Experimental Study of Substrate Limitation and Light Acclimation in Cultures of the Microalgae Scenedesmus obliquus—Parameter Identification and Model Predictive Control

 ,
,  ,
,  ,
,  ,
, 

Abstract
:1. Introduction
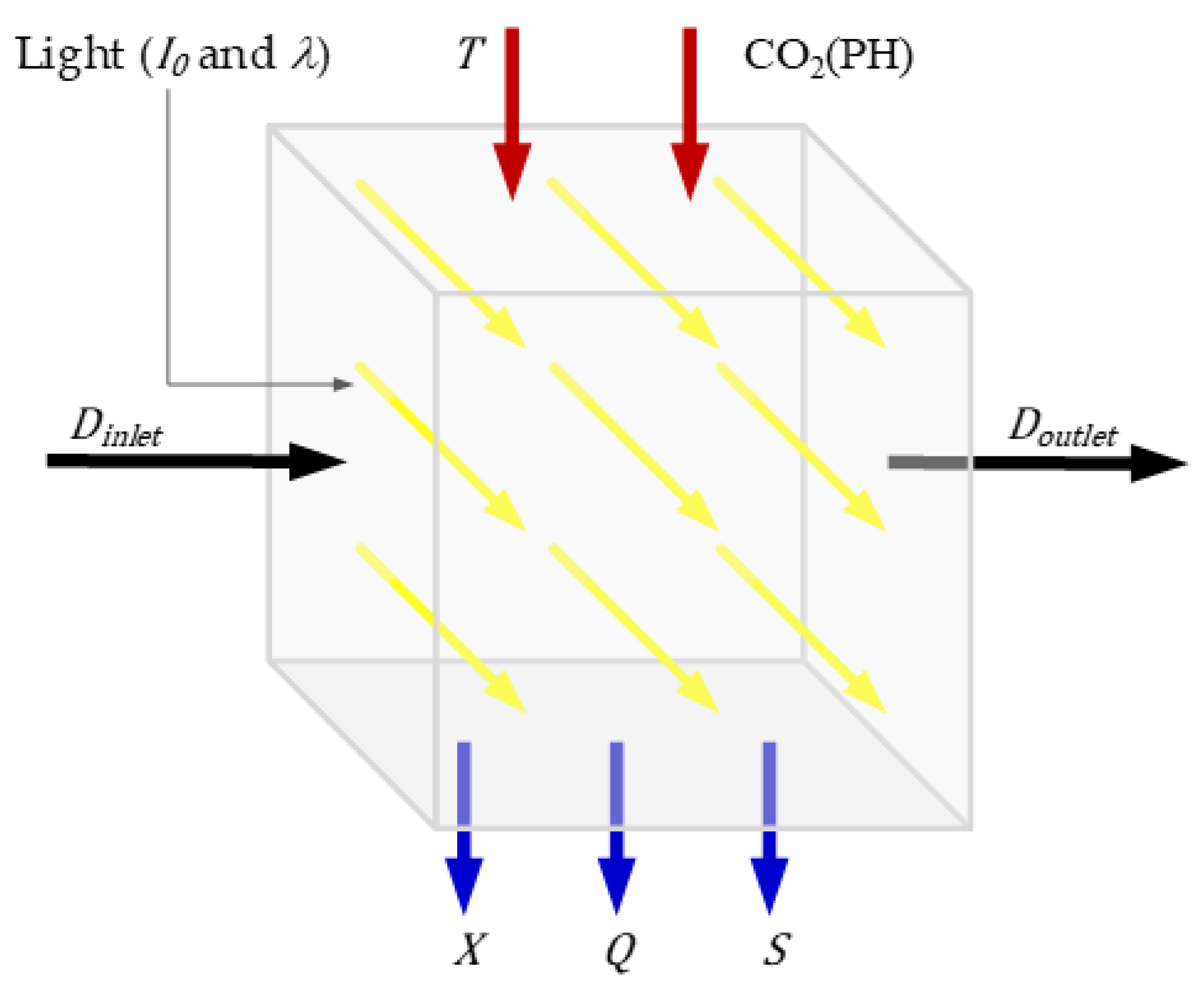
2. Model Description
3. Materials and Methods
3.1. Variable Measurement
3.1.1. Biomass Measurement
Operation Procedure
- Place cellulose acetate membrane filters (Whatman® (Sigma-Aldrich, St. Louis, MI, USA)) inside an oven at 105 °C and leave them overnight.
- Retrieve the dried filters and allow them to cool down in a desiccator for 8 h. Weigh the chosen filter and record the value .
- Extract a sample from the culture. Filter the sample using a vacuum pump; this leaves the biomass separated on the membrane aside from the medium.
- Dry the filter together with the algal biomass in a furnace at 105 °C until a constant weight is achieved and then cool in a desiccator for 20 min.
- Weight the sample on an analytical balance and record the value .
- Determine the biomass weight as the difference between and , with the corresponding factor adjustment.
3.1.2. Quota Measurement
Operation Procedure
- Take the sample from the medium culture into a 15 mL vial.
- Centrifuge it for 5 min at 5000 rpm. Remove the supernatant and gently shake the vial to dilute biomass. Use demineralized water to fill up to 15 mL.
- Repeat the previous step to wash the biomass and ensure a proper removal of substrate in the medium.
- Process the sample with the equipment to obtain the total nitrogen content ().
3.1.3. Substrate Measurement
Operation Procedure
- Collect a 5 mL sample from the reactor.
- Centrifuge the sample over 5 min at 5000 rpm.
- Take out 4 mL of supernatant and introduce it into a spectrophotometer cuvette.
- Measure the absorbance with the spectrophotometer at 210 nm and 270 nm.
- Determine the concentration of the substrate according to the correlation curve.
3.1.4. Chlorophyll Measurement
Operation Procedure
- Collect a 10 mL sample from the reactor in a 15 mL test tube.
- Centrifuge the sample over 5 min at 5000 rpm.
