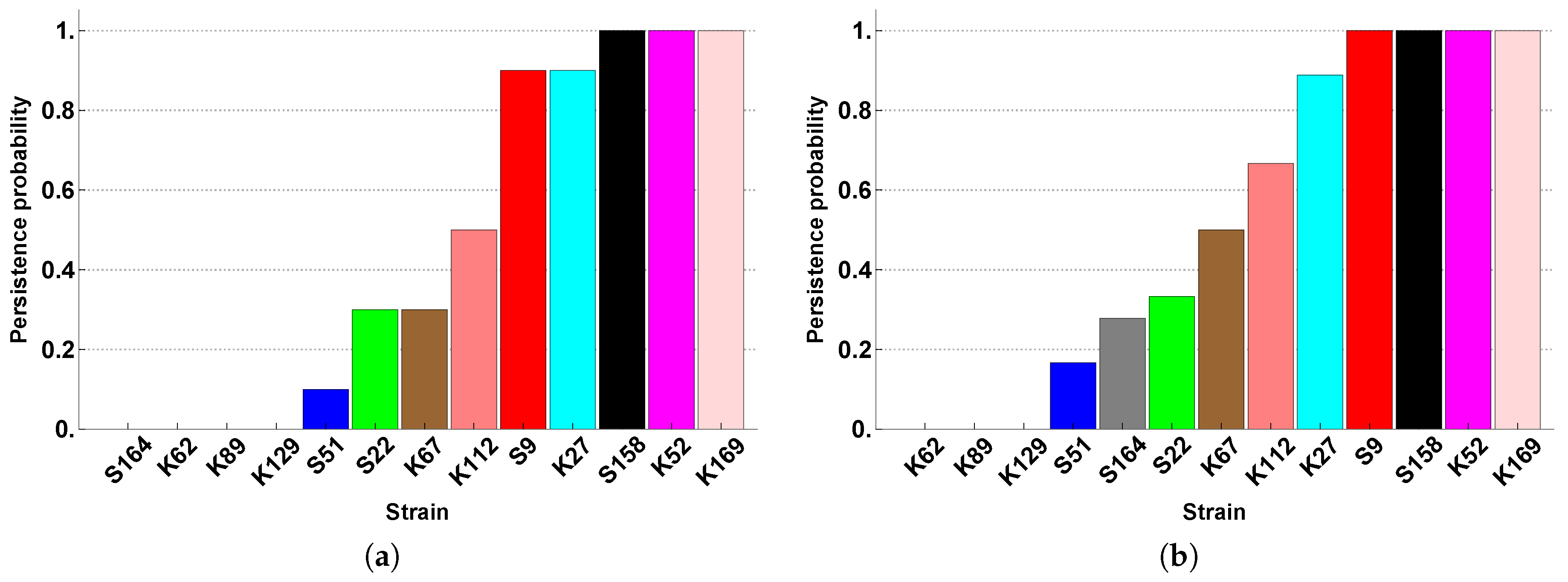
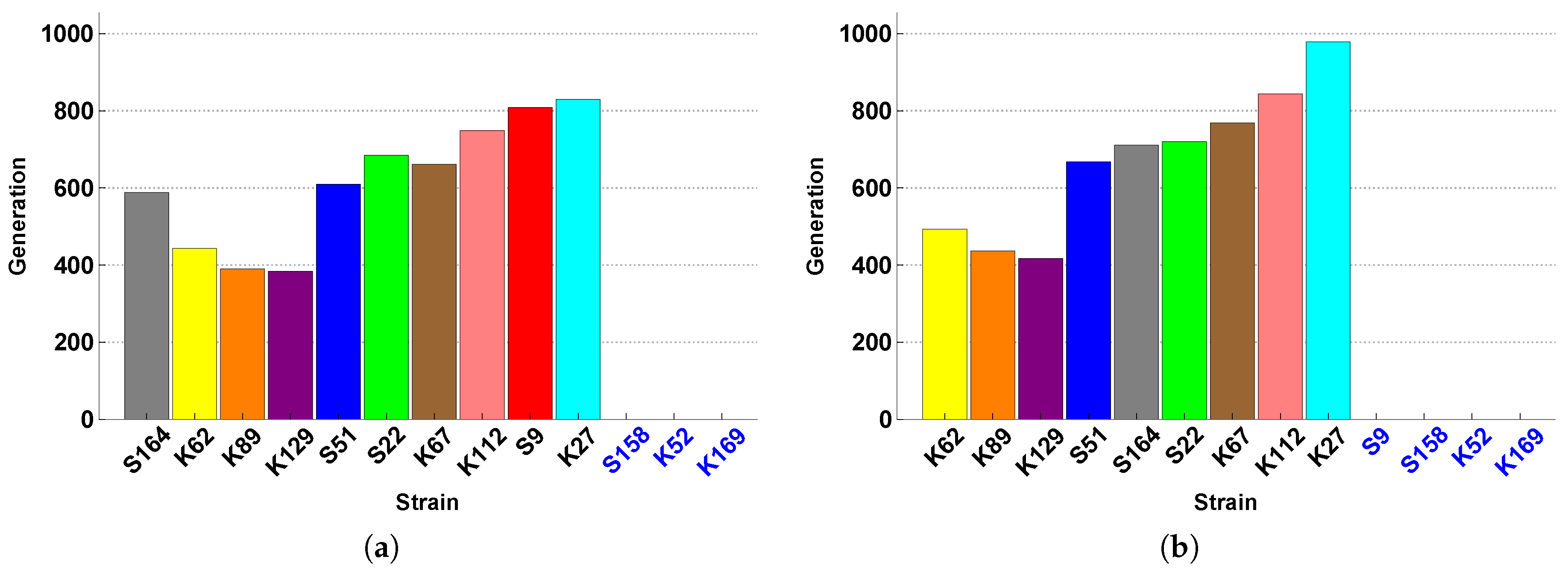
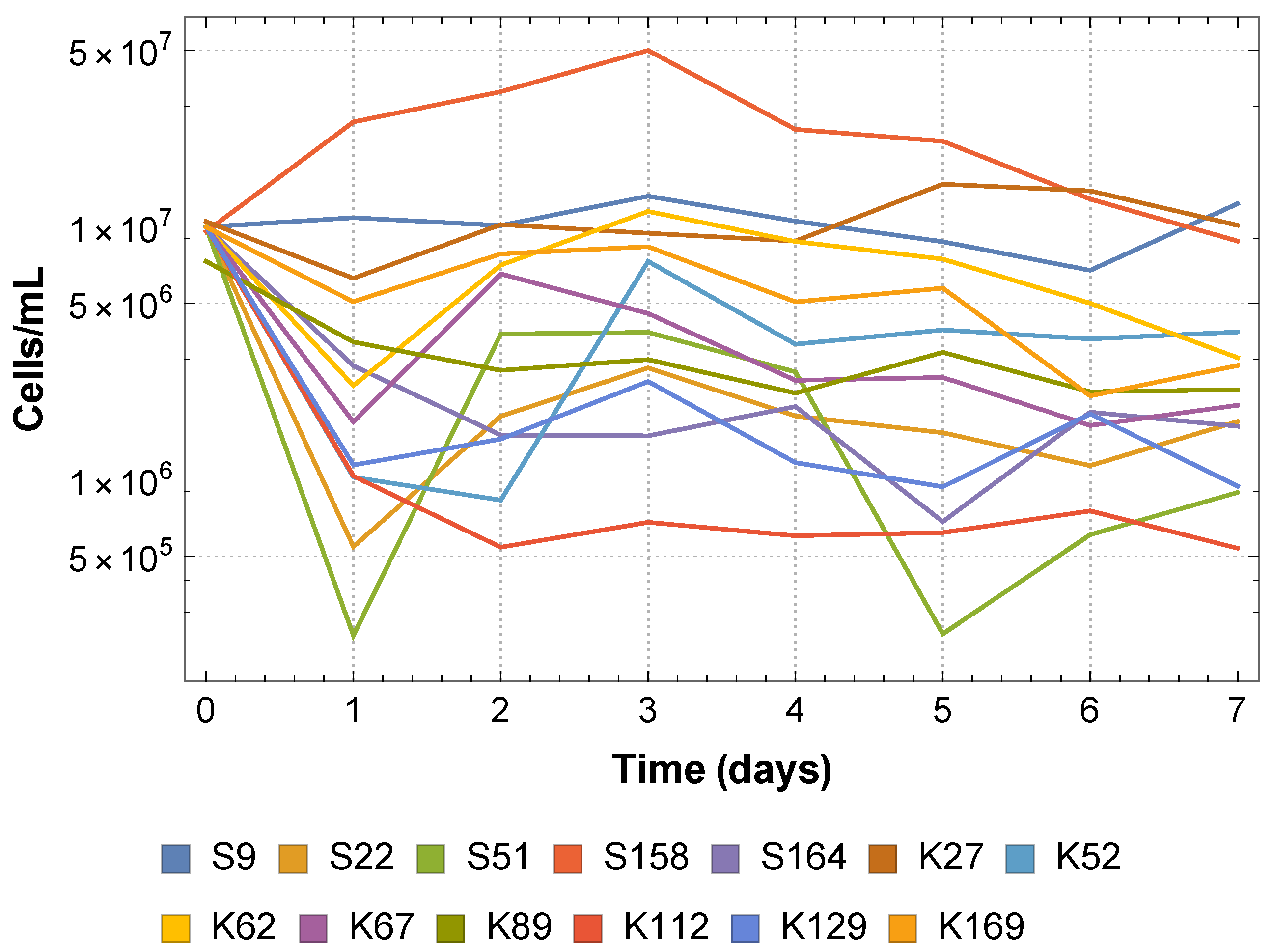
The experiments of Vandermaesen et al. [
11] focused on bioaugmentation success and therefore collected data related to BAM mineralization and MSH1 survival. The data related to the SFIs themselves are their monoculture growth curves and their monoculture survival curves on acetate (see
Appendix A). These data can give us an idea of how the SFIs grow and persist in isolation, and on this basis Vandermaesen et al. [
11] classified the strains according to their “intrinsic competitiveness”, a classification that we can use as an additional feature of the strains. However, these data do not give us any information about how the SFIs may interact, and in particular compete, when they are inoculated together in co-culture.
The information we do have regarding the interactions between SFIs is indirect. From the differences in mineralization parameters between the different co-culture combinations, we can infer when there are interaction effects occurring between strains, by comparing the mineralization performances of MSH1 alone, in co-culture with individual SFIs, and in co-culture with both strains. The mineralization performance was studied using the Gompertz model (see
Section 2.2 for details). This model has four parameters: the lag time
, the maximum mineralization rate
, the total extent of mineralization
A, and the endogenous mineralization rate
c.
To study the strain interaction effects, we focus on two of these parameters: the lag time
and the maximum mineralization rate
. These two parameters have been highlighted as key to the success of bioaugmentation strategies and are more strongly linked with both positive and negative mineralization effects than the other mineralization parameters [
25].
