukaryotic β-Barrel Proteins. FEBS Lett. 2017, 591, 2671–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulatskaya, A.I.; Kosolapova, A.O.; Bobylev, A.G.; Belousov, M.V.; Antonets, K.S.; Sulatsky, M.I.; Kuznetsova, I.M.; Turoverov, K.K.; Stepanenko, O.V.; Nizhnikov, A.A. β-Barrels and Amyloids: Structural Transitions, Biological Functions, and Pathogenesis. Int. J. Mol. Sci. 2021, 22, 11316. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, U.; Roske, Y. Cold-Shock Domains—Abundance, Structure, Properties, and Nucleic-Acid Binding. Cancers 2021, 13, 190. [Google Scholar] [CrossRef]
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, Purification and Properties of Aequorin, a Bioluminescent Protein from the Luminous Hydromedusan, Aequorea. J. Cell. Comp. Physiol. 1962, 59, 223–239. [Google Scholar] [CrossRef]
- Current and Potential Biotechnological Applications of Odorant-Binding Proteins-PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/32888038/ (accessed on 13 September 2022).
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.-K.; Yadav, D.; Sharma, N.; Jin, J.-O. Dipeptidyl Peptidase (DPP)-IV Inhibitors with Antioxidant Potential Isolated from Natural Sources: A Novel Approach for the Management of Diabetes. Pharmaceuticals 2021, 14, 586. [Google Scholar] [CrossRef]
- Iida, H.; Ishii, E.; Nakahara, M.; Urata, M.; Wakiyama, M.; Kurihara, M.; Watanabe, K.; Kai, T.; Ihara, K.; Kinoshita, S.; et al. A Case of Congenital Afibrinogenemia: Fibrinogen Hakata, a Novel Nonsense Mutation of the Fibrinogen Gamma-Chain Gene. Thromb. Haemost. 2000, 84, 49–53. [Google Scholar] [CrossRef]
- Tamayo-Velasco, Á.; Cebeira, M.J.; Bombín-Canal, C.; Acevedo-García, R.M.; Peñarrubia-Ponce, M.J. Fibrinogen Deficiency with Thrombotic Manifestations. Eur. J. Case Rep. Intern. Med. 2022, 9, 003400. [Google Scholar] [CrossRef]
- Ding, X.; Hou, L.; Zhang, H.; Chen, Z.; Liu, Z.; Gong, J.; Tang, Z.; Hu, R. EIF3C Promotes Lung Cancer Tumorigenesis by Regulating the APP/HSPA1A/LMNB1 Axis. Dis. Markers 2022, 2022, 9464094. [Google Scholar] [CrossRef]
- Oroń, M.; Grochowski, M.; Jaiswar, A.; Legierska, J.; Jastrzębski, K.; Nowak-Niezgoda, M.; Kołos, M.; Kaźmierczak, W.; Olesiński, T.; Lenarcik, M.; et al. The Molecular Network of the Proteasome Machinery Inhibition Response Is Orchestrated by HSP70, Revealing Vulnerabilities in Cancer Cells. Cell Rep. 2022, 40, 111428. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Y.; Chen, D.; Lee, T.H. Peptidyl-Prolyl Cis/Trans Isomerase Pin1 and Alzheimer’s Disease. Front. Cell Dev. Biol. 2020, 8, 355. [Google Scholar] [CrossRef] [PubMed]
- Fagiani, F.; Govoni, S.; Racchi, M.; Lanni, C. The Peptidyl-Prolyl Isomerase Pin1 in Neuronal Signaling: From Neurodevelopment to Neurodegeneration. Mol. Neurobiol. 2021, 58, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- D’Azzo, A.; Machado, E.; Annunziata, I. Pathogenesis, Emerging Therapeutic Targets and Treatment in Sialidosis. Expert Opin. Orphan Drugs 2015, 3, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Aso, K.; Soutome, T.; Satoh, M.; Aoki, T.; Ogura, H.; Yamamoto, T.; Kanno, H.; Takahashi, H. Association of Autosomal-Recessive-Type Distal Renal Tubular Acidosis and Glanzmann Thrombasthenia as a Consequence of Runs of Homozygosity. Clin. Case Rep. 2022, 10, e6070. [Google Scholar] [CrossRef] [PubMed]
- Rappold, I.; Iwabuchi, K.; Date, T.; Chen, J. Tumor Suppressor P53 Binding Protein 1 (53BP1) Is Involved in DNA Damage-Signaling Pathways. J. Cell Biol. 2001, 153, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braithwaite, A.W.; Del Sal, G.; Lu, X. Some P53-Binding Proteins That Can Function as Arbiters of Life and Death. Cell Death Differ. 2006, 13, 984–993. [Google Scholar] [CrossRef] [Green Version]
- Petro, T.M. IFN Regulatory Factor 3 in Health and Disease. J. Immunol. Baltim. Md. 1950 2020, 205, 1981–1989. [Google Scholar] [CrossRef]
- Matta, B.; Song, S.; Li, D.; Barnes, B.J. Interferon Regulatory Factor Signaling in Autoimmune Disease. Cytokine 2017, 98, 15–26. [Google Scholar] [CrossRef]