- Take out 9 mL of supernatant, add 3 mL of acetone and mix.
- Transfer the content to a 10 mL capsule of a ball mill.
- Add glass microspheres under 0.50 mm into the capsule, and shake it at 30 Hz for 20 min.
- Remove the capsule and transfer the content to a test tube.
- Fill the tube up to 10 mL of content adding acetone. This way the full content will be 90% acetone and 10% water, with the pigments solved in it.
- Centrifuge over for 5 min at 5000 rpm.
- Take out 4 mL of supernatant and transfer it to a 10 mm glass cuvette for the spectrophotometer.
- Measure the absorption at 630, 647, 664, and 750 nm.
3.1.5. Irradiance Measurement
3.2. Cell Culture and Medium Preparation
3.3. Laboratory Scale Process
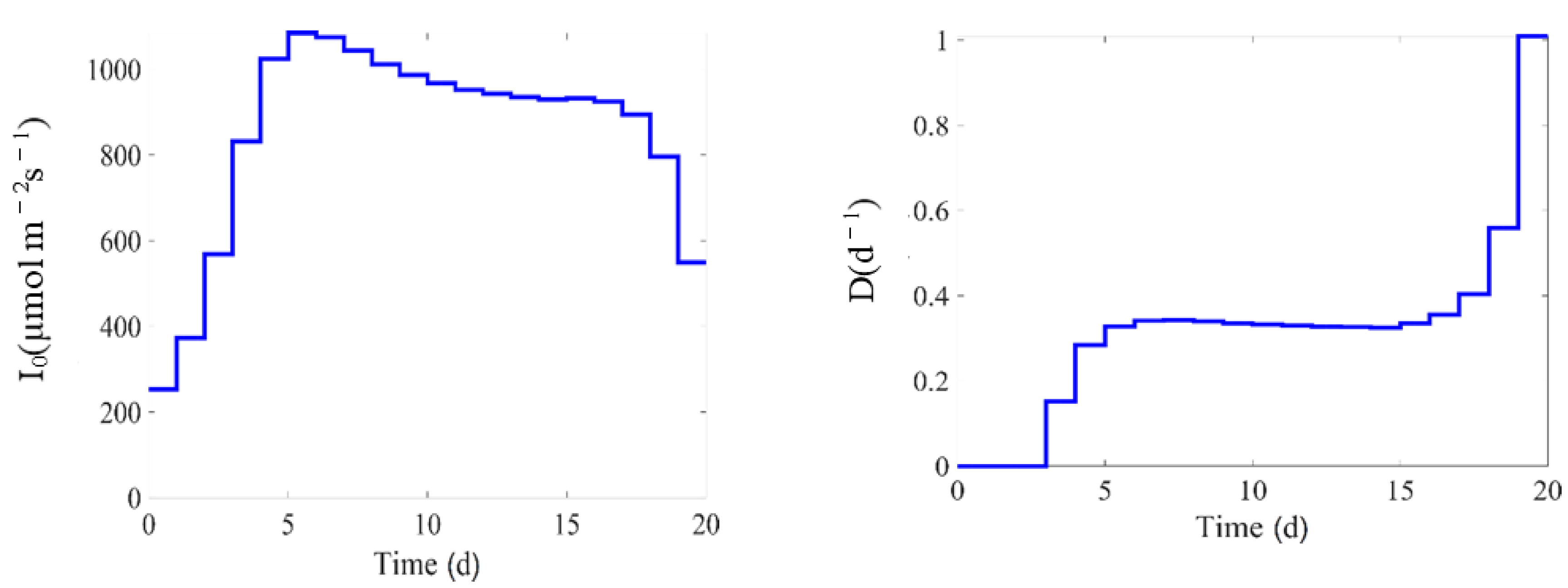
3.4. Experimental Design
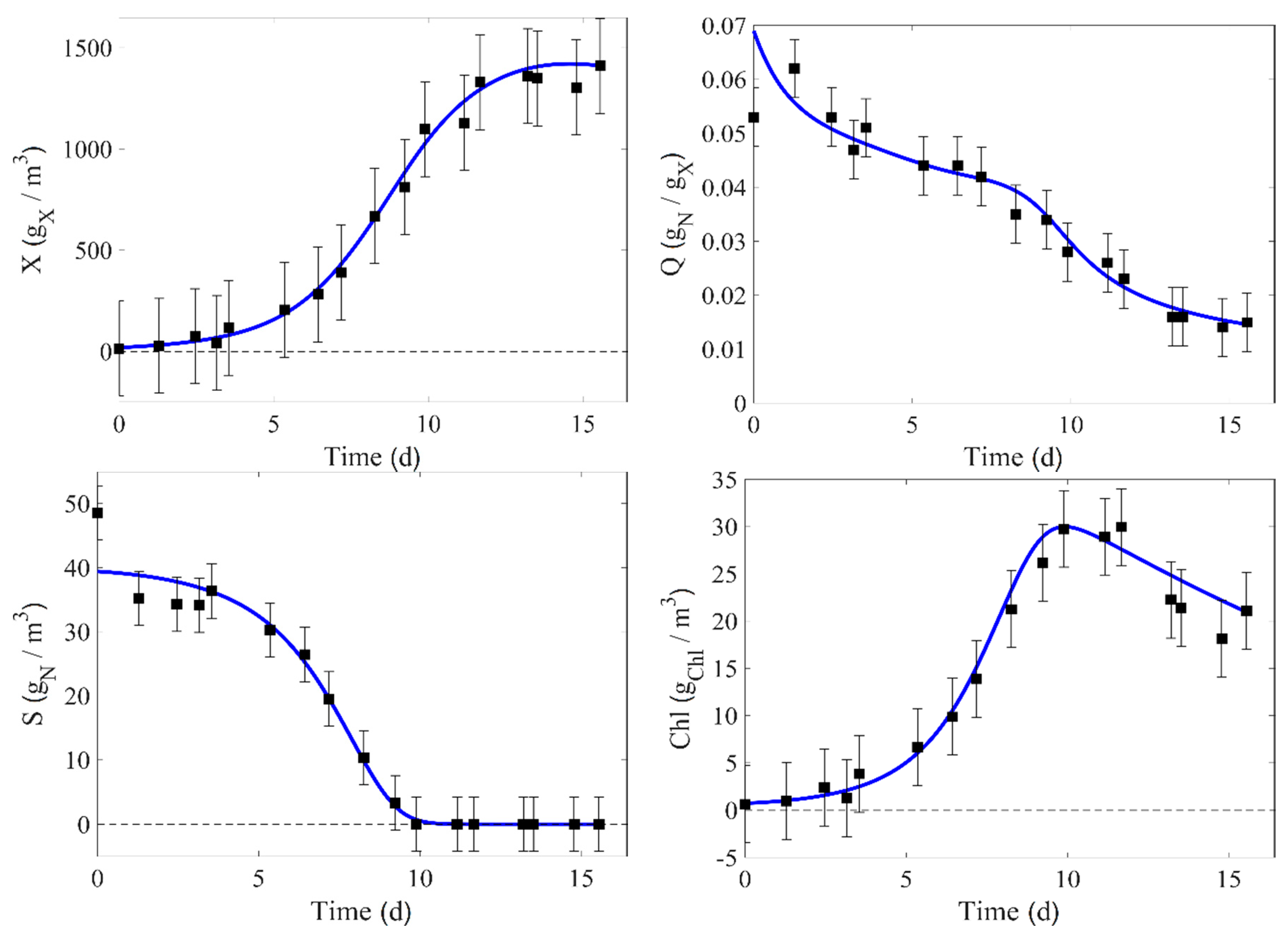
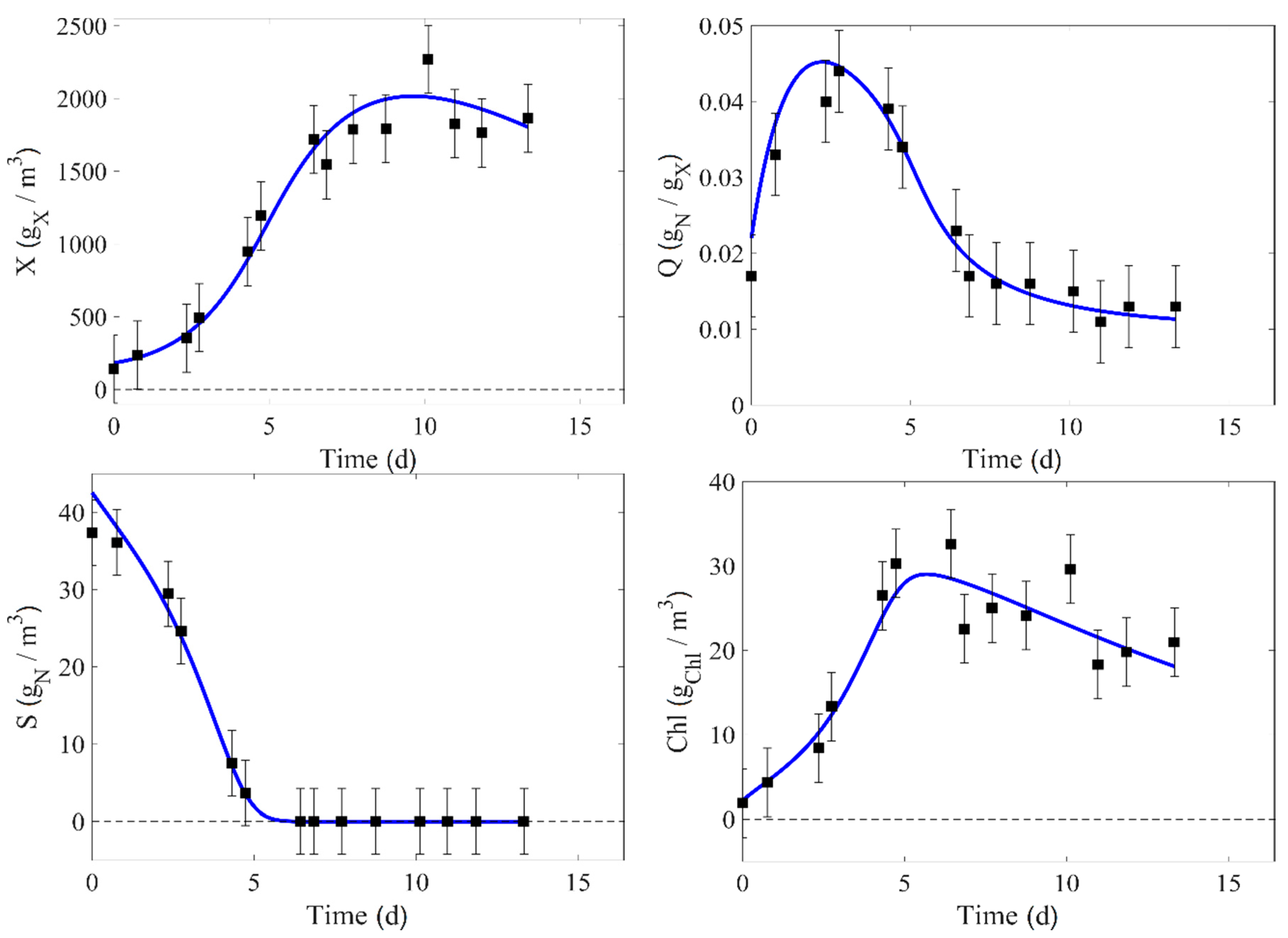
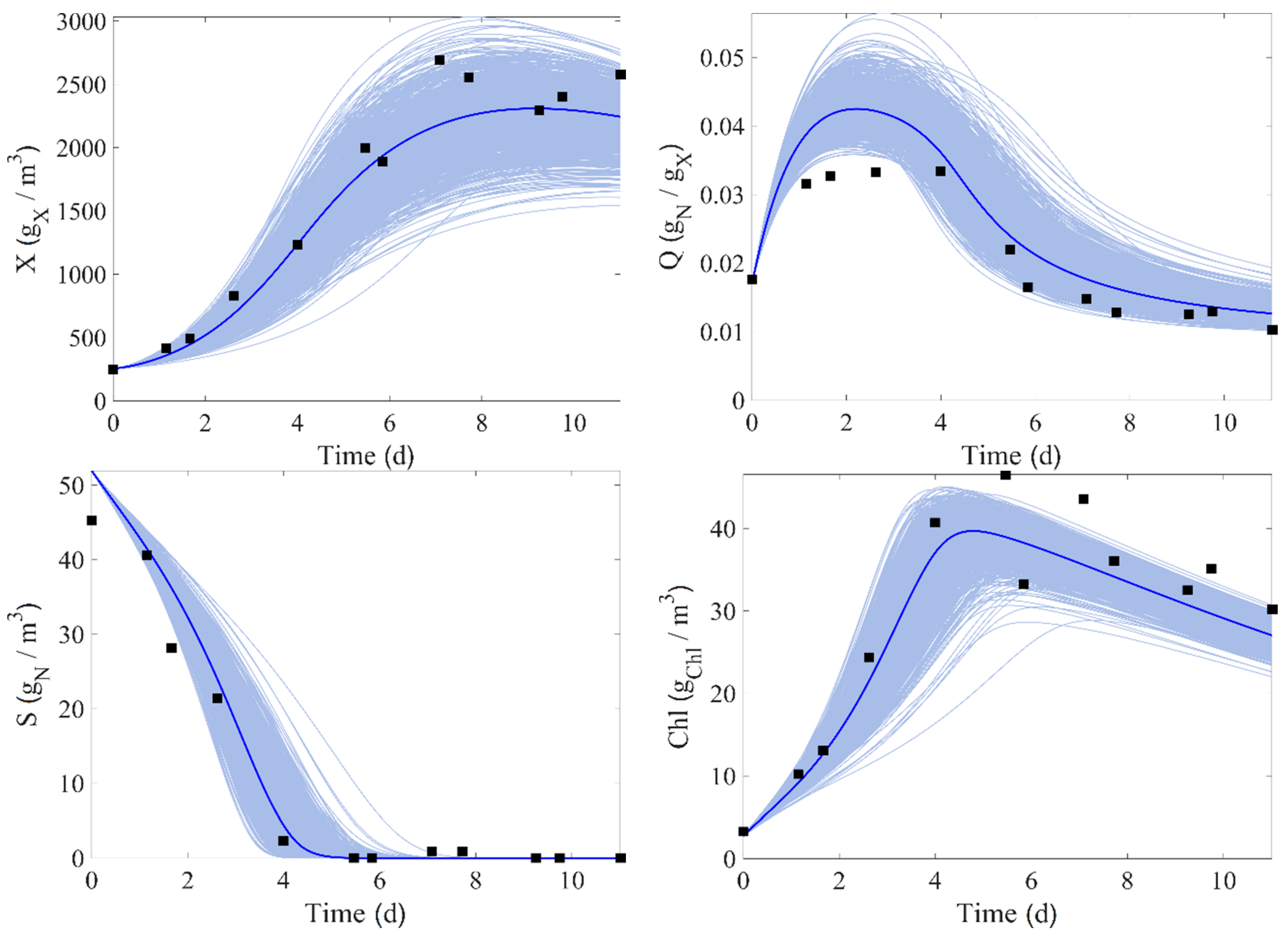
4. Results
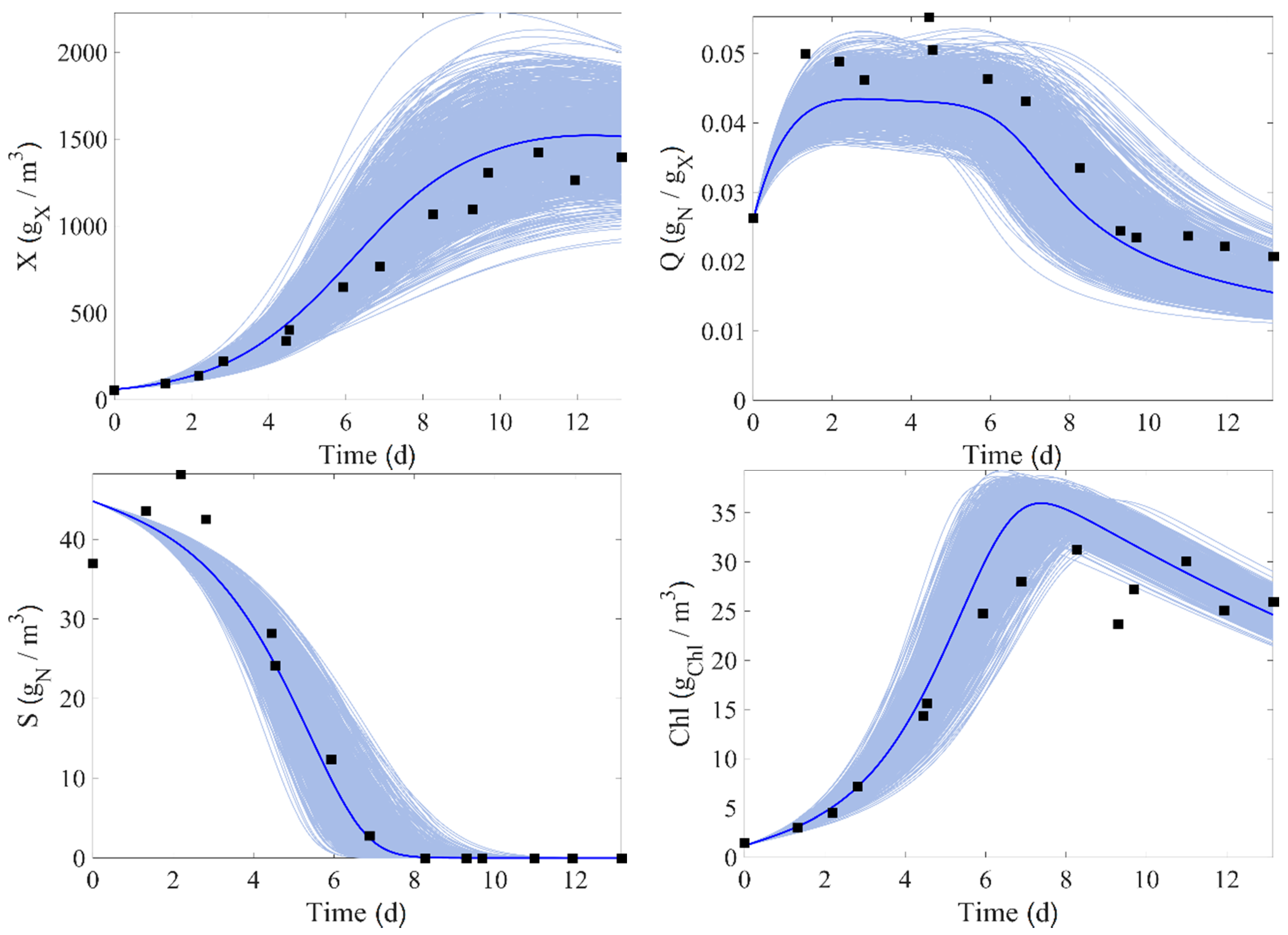
5. Model Parameter Identification
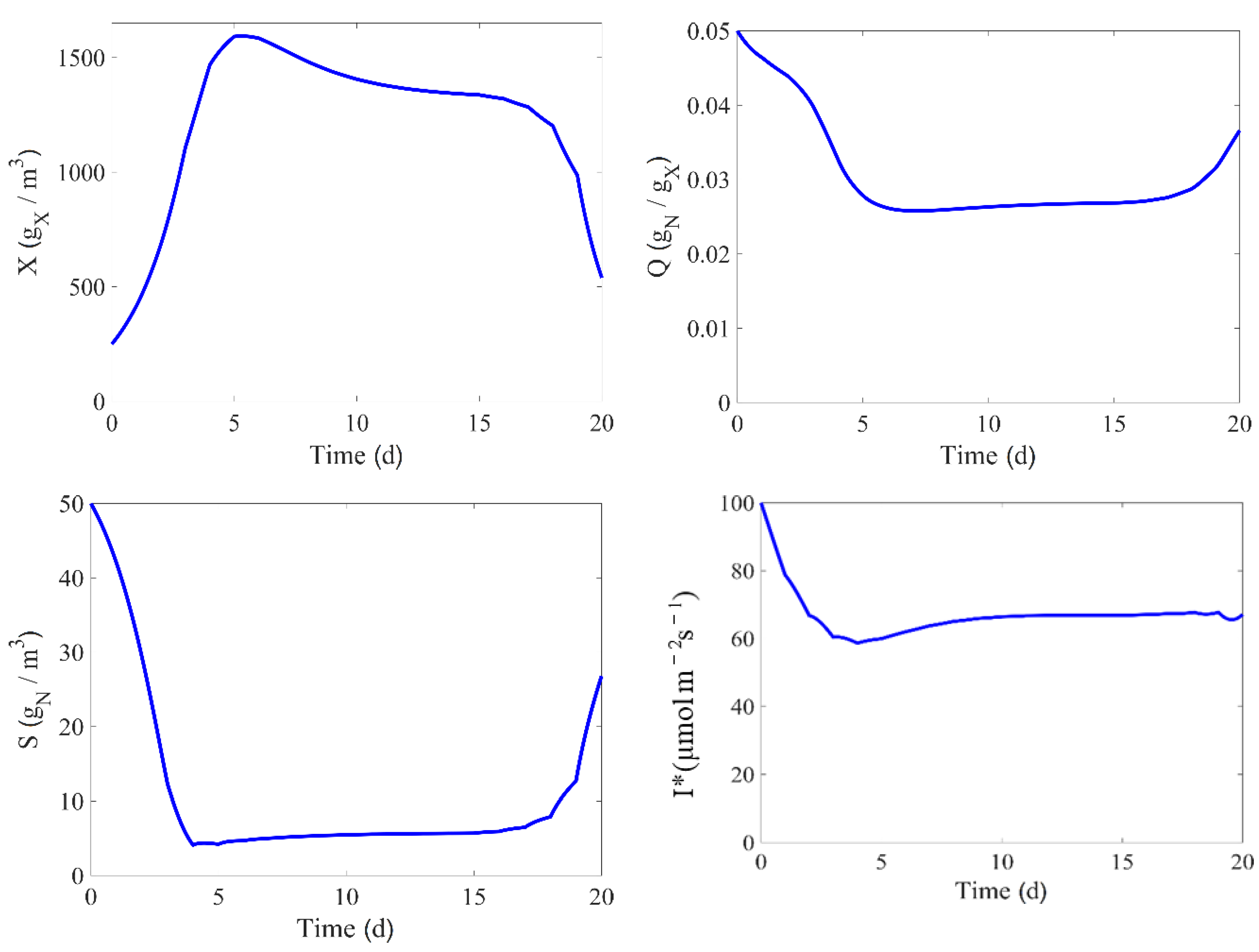
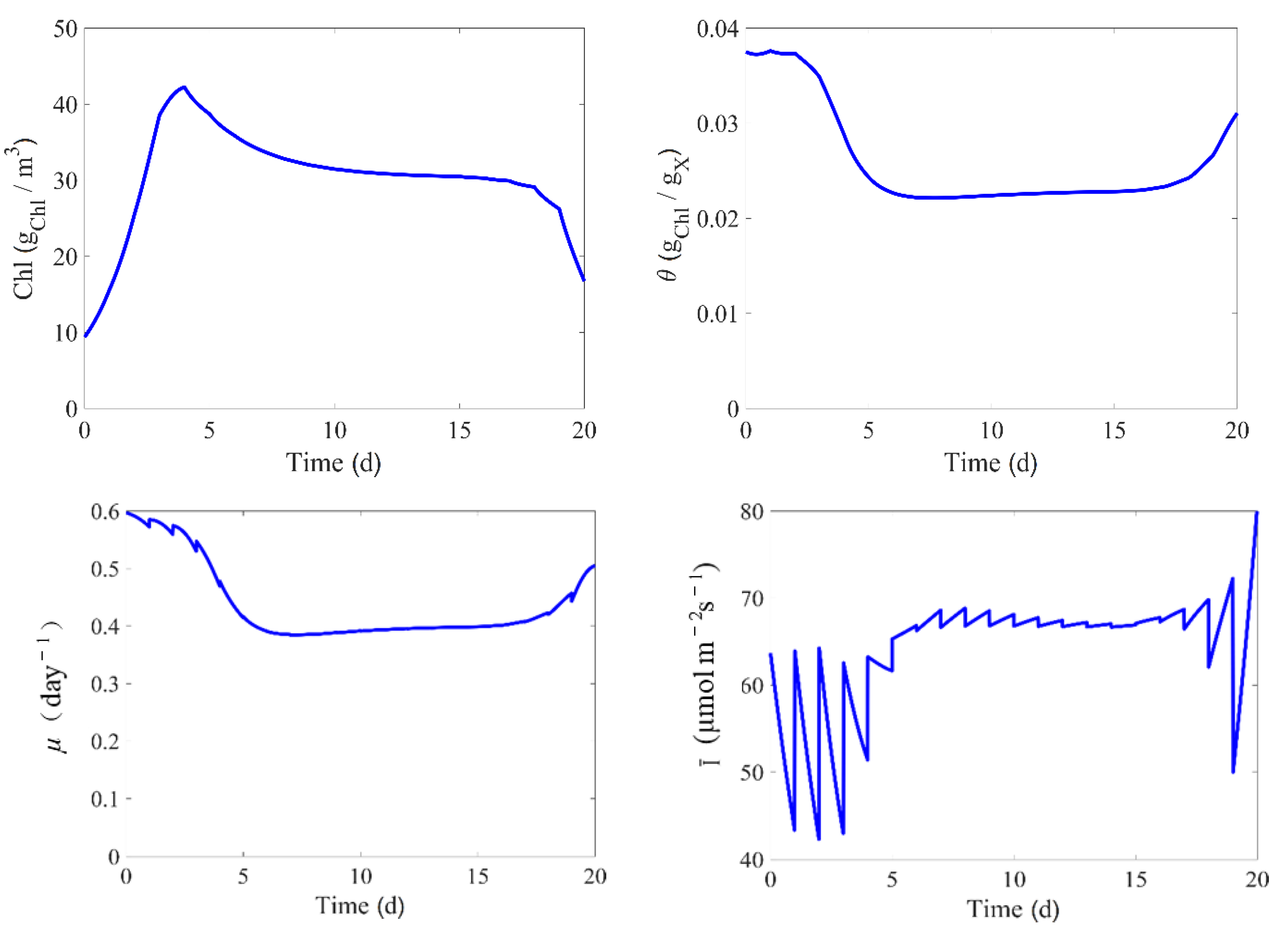
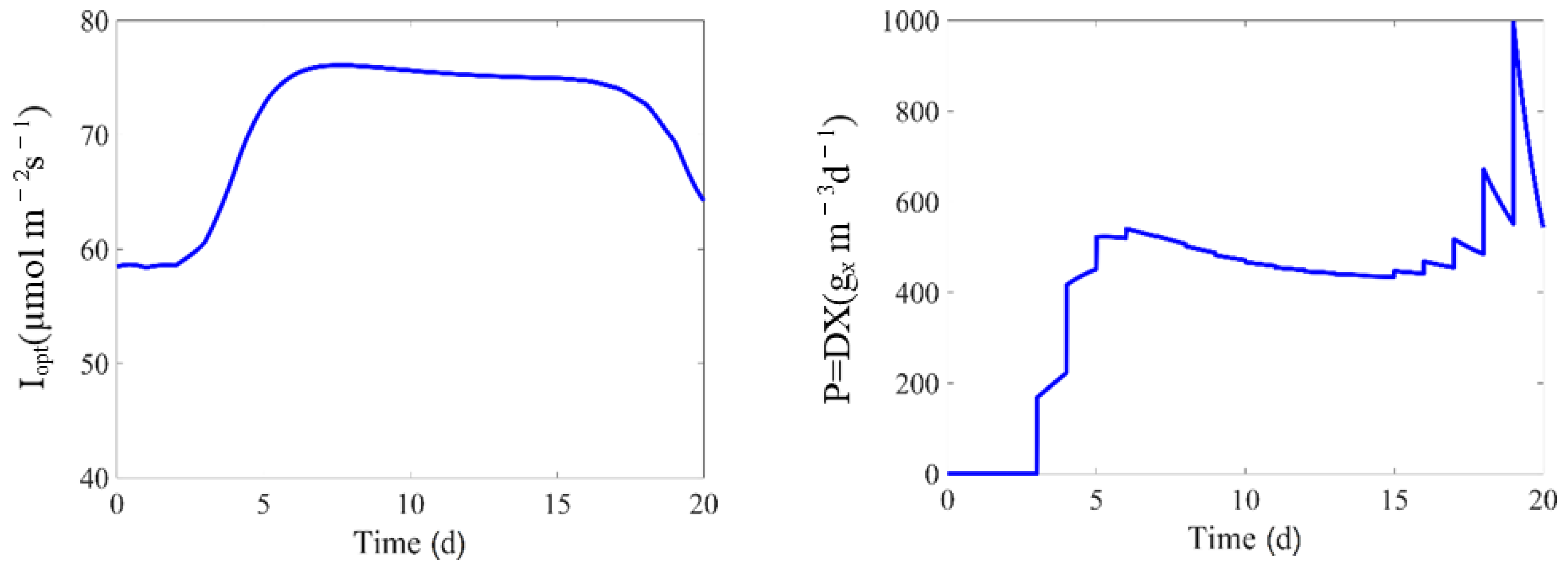
6. Non-Linear Model Predictive Control (NMPC)
6.1. Background on Model-Based Control of Cultures of Microalgae
6.2. NMPC Setup
6.3. NMPC Performance
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Definition | |
| Biomass bulk concentration | ||
| Nitrogen quota intracellular concentration | ||
| Nitrogen substrate bulk concentration | ||
| Photon flux density acclimation | ||
| Input | Definition | |
| Photon flux density | ||
| Dilution rate | ||