3.1.1. Identifying Strain Identity Effects
Since each synthetic community included MSH1, the total richness of a community
is given by
=
+ 1, where
is the number of SFIs present. In addition to all combinations of individual SFIs with MSH1 (13 combinations at
= 1), all 78 different pair combinations of two SFIs with MSH1 (
= 2) were tested (further details given in
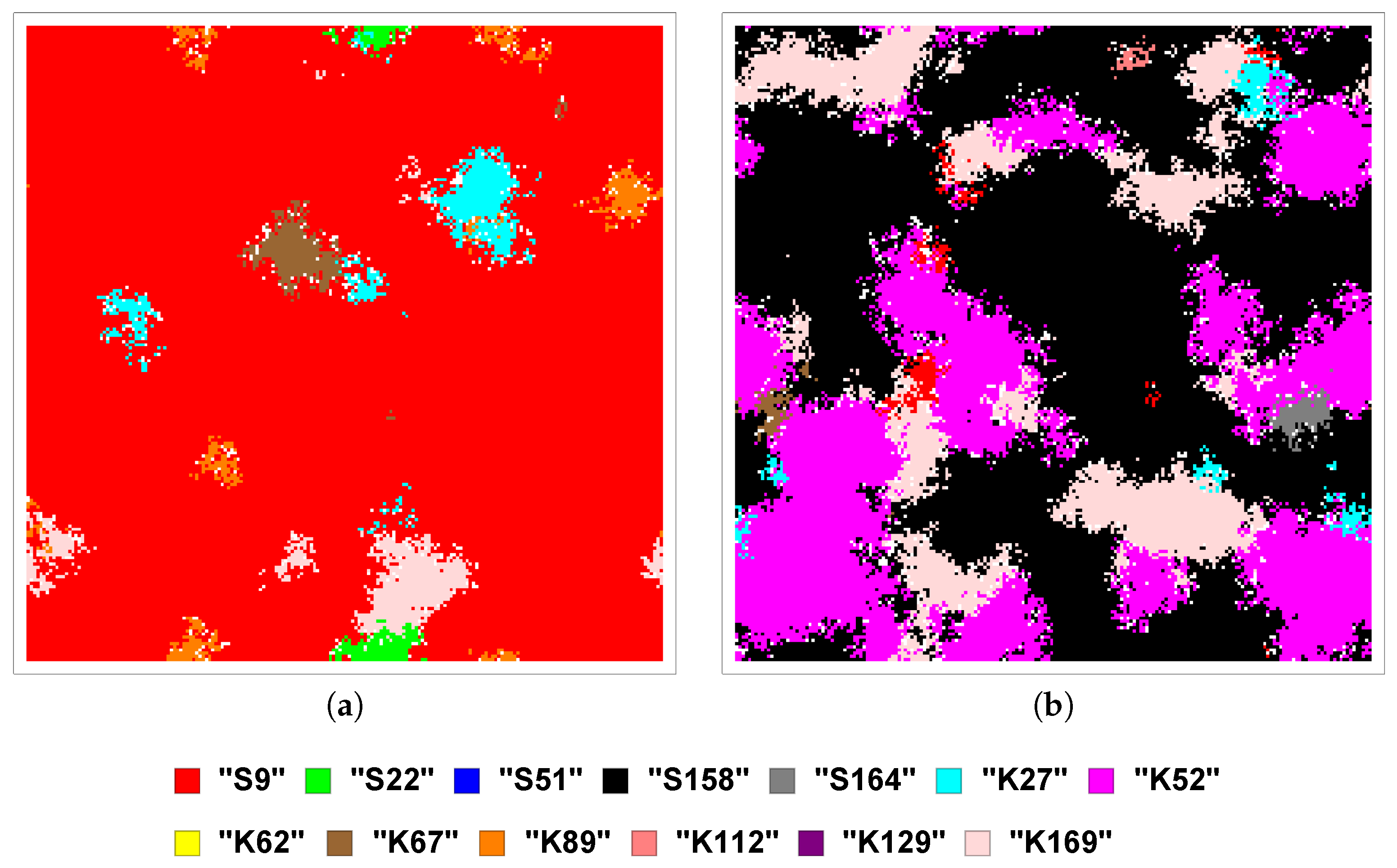
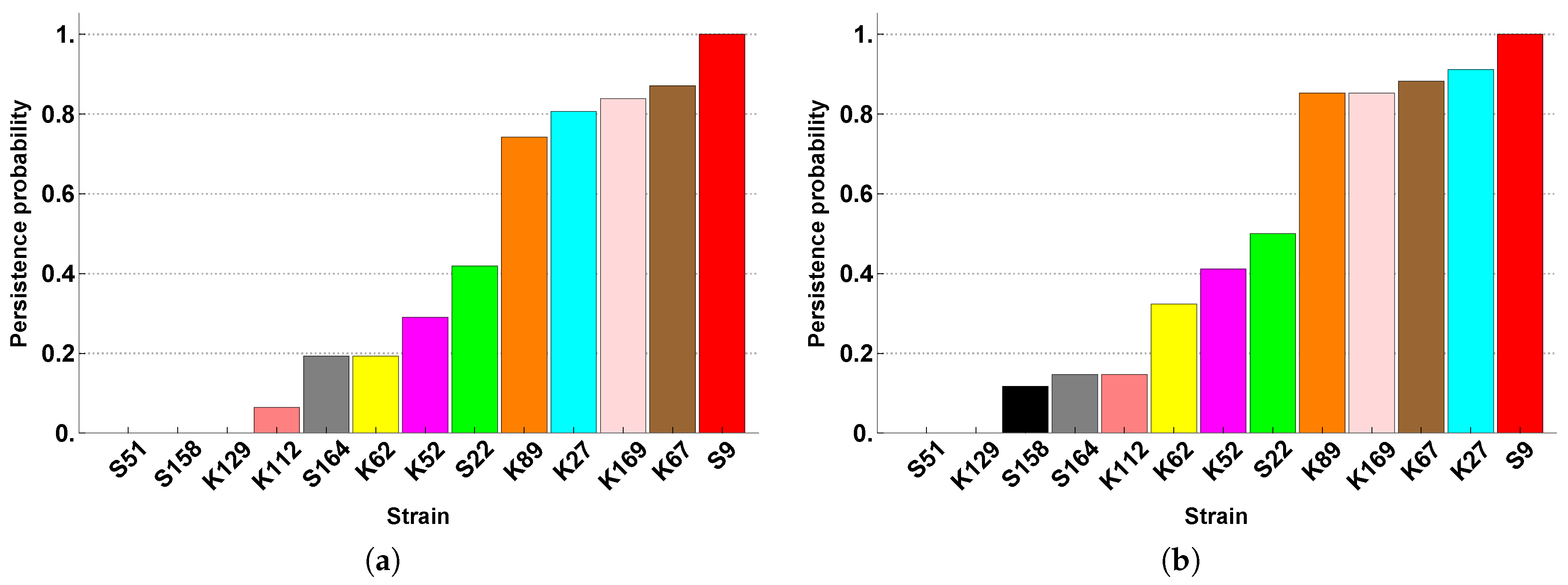
Section 2). The 13 SFIs are assigned the following labels: S9, S22, S51, S158, S164, K27, K52, K62, K67, K89, K112, K129, and K169.
