
Screening of Azotobacter, Bacillus and Pseudomonas Species as Plant Growth-Promoting Bacteria

 ,
, 
 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
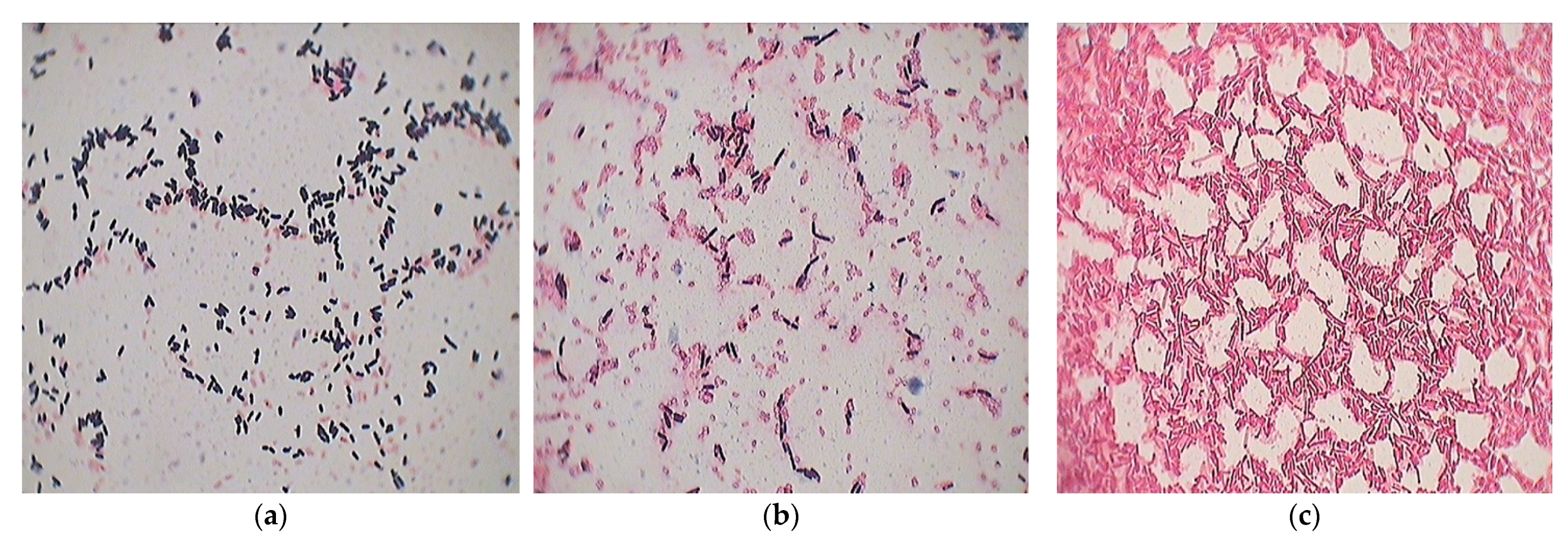
2.1. Isolation and Identification of Bacteria Genus
2.2. Soil Sterilization
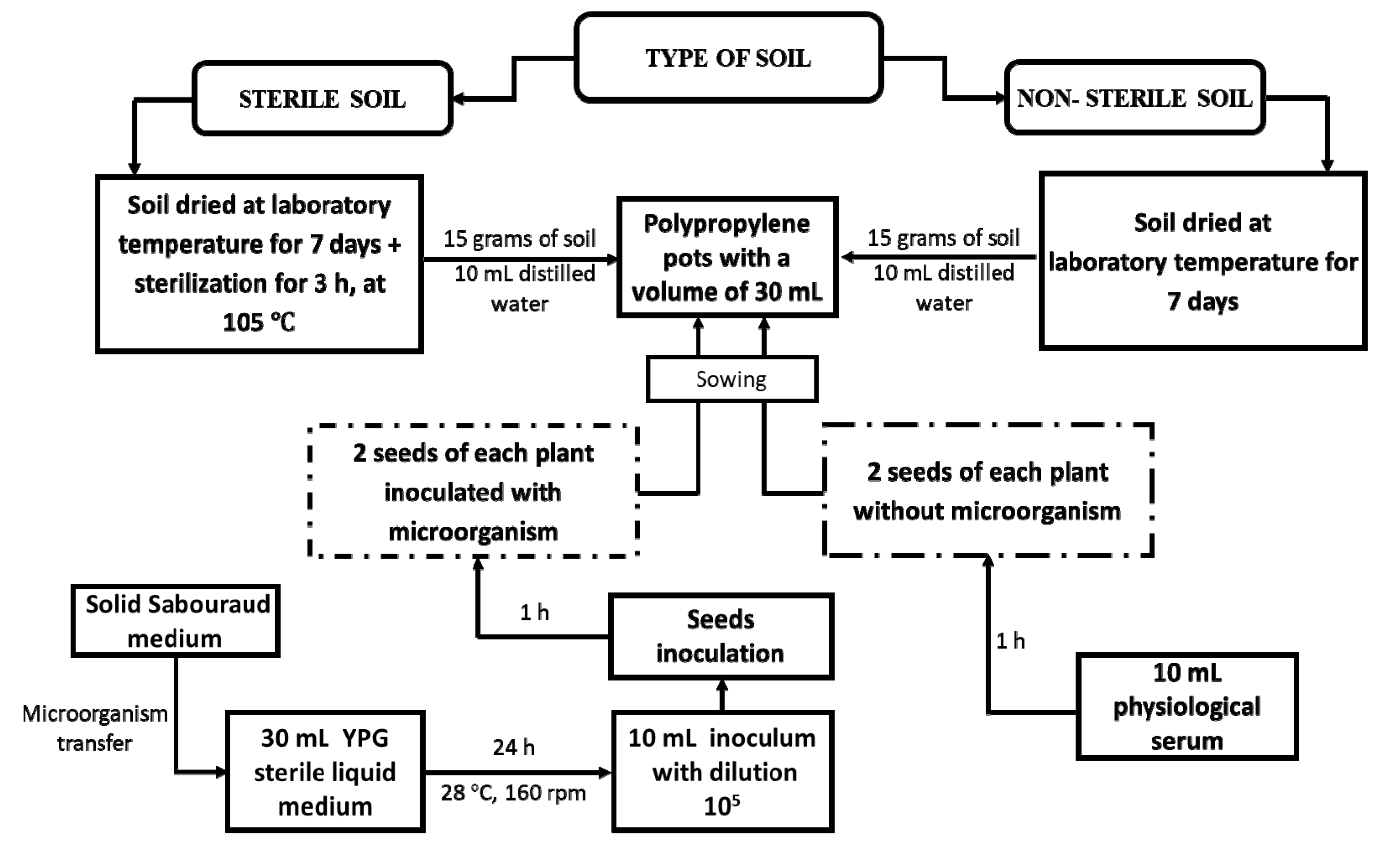
2.3. Experimental Protocol Used for Synergism Studies
2.4. Plant Length and Dry Biomass Measurement
2.5. Statistical Analysis
3. Results
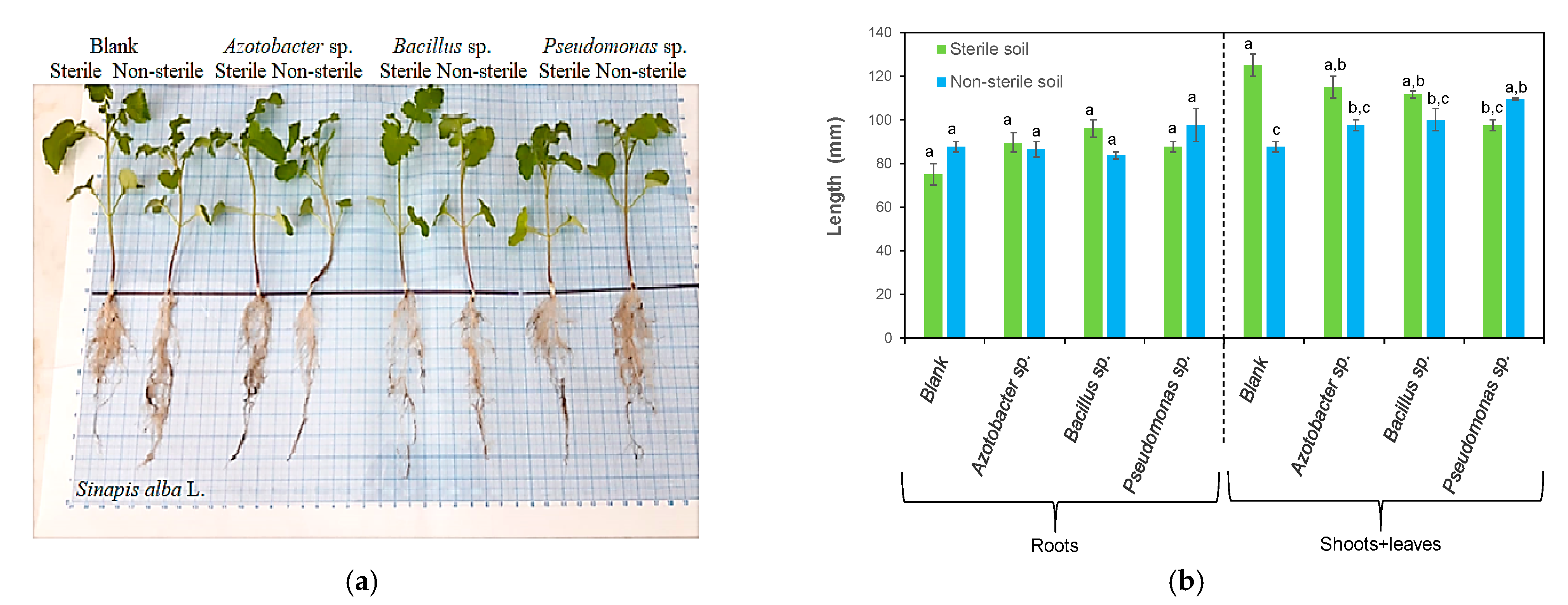
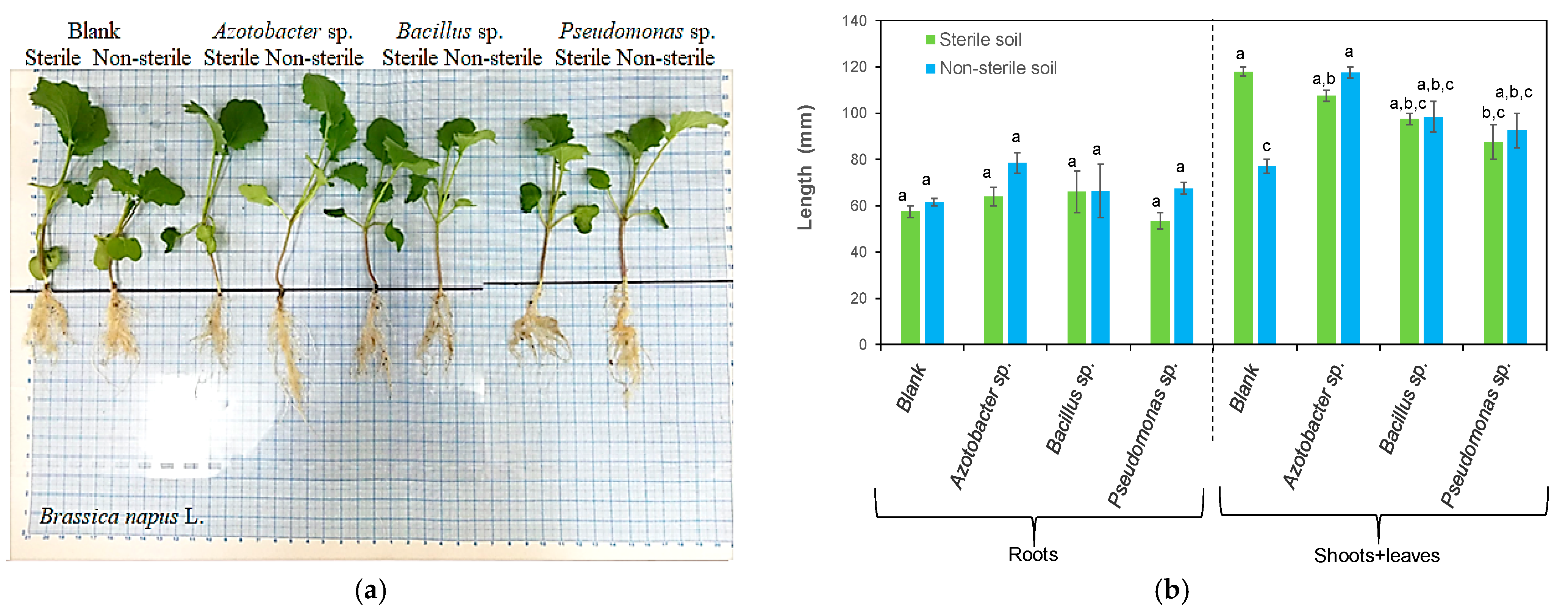
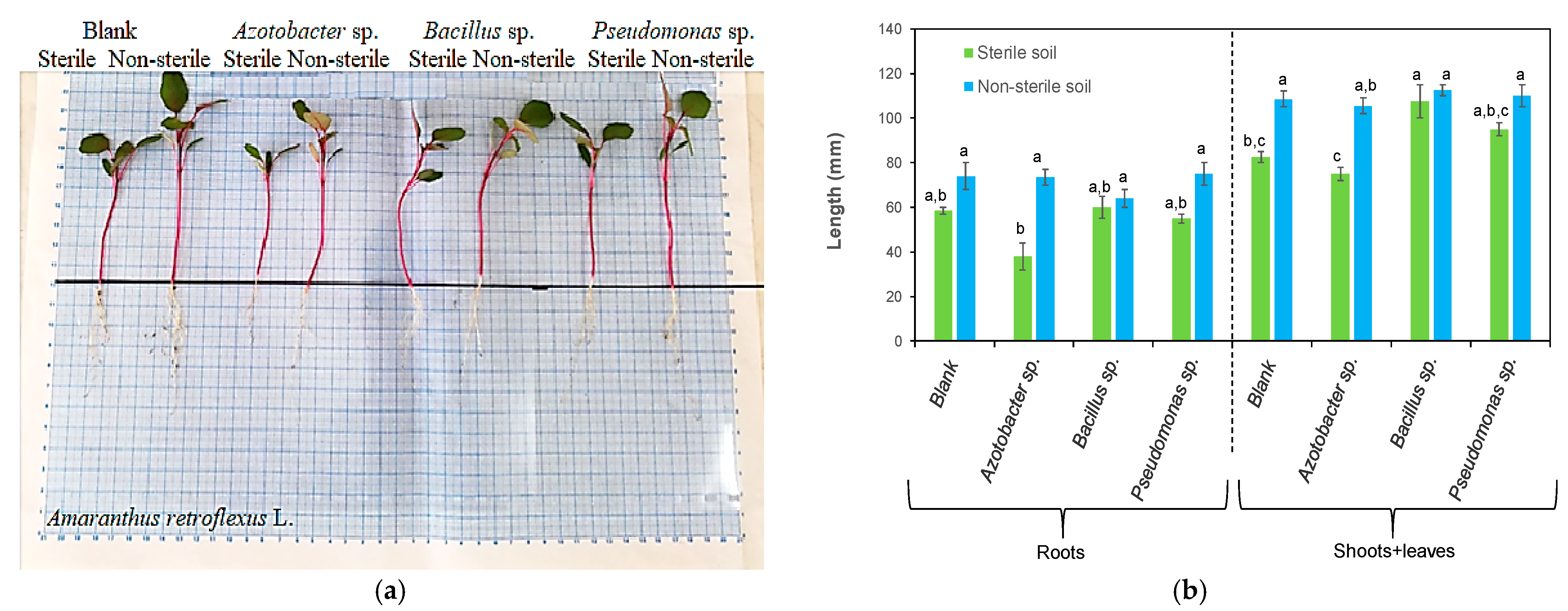
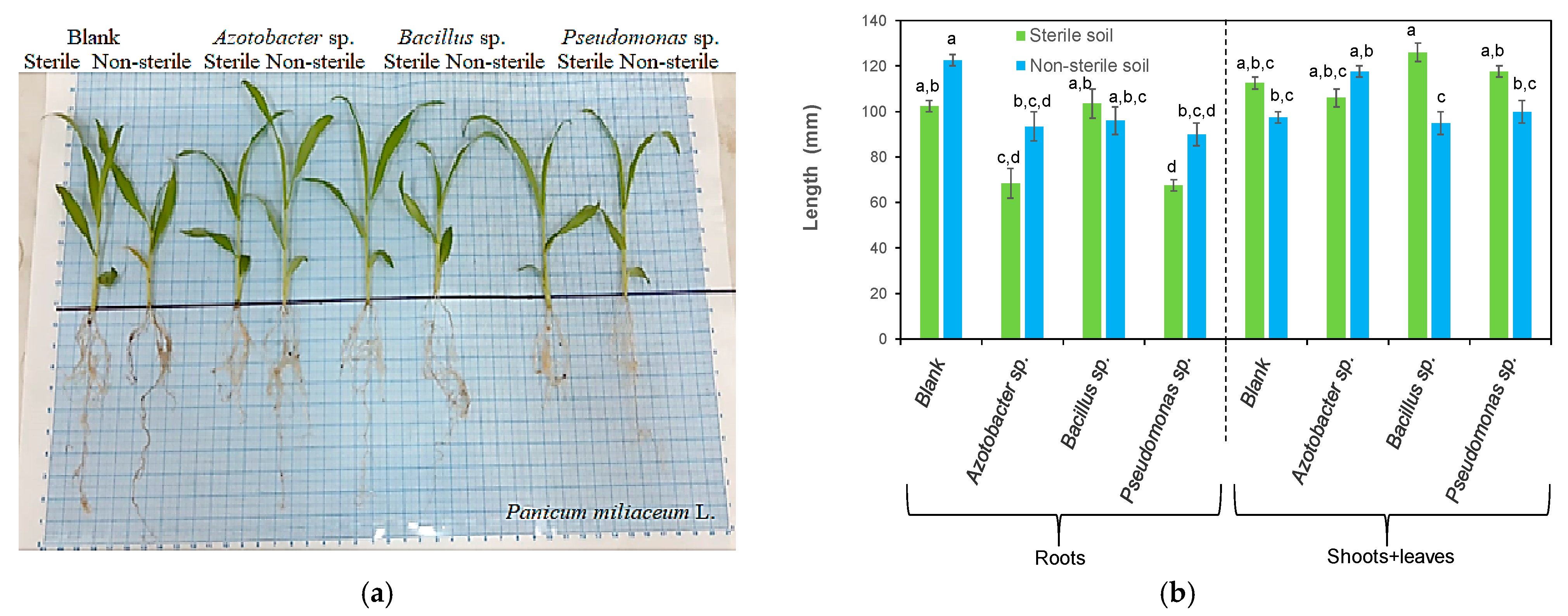
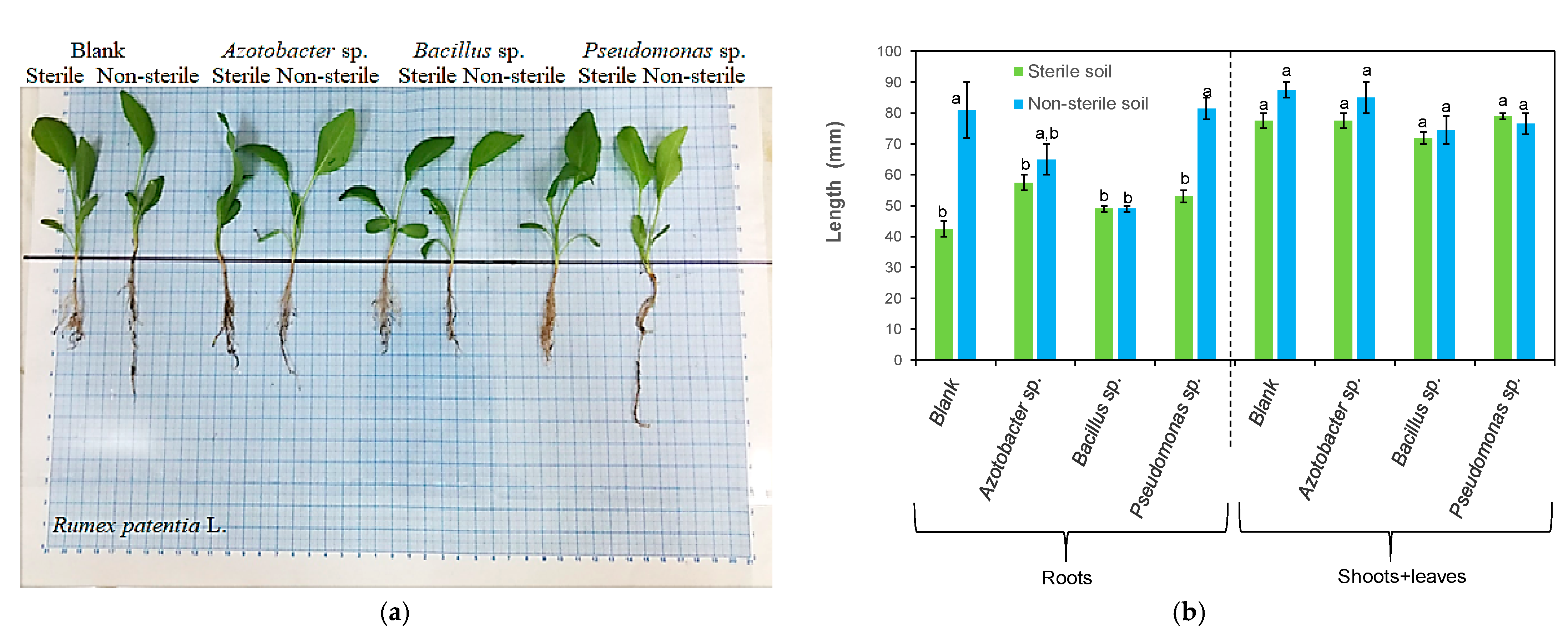
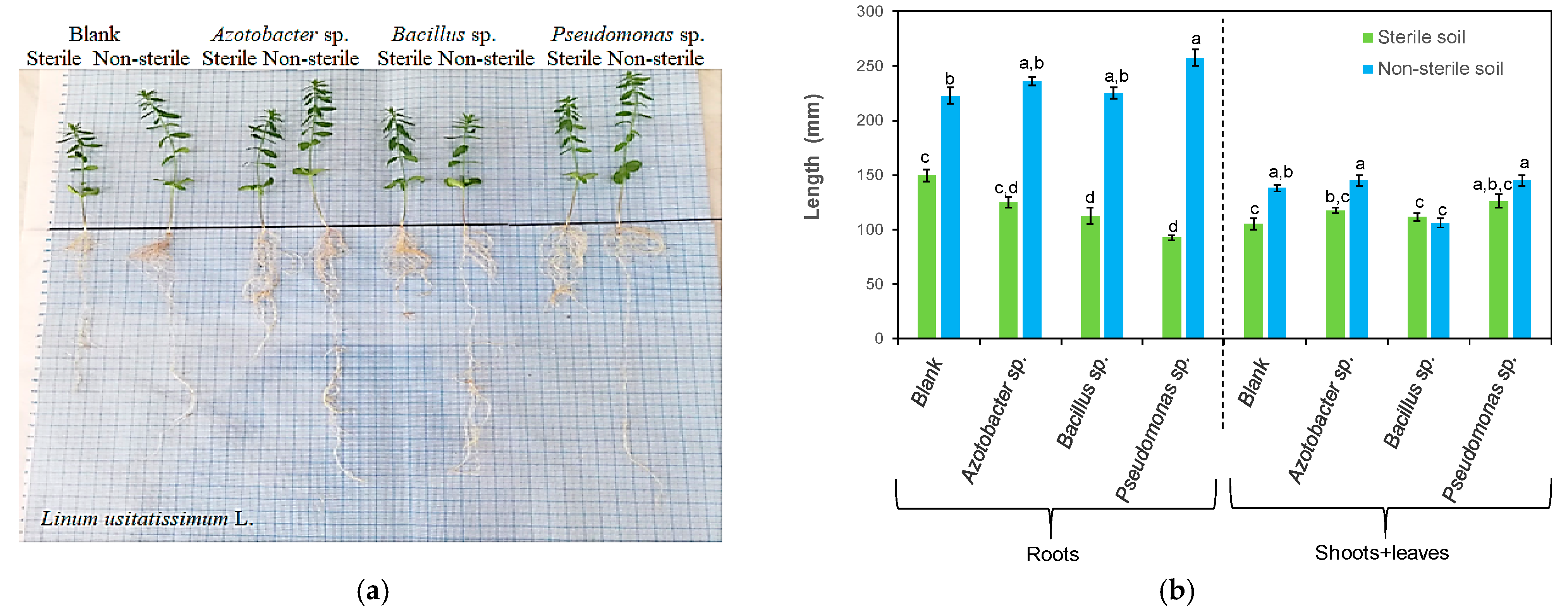
3.1. Effects of Bacteria on Plant Length
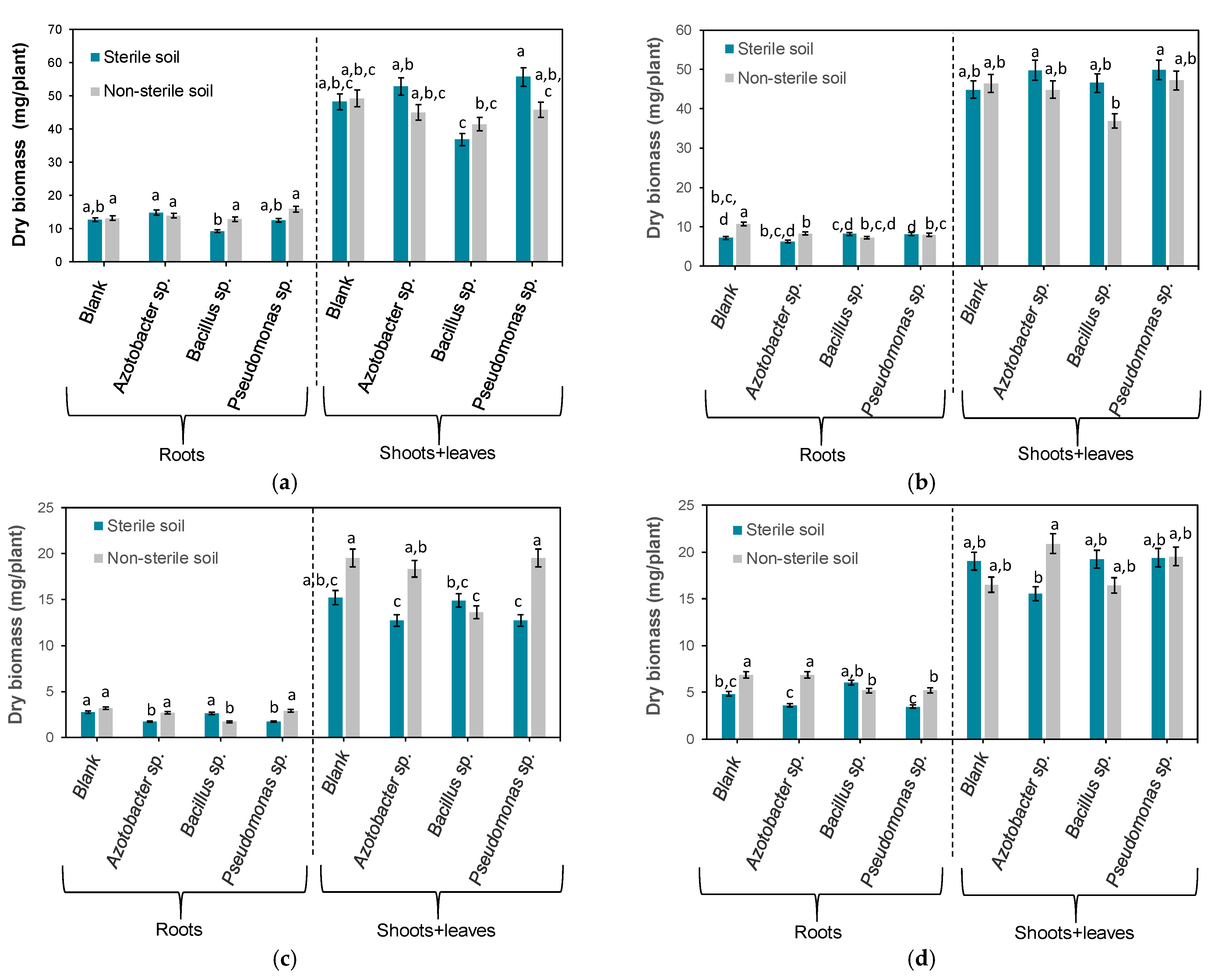
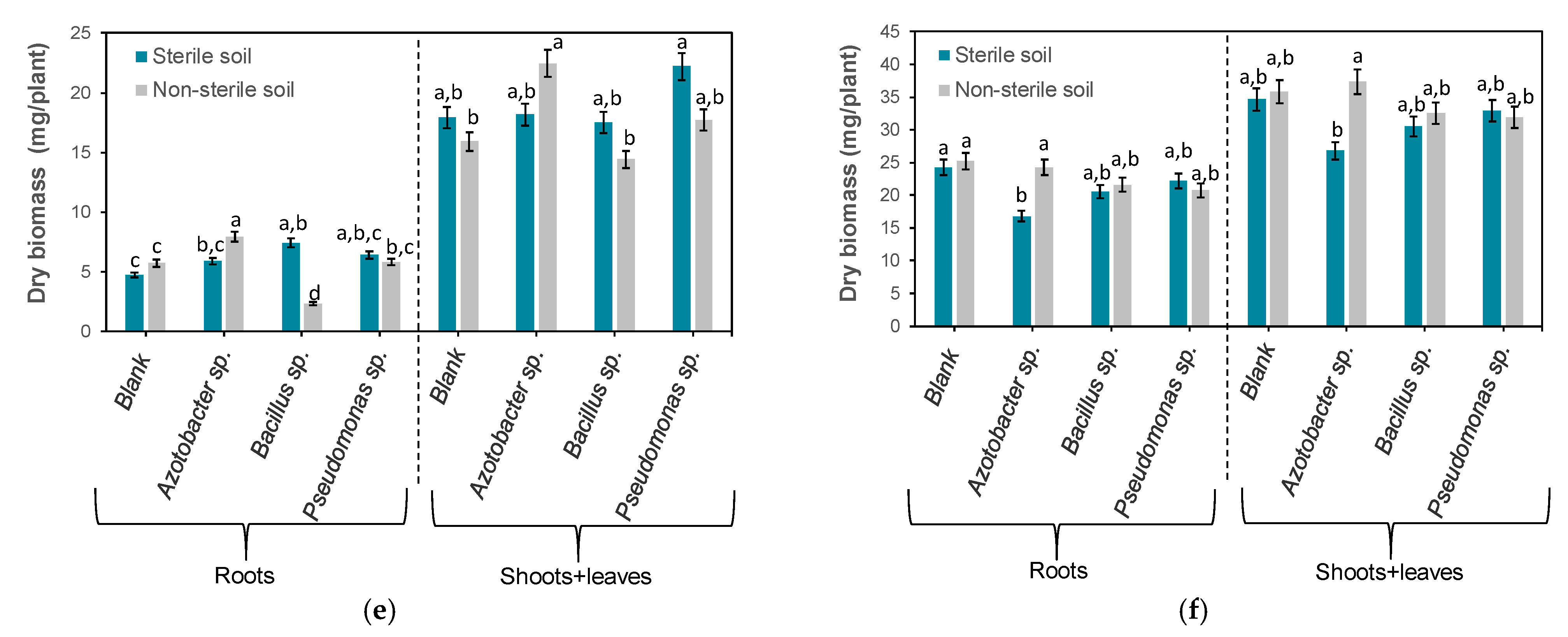
3.2. Effects of Bacteria on Plant Dry Biomass
3.3. Statistical Analysis of Experimental Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Diaconu, M.; Vasile Pavel, L.; Hlihor, R.-M.; Rosca, M.; Fertu, D.I.; Lenz, M.; Corvini, P.X.; Gavrilescu, M. Characterization of heavy metal toxicity in some plants and microorganisms—A preliminary approach for environmental bioremediation. New Biotechnol. 2020, 56, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; O’Connor, D.; Igalavithana, A.D.; Alessi, D.S.; Luo, J.; Tsang, D.C.W.; Sparks, D.L.; Yamauchi, Y.; Rinklebe, J.; Ok, Y.S. Metal contamination and bioremediation of agricultural soils for food safety and sustainability. Nat. Rev. Earth Environ. 2020, 1, 366–381. [Google Scholar] [CrossRef]
- Hlihor, R.M.; Cozma, P.; Gavrilescu, M. Removal of Heavy Metals from the Environment by Phytoremediation and Microbial Remediation. In Sustainable Solutions for Environmental Pollution; Scrivener Publishing: Beverly, MA, USA, 2022; pp. 95–146. [Google Scholar] [CrossRef]
- Gavrilescu, M. Enhancing phytoremediation of soils polluted with heavy metals. Curr. Opin. Biotechnol. 2022, 74, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Altamuiri, A.; Aladwani, S.; Almutairi, B. Soil temperature profile investigation under arid climate of Kuwait using mechanistic and mixed models. Environ. Eng. Manag. J. 2021, 20, 1183–1192. [Google Scholar]