| Nitrogen concentration in the reactor inlet | ||
| Parameter | Definition | |
| Chlorophyll Parameters | ||
| Chlorophyll concentration | ||
| Chlorophyll quota | ||
| Chlorophyll adaptation rate | ||
| Average photon flux density throughout the culture | ||
| Growth Kinetics Parameters | ||
| Specific growth rate | ||
| Maximum specific growth rate | ||
| Average specific growth rate | ||
| Light Parameters | ||
| Nitrogen intake rate | ||
| Maximum nitrogen intake rate | ||
| Substrate uptake half saturation constant | ||
| Maximum nitrogen quota | ||
| Minimum nitrogen quota | ||
| Light Parameters | ||
| Optimum photon flux density | ||
| Normalized growth half saturation constant | ||
| Growth half saturation constant | ||
| Photon flux density saturation constant over growth | ||
| Chlorophyll saturation function | ||
| Maximum chlorophyll saturation function | ||
| Chlorophyll saturation function constant | ||
| Turbidity Parameters | ||
| Light attenuation rate light attenuation coefficient | ||
| Optical depth | ||
| Culture depth | ||
| Attenuation coefficient due to chlorophyll | ||
| Attenuation coefficient due to biomass | ||
| Attenuation coefficient due to background turbidity | ||
| Average photon flux density saturation function constant | ||
References
- Chisti, Y. Biodiesel from microalgae beats bioethanol. Trends Biotechnol. 2008, 26, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Demirbas, A. Use of algae as biofuel sources. Energy Convers. Manag. 2010, 51, 2738–2749. [Google Scholar] [CrossRef]