Previous studies have also used growth model parameters to identify different growth behaviours between microbial species, for example through the use of regression models [
26]. We employ a statistical test known as the pairwise Tukey test [
27] to compare values of the lag time
and mineralization rate
across different
levels. With this test it is possible to evaluate whether values of
or
observed for a specific synthetic community are significantly different from the respective parameter values observed for a different community.
The Tukey test statistic is
, where
and
are the respective means of the observations of two populations being compared, and
is the data’s standard error [
28]. The null hypothesis of the test is that the means are from the same population. The test statistic is then compared to a critical test statistic value
which is obtained from the studentized range distribution [
29]. If
z is larger than
, then the null hypothesis is rejected and it is concluded that the two populations are significantly different. Tests were performed at the 95% significance level, using Mathematica (version 11.0, Wolfram Research, Champaign, IL, USA).
Two types of tests were conducted. First, we compared values of or for communities against (i.e., MSH1 alone) as a benchmark population. To determine the sign of the change, we consider the biological interpretation of a positive or beneficial change in these parameters. For the lag time , a decrease in this parameter is considered a positive effect while an increase is considered a negative effect. For the mineralization rate , the opposite is true.
The second type of test required selecting one of the SFIs as the focal strain. The test then compared values of or for the communities including this focal strain, against the values of or for the corresponding community for the non-focal strain. For example, when S9 was the focal strain of the test and the parameter under consideration was , we selected all communities containing S9. One such community contained S9, S22 and MSH1. We then compared the values of of this community against the values of of the community containing S22 and MSH1. This allowed us to conclude if in this case there were significant differences in lag time due to the inclusion of S9. This analysis was repeated for every strain other than the focal strain.