- Bulgariu, D.; Bulgariu, L. Sustainable Utilization of Marine Algae Biomass for Environmental Bioremediation. In Prospects and Challenges in Algal Biotechnology; Tripathi, B.N., Kumar, D., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2017; pp. 179–217. [Google Scholar] [CrossRef]
- Hlihor, R.M.; Apostol, L.C.; Smaranda, C.; Pavel, L.V.; Căliman, F.A.; Robu, B.M.; Gavrilescu, M. Bioavailability processes for contaminants in soils and their use in risk assessment. Environ. Eng. Manag. J. 2009, 8, 1199–1206. [Google Scholar]
- Nadeem, S.M.; Ahmad, M.; Zahir, A.Z.; Javaid, A.; Ahraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef]
- Vincze, É.B.; Salamon, R.V.; Kovács, E.; Mara, G. Effect of metal tolerant plant growth promoting rhizobacteria on bean growth cadmium and zinc uptake and stress responses. Environ. Eng. Manag. J. 2018, 17, 803–811. [Google Scholar]
- NRC. Assessment of Ecotoxicity. In A Framework to Guide Selection of Chemical Alternatives; The National Academies Press, National Research Council: Washington, DC, USA, 2014; pp. 81–92. [Google Scholar]
- Manoj, S.R.; Karthik, C.; Kadirvelu, K.; Arulselvi, P.I.; Shanmugasundaram, T.; Bruno, B.; Rajkumar, M. Understanding the molecular mechanisms for the enhanced phytoremediation of heavy metals through plant growth promoting rhizobacteria: A review. J. Environ. Manag. 2020, 254, 1–14. [Google Scholar] [CrossRef]
- Wani, S.P.; Gopalakrishnan, S. Plant Growth-Promoting Microbes for Sustainable Agriculture. In Plant Growth Promoting Rhizobacteria (PGPR): Prospects for Sustainable Agriculture; Sayyed, R., Reddy, M., Antonius, S., Eds.; Springer: Singapore, 2019. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.; Tabassum, B.; Fathi Abd_Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.; Sahoo, R.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef]
- Ateş, O.; Kivan, M. Effects of Arthrobacter arilaitensis and Pseudomonas putida on salt stress tolerance in wheat. Environ. Eng. Manag. J. 2021, 20, 2025–2032. [Google Scholar] [CrossRef]
- Domanska, M.; Kamińska, J. Quantification of proteobacteria with fluorescence in situ hybridization and next-generation sequencing. Environ. Eng. Manag. J. 2022, 21, 981–993. [Google Scholar] [CrossRef]
- Sumbul, A.; Ali Ansari, R.; Rizvi, R.; Mahmood, I. Azotobacter: A potential bio-fertilizer for soil and plant health management. Saudi, J. Biol. Sci. 2020, 27, 3634–3640. [Google Scholar] [CrossRef]