- Lam, M.; Lee, K.T. Microalgae biofuels: A critical review of issues, problems and the way forward. Biotechnol. Adv. 2012, 30, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Shin, J.; Kim, J.-D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, G.E.; Ranglová, K.; Manoel, J.C.; Grivalský, T.; Kopecký, J.; Masojídek, J. Bioethanol production from microalgae polysaccharides. Folia Microbiol. 2019, 64, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Mata, T.; Martins, A.; Caetano, N. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Martínez, C.; Bernard, O.; Mairet, F. Maximizing microalgae productivity by shading outdoor cultures. IFAC-PapersOnLine 2017, 50, 8734–8739. [Google Scholar] [CrossRef]
- Olivieri, G.; Salatino, P.; Marzocchella, A. Advances in photobioreactors for intensive microalgal production: Configurations, operating strategies and applications. J. Chem. Technol. Biotechnol. 2014, 89, 178–195. [Google Scholar] [CrossRef]
- Hartmann, P.; Nikolaou, A.; Chachuat, B.; Bernard, O. A Dynamic Model Coupling Photoacclimation and Photoinhibition in Microalgae. In Proceedings of the 12th European Control Conference, Zürich, Switzerland, 17–19 July 2013; Institute of Electrical and Electronics Engineers: Zürich, Switzerland, 2013; pp. 4178–4183. [Google Scholar] [CrossRef] [Green Version]
- Peeters, J.C.H.; Eilers, P.H.C. The relationship between light intensity and photosynthesis—A simple mathematical model. Hydrobiol. Bull. 1978, 12, 134–136. [Google Scholar] [CrossRef]
- Eilers, P.H.C.; Peeters, J.C.H. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecol. Model. 1988, 42, 199–215. [Google Scholar] [CrossRef]