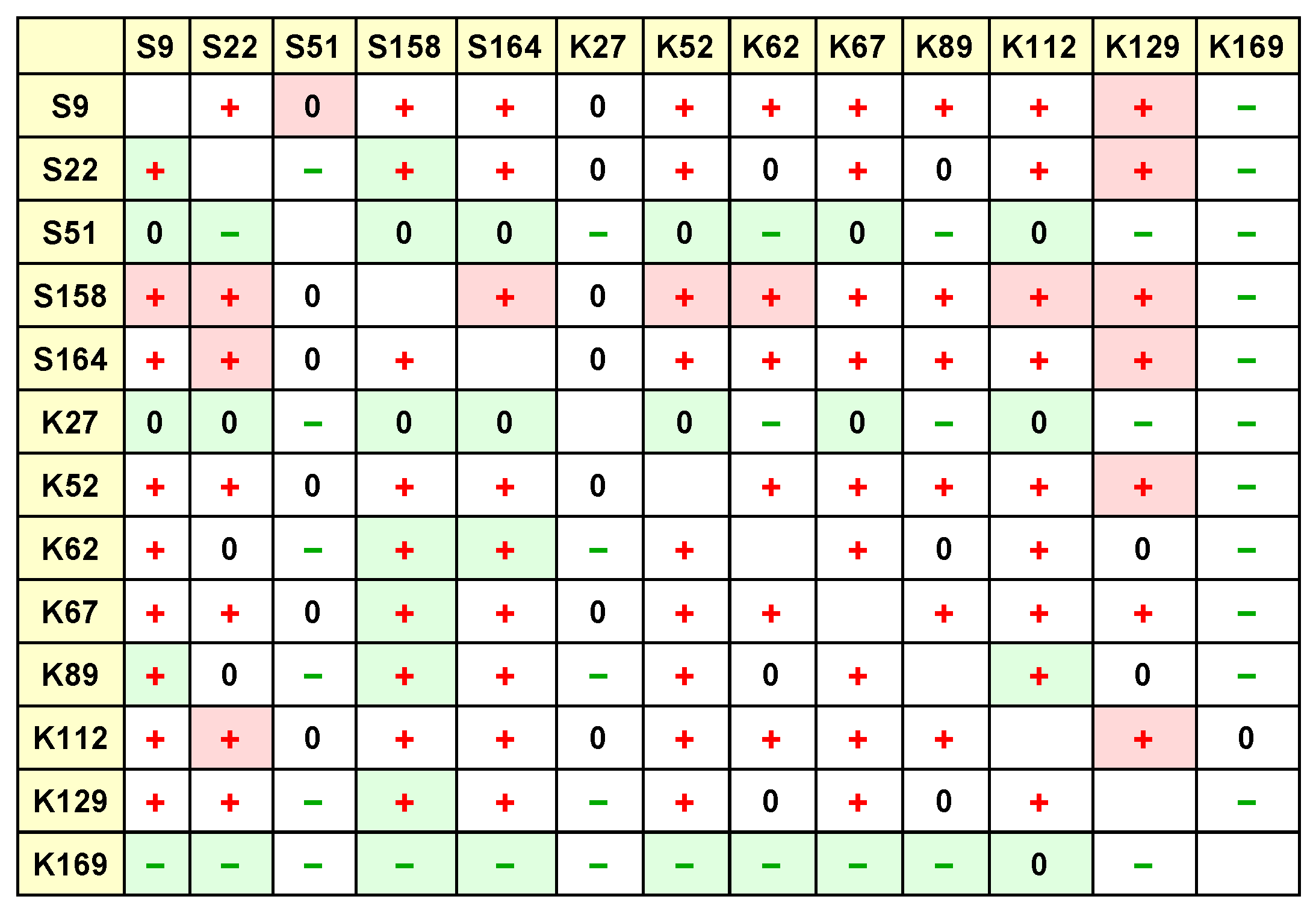
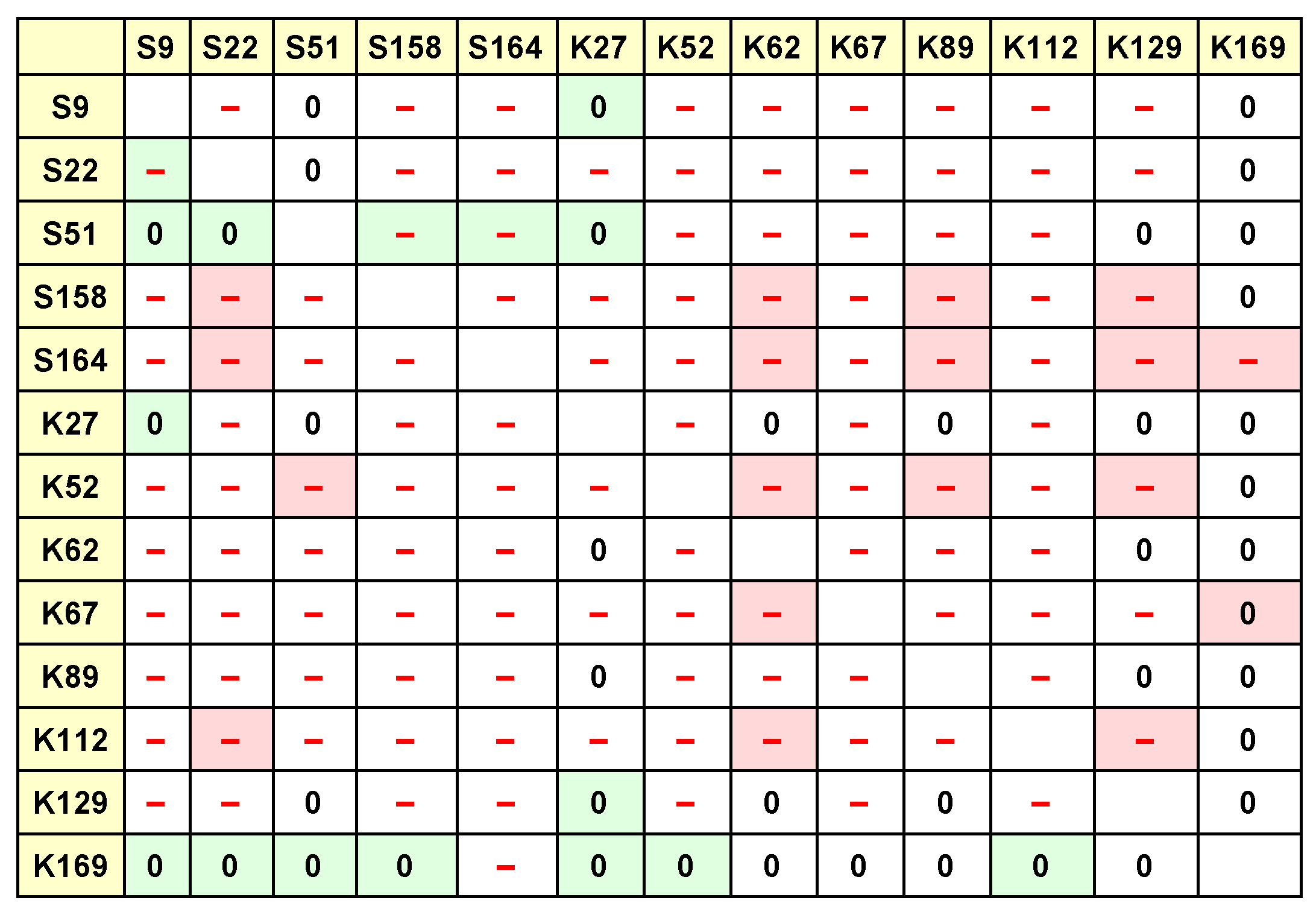
This test was done 13 times for each parameter, so that each of the strains was used once as the focal strain. The results of these tests are collected in the tables shown in
Figure 1 and
Figure 2. In these tables, each row collects the results of Tukey tests with a particular focal strain, e.g., the first row shows the results of tests where S9 was the focal strain, and the columns indicate the other strains being tested for interaction effects with S9.
3.1.2. Building the Competition Structures
Using the information gathered in
Figure 1 and
Figure 2, we represented the competition occurring between the SFIs using so-called tournament matrices. Such a matrix
M for
s species has dimensions
. If the species represented by row
i outcompetes the species represented by column
j, then
. On the other hand, if the species represented by row
i is outcompeted by the species represented by column
j, we have
. If
, then
. Using the information in
Figure 1 and
Figure 2, we can compile such a tournament or competition matrix. The question remains how precisely to do so.
We have two possibilities: to merge the information about the lag time
and mineralization rate
interaction effects, or to treat the parameters separately. The latter option is justified by considering that the parameters represent different biological attributes and different underlying processes [
25]. This is most noticeable in their opposing effects on mineralization performance in particular; an increased parameter
is considered a negative effect while an increased parameter
is considered a positive effect.
This approach results in two competition matrices, the first based on lag time
interaction effects, and the second based on mineralization rate
interaction effects. We look in
Figure 1 (
interaction effects) or
Figure 2 (
interaction effects) for pairs of SFIs that appear to interact with each other, and check what kind of interaction appears to be taking place: is it positive or negative with respect to each of the SFIs?