- Xie, L.; Lehvävirta, S.; Timonen, S.; Kasurinen, J.; Niemikapee, J.; Valkonen, J.P.T. Species-Specific Synergistic effects of two plant growth-promoting microbes on green roof plant biomass and photosynthetic efficiency. PLoS ONE 2018, 13, 0209432. [Google Scholar] [CrossRef] [PubMed]
- David, B.V.; Chandrasehar, G.; Selvam, P.N. Pseudomonas fluorescens: A plant- growth-promoting rhizobacterium (PGPR) with potential role in biocontrol of pests of crops. In Crop Improvement through Microbial Biotechnology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 221–243. [Google Scholar] [CrossRef]
- Goswami, M.; Suresh Deka, S. Plant growth-promoting rhizobacteria—Alleviators of abiotic stresses in soil: A review. Pedosphere 2020, 30, 40–61. [Google Scholar] [CrossRef]
- Botelho, G.R.; Mendonça-Hagler, L.C. Fluorescent Pseudomonads associated with the rhizosphere of crops: An overview. Braz. J. Microbiol. 2006, 37, 401–416. [Google Scholar] [CrossRef] [Green Version]
- Prajapati, K.; Yami, K.D.; Singh, A. Plant growth promotional effect of Azotobacter chroococcum, Piriformospora indica and vermicompost on rice plant. Nepal J. Sci. Technol. 2010, 9, 3170. [Google Scholar] [CrossRef] [Green Version]
- Gavrilescu, M.; Diaconu, M.; Bulgariu, L.; Volf, I.; Catrinescu, C.; Cozma, P.; Hlihor, R.-M.; Ghinea, C.; Apostol, L.C.; Comăniță, E.-D.; et al. Exploring and Exploiting Microbial and Plant Abilities and Interactions in Environmental Bioremediation; Perfomantica Publishing House: Iași, Romania, 2019. (In Romanian) [Google Scholar]
- Khalid, A.H.; Jin, H.J. Heavy metal resistance of bacteria and its impact on the production of antioxidant enzymes. Afr. J. Microbiol. Res. 2013, 7, 2288–2296. [Google Scholar] [CrossRef]
- Samuel, A.D.; Oneț, A.; Dincă, L.; Enescu, R.; Deleanu, E.; Oneț, C.; Stanciu, A. Enzymatic indicators of soil quality and nutrients content in the forest soils from Romania. Environ. Eng. Manag. J. 2022, 21, 1245–1253. [Google Scholar]
- Liu, W.; Yang, C.; Shi, S.; Shu, W. Effects of plant growth-promoting bacteria isolated from copper tailings on plants in sterilized and non-sterilized tailings. Chemosphere 2013, 97, 47–53. [Google Scholar] [CrossRef]