- Eilers, P.H.C.; Peeters, J.C.H. Dynamic behaviour of a model for photosynthesis and photoinhibition. Ecol. Model. 1993, 69, 113–133. [Google Scholar] [CrossRef]
- Kok, B. On the inhibition of photosynthesis by intense light. Biochim. Biophys. Acta 1956, 21, 234–244. [Google Scholar] [CrossRef]
- Long, S.P.; Humphries, S.; Falkowski, P.G. Photoinhibition of Photosynthesis in Nature. Annu. Rev. Plant Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Gojkovic, Ž.; Garbayo-Nores, I.; Gómez-Jacinto, V.; García-Barrera, T.; Gómez-Ariza, J.L.; Márová, I.; Vilchez-Lobato, C. Continuous production of selenomethionine-enriched Chlorella sorokiniana biomass in a photobioreactor. Process Biochem. 2013, 48, 1235–1241. [Google Scholar] [CrossRef]
- Guterman, H.; Vonshak, A.; Ben-Yaakov, S. A macromodel for outdoor algal mass production. Biotechnol. Bioeng. 1990, 35, 809–819. [Google Scholar] [CrossRef]
- Benavides, M.; Hantson, A.-L.; Van Impe, J.F.M.; Vande Wouwer, A. Parameter identification of Droop model: An experimental case study. Bioprocess Biosyst. Eng. 2015, 38, 1783–1793. [Google Scholar] [CrossRef] [Green Version]
- Mairet, F.; Bernard, O.; Masci, P.; Lacour, T.; Sciandra, A. Modelling neutral lipid production by the microalga Isochrysis aff. galbana under nitrogen limitation. Bioresour. Technol. 2011, 102, 142–149. [Google Scholar] [CrossRef]