This corresponds in
Figure 1 and
Figure 2 to both the cell entries and the cell background colours. The cell entries indicate which kind of difference (if any) exists between the control community and the
community containing the particular species corresponding to the cell row and column. These relationships can be positive, negative, or not significant. The cell background colours indicate the difference (if any) between the
community containing the species corresponding to the cell column, and the
community containing the particular species corresponding to the cell row and column. These relationships can also be positive, negative, or not significant.
We then obtain the following matrices representing competition between the SFIs. When considering interactions based on lag time
effects, the matrix reads:
When considering interactions based on mineralization rate
effects, the matrix has the form:
An additional extension of our modelling approach that will bring it closer to reality is to also consider non-deterministic competition. Deterministic competition assumes that, if the competition structure specifies that A beats B, this will always occur: it will never be possible for B to beat A. This is reflected in the competition matrices
and
, which contain only 1’s (implying certain victory), −1’s (certain defeat) and 0’s (no competition). But this is not always realistic [
30,
31,
32]. Variation between individuals can result in an individual of species A that is a particularly weak competitor, and an individual of species B that is a particularly strong competitor. If these two specific individuals meet, the outcome of the competition can be in doubt. It may be more realistic to specify a so-called winning probability [
33,
34]: the probability that A beats B. Including a winning probability allows for different competition outcomes to occur, and the value of the winning probability allows us to account for the relative strengths of the individuals.
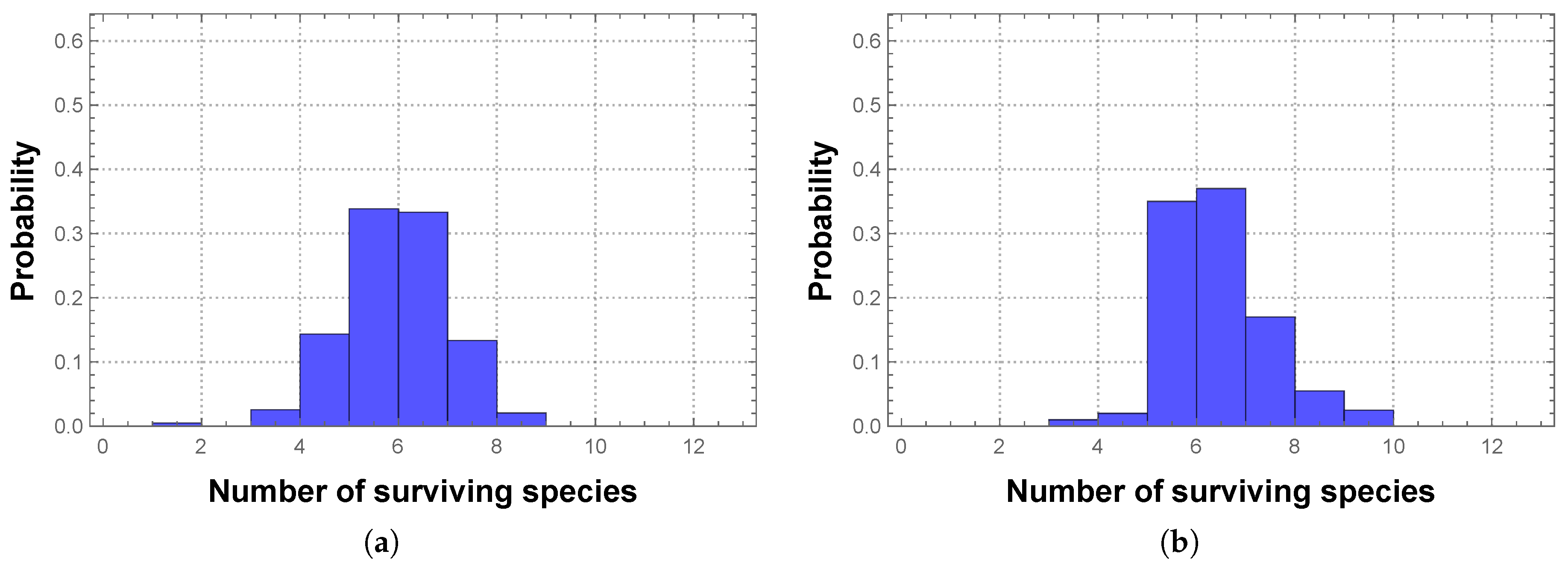
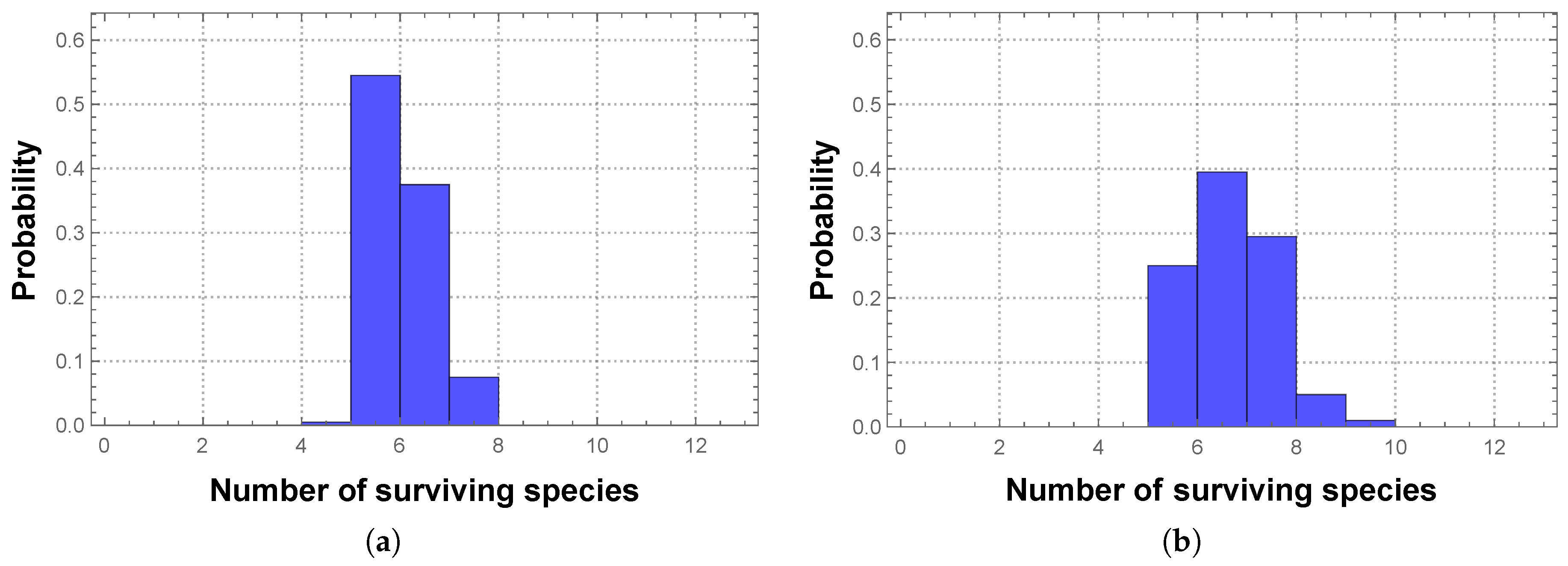
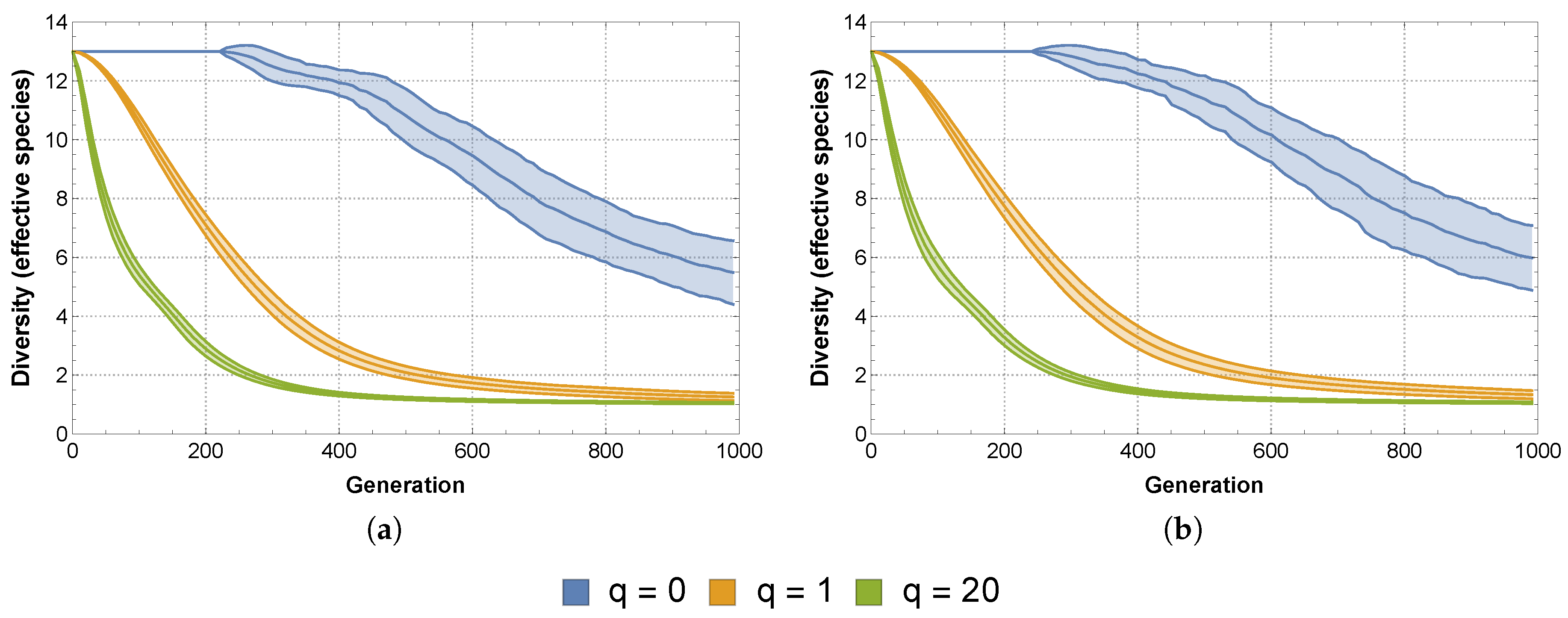
Therefore we will also consider non-deterministic competition between the SFIs, not only in terms of its effects on the diversity and stability of the community (and possible subcommunity), but in comparison with the same effects due to deterministic competition. Our immediate question is then how to assign the winning probabilities to the different pairwise competitions.
Using data related to the SFIs’ monoculture growth and survival curves, Vandermaesen et al. [
11] classified the “intrinsic competitiveness” of the SFIs and on this basis grouped them into strong, intermediate and weak competitors. Using this information, we can assign winning probabilities to each pairwise competition based on the relative differences in intrinsic competitiveness between the two strains. For example, competition between a weak intrinsic competitor and a strong intrinsic competitor will most likely result in the success of the latter. It should also be clear that this winning probability should be higher than the winning probability assigned to an intermediate intrinsic competitor when faced with a weak intrinsic competitor. Using this approach, we replace the 1’s and −1’s populating our matrices
and
with rational numbers of absolute value less than 1, corresponding to the appropriate winning probability.