- Wang, Y.; Narayanan, M.; Shi, X.; Chen, X.; Li, Z.; Natarajan, D.; Ma, Y. Plant growth-promoting bacteria in metal-contaminated soil: Current perspectives on remediation mechanisms. Front. Microbiol. 2022, 13, 966226. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Nagendran, R.; Lee, K.J.; Lee, W.H.; Kim, S.Z. Influence of plant growth promoting bacteria and Cr6+ on the growth of Indian mustard. Chemospere 2006, 62, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Freitas, H. Effects of inoculation of plant-growth promoting bacteria on Ni uptake by Indian mustard. Bioresour. Technol. 2008, 99, 3491–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandlic, C.J.; Mendez, M.O.; Chorover, J.; Machado, B.; Maier, R.M. Plant growth-promoting bacteria for phytostabilization of mine tailings. Environ. Sci. Technol. 2008, 42, 2079–2084. [Google Scholar] [CrossRef]
- Grandlic, C.J.; Palmer, M.W.; Maier, R.M. Optimization of plant growth- promoting bacteria-assisted phytostabilization of mine tailings. Soil Biol. Biochem. 2009, 41, 1734–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreiro, H.; Pereira, S.I.A.; Marques, A.P.G.C.; Rangel, A.O.S.S.; Castro, P.M.R. Effects of soil sterilization and metal spiking in plant growth promoting rhizobacteria selection for phytotechnology purposes. Geoderma 2019, 334, 72–81. [Google Scholar] [CrossRef]
- Li, K.; DiLegge, M.J.; Minas, I.S.; Hamm, A.; Manter, D.; Vivanco, J. Soil sterilization leads to re-colonization of a healthier rhizosphere microbiome. Rhizosphere 2019, 12, 100176. [Google Scholar] [CrossRef]
- Stefanescu, I.A. Impact of Bacillus megaterium on fertilization with phosphogypsum. Int. J. Eng. Res. Technol. 2011, 17, 93–97. [Google Scholar]
- Mourato, M.; Moreira, I.; Leitão, I.; Pinto, F.; Sales, J.; Martins, L. Effect of heavy metals in plants of the genus Brassica. Int. J. Mol. Sci. 2015, 16, 17975–17998. [Google Scholar] [CrossRef] [Green Version]
- Drozdova, I.; Alekseeva-Popova, N.; Dorofeyev, V.; Bech, J.; Belyaeva, A.; Roca, N.A. Comparative study of the accumulation of trace elements in Brassicaceae plant species with phytoremediation potential. Appl. Geochem. 2019, 108, 104377. [Google Scholar] [CrossRef]
- Kuramshima, Z.M.; Smirnova, Y.V.; Khairullin, R.M. Cadmium and nickel toxicity for Sinapis alba plants inoculated with endophytic strains of Bacillus subtilis. Russ. J. Plant Physiol. 2018, 65, 269–277. [Google Scholar] [CrossRef]