- Kumar, A.; Chandan, G.; Chitres, G.; Chakraborty, A.; Pathak, A.K. Modelling of microalgal growth and lipid production in Dunaliella tertiolecta using nitrogen-phosphorus-potassium fertilizer medium in sintered disk chromatographic glass bubble column. Bioresour. Technol. 2016, 218, 1021–1036. [Google Scholar] [CrossRef]
- Sinha, S.; Kumar, M.; Guria, C.; Kumar, A.; Banerjee, C. Biokinetic model-based multi-objective optimization of Dunaliella tertiolecta cultivation using elitist non-dominated sorting genetic algorithm with inheritance. Bioresour. Technol. 2017, 242, 206–217. [Google Scholar] [CrossRef]
- Hu, D.; Liu, H.; Yang, C.; Hu, E. The design and optimization for light-algae bioreactor controller based on Artificial Neural Network-Model Predictive Control. Acta Astronaut. 2008, 63, 1067–1075. [Google Scholar] [CrossRef]
- Incoll, L.D.; Long, S.P.; Ashmore, M.R. SI units in publications in plant science. In Commentaries in Plant Science, 1st ed.; Smith, H., Ed.; Pergamon Press: Oxford, UK, 1981; Volume 2, pp. 87–89. [Google Scholar] [CrossRef]
- Feudjio Letchindjio, C.G.; Deschênes, J.S.; Dewasme, L.; Vande Wouwer, A. Extremum seeking based on a Hammerstein-Wiener representation. IFAC-PapersOnLine 2018, 51, 744–749. [Google Scholar] [CrossRef]
- Bernard, O.; Masci, P.; Sciandra, A. A Photobioreactor Model in Nitrogen Limited Conditions. In Proceedings of the 6th Vienna International Conference on Mathematical Modelling, Vienna, Austria, 11–13 February 2009; European Mathematical Society: Helsinki, Finland, 2009; pp. 1521–1530. [Google Scholar]
- Bernard, O. Hurdles and challenges for modelling and control of microalgae for CO2 mitigation and biofuel production. J. Process Control 2011, 21, 1378–1389. [Google Scholar] [CrossRef]