- Krook, M.A.; Reeser, J.W.; Ernst, G.; Barker, H.; Wilberding, M.; Li, G.; Chen, H.-Z.; Roychowdhury, S. Fibroblast Growth Factor Receptors in Cancer: Genetic Alterations, Diagnostics, Therapeutic Targets and Mechanisms of Resistance. Br. J. Cancer 2021, 124, 880–892. [Google Scholar] [CrossRef]
- Jain, V.K.; Turner, N.C. Challenges and Opportunities in the Targeting of Fibroblast Growth Factor Receptors in Breast Cancer. Breast Cancer Res. 2012, 14, 208. [Google Scholar] [CrossRef]
- Kaelin, W.G. The Jeremiah Metzger Lecture: Von Hippel-Lindau Disease: Insights Into Oxygen Sensing, Cancer and Drugging the Undruggable. Trans. Am. Clin. Climatol. Assoc. 2022, 132, 170–181. [Google Scholar] [PubMed]
- Dou, J.; Vorobieva, A.A.; Sheffler, W.; Doyle, L.A.; Park, H.; Bick, M.J.; Mao, B.; Foight, G.W.; Lee, M.Y.; Gagnon, L.A.; et al. De Novo Design of a Fluorescence-Activating β-Barrel. Nature 2018, 561, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Vorobieva, A.A.; White, P.; Liang, B.; Horne, J.E.; Bera, A.K.; Chow, C.M.; Gerben, S.; Marx, S.; Kang, A.; Stiving, A.Q.; et al. De Novo Design of Transmembrane β Barrels. Science 2021, 371, eabc8182. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt ID | Protein Name | PDB ID (Locus, a.a.) | SCOPE Fold | % Ident * | Core Fold | Disease ** | Class of Protein | Ref. |
|---|---|---|---|---|---|---|---|---|
| P27487 | Dipeptidyl peptidase 4 | 1R9N (402–446) | b.70 | 95.1 | beta-propeller | Carbohydrate metabolic disorder, type 2 diabetes mellitus | All beta proteins | [27,28] |
| P02679 | Fibrinogen gamma chain | 3E1I (F242–279) | d.171 | 94.1 | unusual fold | Congenital afibrinogenemia | Alpha and beta proteins (a + b) | [29,30] |
| P0DMV8 | Heat shock 70 kDa protein 1A | 1S3X (64–116) | c.55 | 93.9 | beta-layer | Cancer | Alpha and beta proteins (a/b) | [31,32] |
| P30405 | Peptidyl-prolyl cis-trans isomerase F | 4J5A (45–108) | b.62 | 95.0 | barrel, closed | Neurodegenerative disease | All beta proteins | [33,34] |
| Q9Y3R4 | Sialidase-2 | 2F24 (163–210) | b.68 | 93.2 | beta-propeller | Glycoproteinosis | All beta proteins | [35] |
| P08514 | Integrin, alpha 2b | 3ZDY (218–272) | b.69 | 92.2 | beta-propeller | Glanzmann’s thrombasthenia | All beta proteins | [36] |
| Q12888 | Tumor protein p53 binding protein 1 | 3LGF (1540–1584) | b.34 | 85.4 | barrel, partly opened | Cancer | All beta proteins | [37,38] |
| Q14653 | Interferon regulatory factor 3 | 1J2F (201–246) | b.26 | 85.2 | sandwich | Autoimmune disease | All beta proteins | [39,40] |
| P05230 | Fibroblast growth factor 1 | 3UD8 (19–60) | b.42 | 84.2 | barrel, closed | Cancer | All beta proteins | [41,42] |
| P40337 | E3 ubiquitin protein ligase | 5NW2 (114–154) | b.3 | 83.8 | sandwich | Hemangioma | All beta proteins | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudnev, V.R.; Petrovsky, D.V.; Nikolsky, K.S.; Kulikova, L.I.; Stepanov, A.A.; Malsagova, K.A.; Kaysheva, A.L.; Efimov, A.V. Biological Role of the 3β-Corner Structural Motif in Proteins. Processes 2022, 10, 2159. https://doi.org/10.3390/pr10112159
Rudnev VR, Petrovsky DV, Nikolsky KS, Kulikova LI, Stepanov AA, Malsagova KA, Kaysheva AL, Efimov AV. Biological Role of the 3β-Corner Structural Motif in Proteins. Processes. 2022; 10(11):2159. https://doi.org/10.3390/pr10112159
Chicago/Turabian StyleRudnev, Vladimir R., Denis V. Petrovsky, Kirill S. Nikolsky, Liudmila I. Kulikova, Alexander A. Stepanov, Kristina A. Malsagova, Anna L. Kaysheva, and Alexander V. Efimov. 2022. "Biological Role of the 3β-Corner Structural Motif in Proteins" Processes 10, no. 11: 2159. https://doi.org/10.3390/pr10112159
APA StyleRudnev, V. R., Petrovsky, D. V., Nikolsky, K. S., Kulikova, L. I., Stepanov, A. A., Malsagova, K. A., Kaysheva, A. L., & Efimov, A. V. (2022). Biological Role of the 3β-Corner Structural Motif in Proteins. Processes, 10(11), 2159. https://doi.org/10.3390/pr10112159