- Berková, V.; Berka, M.; Griga, M.; Kopecká, R.; Prokopová, M.; Luklová, M.; Horáček, J.; Smýkalová, I.; Čičmanec, P.; Novák, J.; et al. Molecular mechanisms underlying flax (Linum usitatissimum L.) tolerance to cadmium: A case study of proteome and metabolome of four different flax genotypes. Plants 2022, 11, 2931. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.H.; Ali, S.; Hussain, S.; Kamran, M.; Chattha, M.S.; Ahmad, S.; Aqeel, M.; Rizwan, M.; Aljarba, N.H.; Alkahtani, S.; et al. Flax (Linum usitatissimum L.): A potential candidate for phytoremediation? Biological and economical points of view. Plants 2020, 9, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiloğlu, S.; Adiloğlu, S.A.; Açıkgöz, F.E.; Yeniaras, T.; Solmaz, Y. Phytoremediation of cadmium from soil using patience dock (Rumex patientia L.). Anal Lett. 2016, 49, 601–606. [Google Scholar] [CrossRef]
- Adiloğlu, S. Interaction of Some Heavy Metals with copper content in dock plant, KSU. J. Agric. Nat. 2020, 23, 1078–1084. [Google Scholar] [CrossRef]
- Alsherif, E.A.; Al-Shaikh, T.M.; AbdElgawad, H. Heavy metal effects on biodiversity and stress responses of plants inhabiting contaminated soil in Khulais, Saudi Arabi. Biology 2022, 11, 164. [Google Scholar] [CrossRef]
- Khoramnejadian, S.; Saeb, K. Accumulation and translocation of heavy metals by Amaranthus retroflexus. J. Environ. Health Sci. 2015, 1, 58–60. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, D.; Yuan, Y.; Chen, P.; Zhang, P.; Jin, F.; Yang, Q.; Feng, B. A promising crop for cadmium-contamination remediation: Broomcorn millet, Ecotoxicol. Environ. Saf. 2021, 224, 112669. [Google Scholar] [CrossRef]
- Toroni, A.O.; Aguoru, C.U.; Ogbonna, I.O.; Olasan, J.O. Comparative studies of heavy metals and mineral residues in some farm crops around mining community of ribi, awe local government area of Nasarawa state. Int. J. Environ. Agric. Biotech. 2019, 4, 28. [Google Scholar] [CrossRef] [Green Version]
- Mora, Y.; Díaz, R.; Vargas-Lagunas, C.; Peralta, H.; Guerrero, G.; Aguilar, A.; Encarnación, S.; Girard, L.; Mora, J. Nitrogen-fixing rhizobial strains isolated from common bean seeds: Phylogeny, physiology, and genome analysis. Appl. Environ. Microbiol. 2014, 80, 5644–5654. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Yang, F.; Wang, S.; Ma, H.; Liang, J.; Chen, Y. Co-existence of Rhizobia and Diverse Non-rhizobial Bacteria in the Rhizosphere and Nodules of Dalbergia odorifera Seedlings Inoculated with Bradyrhizobium elkanii, Rhizobium multihospitium–Like and Burkholderia pyrrocinia–Like Strains. Front. Microbiol. 2017, 8, 2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-López, A.; Rogel, M.A.; Ormeño-Orrillo, E.; Martínez-Romero, J.; Martínez-Romero, E. Phaseolus vulgaris seed-borne endophytic community with novel bacterial species such as Rhizobium endophyticum sp. nov. Syst. App. Microbiol. 2010, 33, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.B.; Costa, L.E.; Vanetti, M.C.; de Araújo, E.F.; de Queiroz, M.V. Endophytic bacteria isolated from common bean (Phaseolus vulgaris) exhibiting high variability showed antimicrobial activity and quorum sensing inhibition. Curr. Microbiol. 2015, 71, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, X.; Chen, W.; Huang, Q. Isolation and Identification of three potassium-solubilizing bacteria from rape rhizospheric soil and their effects on ryegrass. Geomicrobiol. J. 2017, 34, 873–880. [Google Scholar] [CrossRef]
- Diaconu, M. Ecotoxicological Methods and Tests; Performantica Publishing House: Bucharest, Romania, 2016; ISBN 9786066854177. (In Romanian) [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Gordon, S.A.; Robert, P.W. Colorimetric Estimation of indoleacetic Acid. Plant Physiology 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.K.; Cho, H.S.; Moon, Y.C.; Ban, S.J.; Kim, J.Y. Cadmium and lead uptake capacity of energy crops and distribution of metals within the plant structures. KSCE J. Civ. Eng. 2013, 17, 44–50. [Google Scholar] [CrossRef]
- Lally, R.D.; Galbally, P.; Moreira, A.S.; Spink, J.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Application of endophytic Pseudomonas fluorescens and a bacterial consortium to Brassica napus can increase plant height and biomass under greenhouse and field conditions. Front. Plant Sci. 2017, 8, 2193. [Google Scholar] [CrossRef] [Green Version]
- Axinte, M.; Borcean, I.; Roman, G.V.; Muntean, M.S. Phytotechnics, 4th ed.; Ion Ionescu de la Brad’ Publishing House: Iasi, Romania, 2006. (In Romanian) [Google Scholar]
- CABI–Invasive Species Compendium. Available online: www.cabi.org/isc (accessed on 30 September 2022).