- Bernard, O.; Remond, B. Validation of a simple model accounting for light and temperature effect on microalgal growth. Bioresour. Technol. 2012, 123, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Droop, M.R. Vitamin b 12 and marine ecology. iv. The kinetics of uptake, growth and inhibition in monochrysis lutheri. J. Mar. Biol. Assoc. UK 1968, 48, 689–733. [Google Scholar] [CrossRef]
- Deschênes, J.-S.; Vande Wouwer, A. Parameter Identification of a Dynamic Model of Cultures of Microalgae Scenedesmus obliquus—An experimental study. IFAC-PapersOnLine 2016, 49, 1050–1055. [Google Scholar] [CrossRef]
- Dewasme, L.; Feudjio Letchindjio, C.G.; Zuniga, I.T.; Vande Wouwer, A. Micro-Algae Productivity Optimization Using Extremum-Seeking Control. In Proceedings of the 25th Mediterranean Conference on Control and Automation, Valletta, Malta, 3–6 July 2017; Institute of Electrical and Electronics Engineers: Valletta, Malta, 2017; pp. 672–677. [Google Scholar] [CrossRef]
- Feudjio Letchindjio, C.G.; Bogaerts, P.; Deschênes, J.S.; Vande Wouwer, A. Design of a Robust Lipschitz Observer-Application to monitoring of culture of micro-algae Scenesdesmus obliquus. IFAC-PapersOnLine 2016, 49, 1056–1061. [Google Scholar] [CrossRef]
- Feudjio Letchindjio, C.G.; Dewasme, L.; Deschênes, J.S.; Vande Wouwer, A. An Extremum Seeking Strategy Based on Block-Oriented Models: Application to Biomass Productivity Maximization in Microalgae Cultures. Ind. Eng. Chem. Res. 2019, 58, 13481–13494. [Google Scholar] [CrossRef]