Using this approach, we obtain the following matrices representing non-deterministic competition. When considering interactions based on lag time
effects, the matrix has the form:
When considering interactions based on mineralization rate
effects, the matrix has the form:

 ,
, 















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
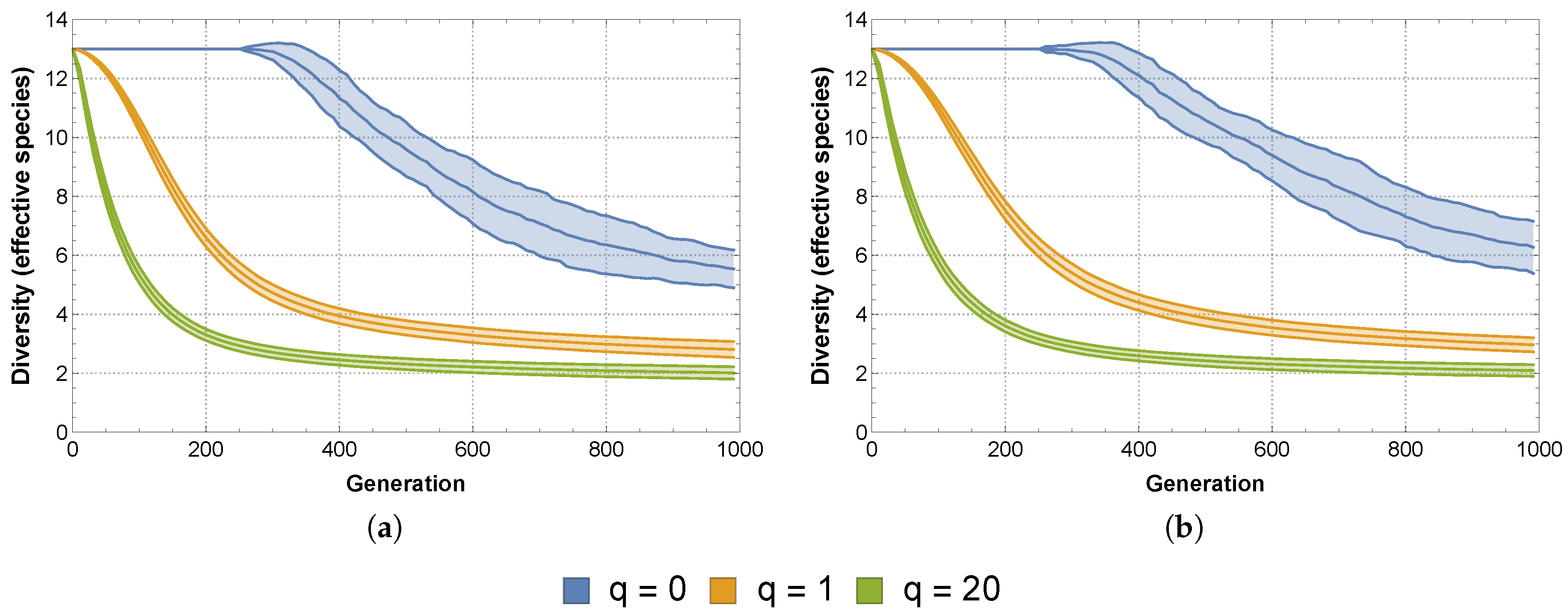{kind=link}
{kind=link}
{kind=link}
{kind=link}
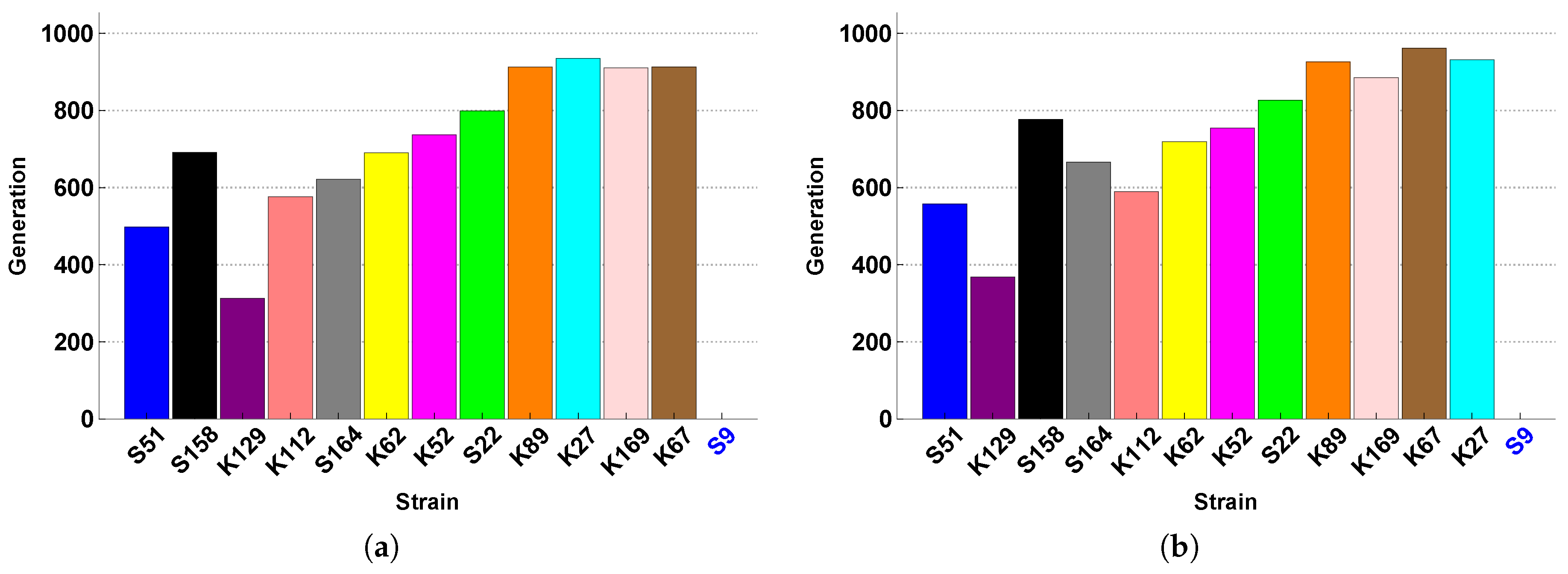{kind=link}
{kind=link}
{kind=link}
{kind=link}