- Habiyaremye, C.; Matanguihan, J.B.; D’Alpoim Guedes, J.; Ganjyal, G.M.; Whiteman, M.R.; Kidwell, K.K.; Murphy, K.M. Proso Millet (Panicum miliaceum L.) and its potential for cultivation in the pacific northwest, U.S.: A review. Front. Plant Sci. 2017, 7, 1961. [Google Scholar] [CrossRef] [Green Version]
- Jnawali, A.D.; Ojha, R.B.; Marahatta, S. Role of Azotobacter in soil fertility and sustainability—A review. Adv. Plants Agric. Res. 2015, 2, 250–253. [Google Scholar] [CrossRef]
- Romero-Perdomo, F.; Abril, J.; Camelo, M.; Moreno-Galván, A.; Pastrana, I.; RojasTapias, D.; Bonilla, R. Azotobacter chroococcum as a potentially useful bacterial biofertilizer for cotton (Gossypium hirsutum): Effect in reducing N fertilization. Rev. Argent. Microbiol. 2017, 49, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Sivasakthi, S.; Usharani, G.; Saranjar, P. Biocontrol potentiality of plant growth promoting bacteria (PGPR)—Pseudomonas fluorescens and Bacillus subtilis: A review. Afr. J. Agric. 2014, 9, 1265–1277. [Google Scholar] [CrossRef]
- Muis, A. Biomass production and formulation of Bacillus subtilis for biological control. Indones. J. Agric. Sci. 2006, 7, 51–56. [Google Scholar] [CrossRef]
- Adam, M.; Heuer, H.; Hallmann, J. Bacterial antagonists of fungal pathogens also control root-not nematodes by induced systemic resistance of tomato plants. PLoS ONE 2014, 9, 90402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndeddy Aka, R.J.; Babalola, O.O. Effect of bacterial inoculation of strains of Pseudomonas aeruginosa, Alcaligenes feacalis and Bacillus subtilis on germination, growth and heavy metal (Cd, Cr, and Ni) uptake of Brassica juncea. Int. J. Phytoremediation 2015, 18, 200–209. [Google Scholar] [CrossRef]
- Sarwar, S.; Khaliq, A.; Yousra, M.; Sultan, T.; Ahmad, N.; Khan, M.Z. Screening of siderophore-producing PGPRs isolated from groundnut (Arachis hypogaea L.) rhizosphere and their influence on iron release in soil. Commun. Soil Sci. Plant Anal. 2020, 51, 1680–1692. [Google Scholar] [CrossRef]
- Wani, S.A.; Chand, S.; Tahir Ali, T. Potential use of Azotobacter chroococcum in crop production: An overview. Curr. Agric. Res. J. 2013, 1, 35–38. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar Behl, R.; Narula, N. Establishment of phosphate-solubilizing strains of Azotobacter chroococcum in the rhizosphere and their effect on wheat cultivars under greenhouse conditions. Microbiol. Res. 2001, 15, 87–93. [Google Scholar] [CrossRef]
- Príncipe, A.; Fernandez, M.; Torasso, M.; Godino, A.; Fischer, S. Effectiveness of Tailocins 791 produced by Pseudomonas fluorescens SF4c in controlling the bacterial-spot disease in 792 tomatoes caused by Xanthomonas vesicatoria. Microbiol. Res. 2018, 212, 94–100. [Google Scholar] [CrossRef]
- Kundan, R.; Pant, G.; Jadon, N.; Agrawal, P.K. Plant growth promoting rhizobacteria: Mechanism and current prospective. J. Fertil. Pestic. 2015, 6, 1000155. [Google Scholar] [CrossRef]
- Loper, J.E.; Gross, H. Genomic analysis of antifungal metabolite production by Pseudomonas fluorescens Pf-5. Eur. J. Plant Pathol. 2007, 119, 265–278. [Google Scholar] [CrossRef]
- Althaf, H.S.; Srinivas, P. Evaluation of plant growth promoting traits by Pseudomonas and Azotobacter isolated from rhizotic soils of two selected agro forestry tree species of Godavari Belt Region, India. Asian J. Exp. Biol. Sci. 2013, 4, 431–436. [Google Scholar]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Dietz, K.J. Characterization of plant growth promoting rhizobacteria isolated from 533 polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2001, 47, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Ashnaei, S.P. Plant growth promoting rhizobacteria and Rhizophagus irregularis: Biocontrol of rice blast in wild type and mycorrhiza-defective mutant. J. Plant Prot. Res 2019, 59, 362–375. [Google Scholar] [CrossRef]
- Bhatia, S.; Dubey, R.; Maheshwari, D. Enhancement of plant growth and suppression of collar rot of sunflower caused by Sclerotium Rolfsii through fluorescent Pseudomonas. Indian Phytopathol. 2005, 58, 17–24. [Google Scholar]
- Egamberdieva, D. Growth response of wheat cultivars to bacterial inoculation in calcareous. Plant Soil Environ. 2010, 56, 570–573. [Google Scholar] [CrossRef] [Green Version]
- Chauchan, S.; Wadhwa, K.; Vasudeva, M.; Narula, N. Potential of Azotobacter spp. as biocontrol agents against Rhizoctonia solani and Fusarium oxysporumin cotton (Gossypium hirsutum), guar (Cyamopsis tetragonoloba) and tomato (Lycopersicum esculentum). Arch. Agron. Soil Sci. 2012, 58, 1365–1385. [Google Scholar] [CrossRef]
- Biari, A.; Gholami, A.; Rahmani, H.A. Growth promotion and enhanced nutrient uptake of maize (Zea mays L.) by application of plant growth promoting rhizobacteria in arid region of Iran. J. Biol. Sci. 2008, 8, 1015–1020. [Google Scholar] [CrossRef] [Green Version]
- Gholami, A.; Shahsavani, S.; Nezarat, S. The effect of plant growth promoting rhizobacteria (PGPR) on germination, seedling growth and yield of maize. Eng. Technol. 2008, 49, 1–8. [Google Scholar]
- Burd, G.I.; Dixon, D.G.; Glick, B.R. Plant growth promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol. 2000, 46, 237–245. [Google Scholar] [CrossRef]
- Nezarat, S.; Gholami, A. Screening plant growth promoting rhizobacteria for improving seed germination, seedling growth and yield of maize. Pak. J. Biol. Sci. 2009, 12, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A. Efficacy of phosphate solubilizing bacteria isolated from vertisols on growth and yield parameters of sorghum. Res. J. Microbiol. 2007, 2, 550–559. [Google Scholar]
- Agbodjato, N.A.; Noumavo, P.A.; Baba-Moussa, F.; Salami, H.A.; Sina, H.; Sèzan, A.; Bankolé, H.; Adjanohoun, A.; Baba-Moussa, L. Characterization of potential plant growth promoting rhizobacteria isolated from maize (Zea mays L.) in central and northern Benin (West Africa). Appl. Environ. Soil Sci. 2015, 2015, 901656. [Google Scholar] [CrossRef] [Green Version]
- Poonguzhali, S.; Madhaiyan, M.; Sa, T. Isolation and identification of phosphate solubilizing bacteria from chinese cabbage and their effect on growth and phosphorus utilization of plants. J. Microbiol. Biotechnol. 2008, 18, 773–777. [Google Scholar] [PubMed]
- Verma, A.; Kukreja, K.; Pathak, D.V.; Suneja, S.; Narula, N. In vitro production of plant growth regulators (PGRs) by Azotobacter chroococcum. Indian J. Microbiol. 2001, 41, 305–307. [Google Scholar]
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis Strain SJ-101 as a bioinoculant for concurrent plant growth promotion and nickel accumulation in Brassica juncea. Chemosphere 2006, 64, 991–997. [Google Scholar] [CrossRef]
- Ravikumar, S.; Kathiresan, K.; Ignatiammal, S.T.M.; Babu Selvam, M.; Shanthy, S. Nitrogen-fixing azotobacters from mangrove habitat and their utility as marine biofertilizers. J. Exp. Mar. Biol. 2004, 312, 5–17. [Google Scholar] [CrossRef]
- Cazorla, F.M.; Romero, D.; Pérez-García, A.; Lugtenberg, B.J.J.; de Vicente, A.; Bloemberg, G. Isolation and characterization of antagonistic Bacillus subtilis strains from the avocado rhizoplane displaying biocontrol activity. J. Appl. Microbiol. 2007, 103, 1950–1959. [Google Scholar] [CrossRef]
- Canbolat, M.Y.; Bilen, S.; Çakmakç, R.; Şahin, F.; Aydın, A. Effect of plant growth-promoting bacteria and soil compaction on barley seedling growth, nutrient uptake, soil properties and rhizosphere microflora. Biol. Fertil. Soils 2006, 42, 350–357. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Samiyappan, R. ACC Deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J. Appl. Microbiol. 2007, 102, 1283–1292. [Google Scholar] [CrossRef]
- Shaharoona, B.; Naveed, M.; Arshad, M.; Zahir, Z.A. Fertilizer-dependent efficiency of Pseudomonas for improving growth, yield, and nutrient use efficiency of wheat (Triticum aestivum L.). Appl. Microbiol. Biotechnol. 2008, 79, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Charles, T.C.; Glic, B.R. Isolation and characterization of new plant growth-promoting bacterial endophytes. Appl. Soil Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Plants | Beneficial Effects | Ref. |
|---|---|---|---|
| Bacillus polymyxa, Bacillus pantothenticus, Bacillus anthracis, Bacillus thuringiensis, Bacillus circulans, Pseudomonas cichorii, Pseudomonas putida, Pseudomonas syringae | Zea mays L. | Improved seed germination and plant growth | [82] |
| Pseudomonas poae, Pseudomonas trivali | Brassica campestris L. spp. Pekinensis | Stimulated plant roots | [83] |
| Azotobacter chroococcum | Brassica juncea L. | Increased seed germination | [84] |
| Bacillus subtilis strain SJ-101 | Brassica juncea L. | Increased plant growth | [85] |
| Azotobacter chroococcum, Azotobacter virelandii and Azotobacter beijerinckii | Rhizophora mangle | Increased the roots, shoots, leaves of plants and chlorophyll content | [86] |
| Azotobacter chroococcum | Triticum aestivum L. | Increased the plant biomass, the length of wheat and the wheat yield | [67] |
| Bacillus subtilis PCL1608 Bacillus subtilis PCL1612 | Persea americana mill, Solanum lycopersicum | Produced antifungal lipopeptides and good colonization abilities | [87] |
| Bacillus RC01, Bacillus RC02, Bacillus RC03, Bacillus M-13 | Hordeum vulgare L. | Increased the root and shoot biomass | [88] |
| Pseudomonas fluorescens | Arachis hypogea L. | Induced systemic resistance, antifungal activity | [89] |
| Pseudomonas fluorescens ACC5 Pseudomonas fluorescens biotype F (ACC73) | Triticum aestivum L. | Increased root weight, grain yield, number of tillers per plant and straw yield | [90] |
| Bacillus subtilis KP717559 | Brassica juncea | Increased root and shoot length and the content of fresh and dry biomass | [64] |
| Pseudomonas brassicacearum Am3, Pseudomonas putida Bm3, Pseudomonas marginalis D | Brassica napus | More vigorous plant | [72] |
| Pseudomonas fluorescens YsS6 | Brassica rapa | Promoted root elongation Plants much healthier and higher | [91] |
| Azotobacter sp. | Sinapis alba L. | Increased the root, shoot and leaf length and the root, shoot and leaf dry weight | This study |
| Azotobacter sp. | Brassica napus L. | Increased the root, shoot and leaf length and the root, shoot and leaf dry weight | This study |
| Azotobacter sp. | Amaranthus retroflexus L. | Did not improve the growth | This study |
| Azotobacter sp. | Panicum miliaceum L. | Increased the shoot and leaf length and dry weight Vigorous shoots | This study |
| Azotobacter sp. | Rumex patientia L. | Increased the root length and the root, shoot and leaf dry weight Stimulated root branching | This study |
| Azotobacter sp. | Linum usitatissimum L. | Increased the root, shoots and leaf length and the shoot and leaf dry weight Stimulated root branching | This study |
| Bacillus sp. | Sinapis alba L. | Increased the root, shoot and leaf length | This study |
| Bacillus sp. | Amaranthus retroflexus L. | Increased the root, shoot and leaf length | This study |
| Bacillus sp. | Brassica napus L. | Increased the root, shoot and leaf length and their dry weight biomass | This study |
| Bacillus sp. | Panicum miliaceum L. | Increased the shoot and leaf length and the root dry weight Vigorous shoots | This study |
| Bacillus sp. | Rumex patientia L. | Increased the root length and dry weight Stimulated root branching | This study |
| Bacillus sp. | Linum usitatissimum L. | Increased the root shoot and leaf length Branched roots | This study |
| Pseudomonas sp. | Sinapis alba L. | Increased the root, shoot and leaf length Increased the root, shoot and leaf dry weight | This study |
| Pseudomonas sp. | Brassica napus L. | Increased the root, shoot and leaf length Increased the root, shoot and leaf dry weight Stimulated root branching | This study |
| Pseudomonas sp. | Amaranthus retroflexus L. | Increased the shoots + leaves length | This study |
| Pseudomonas sp. | Panicum miliaceum L. | Increased the shoots + leaves length and dry weight | This study |
| Pseudomonas sp. | Rumex patientia L. | Increased the root, shoot and leaf length and dry weight Stimulated root branching Vigorous shoots | This study |
| Pseudomonas sp. | Linum usitatissimum L. | Increased the root, shoot and leaf length Stimulated root branching Vigorous shoots | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minuț, M.; Diaconu, M.; Roșca, M.; Cozma, P.; Bulgariu, L.; Gavrilescu, M. Screening of Azotobacter, Bacillus and Pseudomonas Species as Plant Growth-Promoting Bacteria. Processes 2023, 11, 80. https://doi.org/10.3390/pr11010080
Minuț M, Diaconu M, Roșca M, Cozma P, Bulgariu L, Gavrilescu M. Screening of Azotobacter, Bacillus and Pseudomonas Species as Plant Growth-Promoting Bacteria. Processes. 2023; 11(1):80. https://doi.org/10.3390/pr11010080
Chicago/Turabian StyleMinuț, Mariana, Mariana Diaconu, Mihaela Roșca, Petronela Cozma, Laura Bulgariu, and Maria Gavrilescu. 2023. "Screening of Azotobacter, Bacillus and Pseudomonas Species as Plant Growth-Promoting Bacteria" Processes 11, no. 1: 80. https://doi.org/10.3390/pr11010080
APA StyleMinuț, M., Diaconu, M., Roșca, M., Cozma, P., Bulgariu, L., & Gavrilescu, M. (2023). Screening of Azotobacter, Bacillus and Pseudomonas Species as Plant Growth-Promoting Bacteria. Processes, 11(1), 80. https://doi.org/10.3390/pr11010080