- Bastin, V.G.; Dochain, D. Dynamical models of bioreactors. In On-Line Estimation and Adaptive Control of Bioreactors, 1st ed.; Elsevier: Amsterdam, The Netherlands, 1990; p. 30. [Google Scholar]
- Kützing, F.T. Synopsis Diatomacearum oder Versuch einer Systematischen Zusammenstellung der Diatomeen. Linnaea 1833, 8, 529–620. [Google Scholar]
- Eaton, A.D.; Clesceri, L.S.; Greenberg, A.E. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association, American Water Works Association, and Water Pollution Control Federation: Washington, DC, USA, 1998. [Google Scholar]
- Edwards, A.C.; Hooda, P.S.; Cook, Y. Determination of Nitrate in Water Containing Dissolved Organic Carbon by Ultraviolet Spectroscopy. Int. J. Environ. Anal. Chem. 2001, 80, 49–59. [Google Scholar] [CrossRef]
- Aminot, A.; Rey, F. Chlorophyll A: Determination by Spectroscopic Methods; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2001; 17p. [Google Scholar] [CrossRef]
- Hosikian, A.; Lim, S.; Halim, R.; Danquah, M.K. Chlorophyll Extraction from Microalgae: A Review on the Process Engineering Aspects. Int. J. Chem. Eng. 2010, 2010, 391632. [Google Scholar] [CrossRef] [Green Version]
- Simon, D.; Helliwell, S. Extraction and quantification of chlorophyll a from freshwater green algae. Water Res. 1998, 32, 2220–2223. [Google Scholar] [CrossRef]
- Ritchie, R.J. Universal chlorophyll equations for estimating chlorophylls a, b, c, and d and total chlorophylls in natural assemblages of photosynthetic organisms using acetone, methanol, or ethanol solvents. Photosynthetica 2008, 46, 115–126. [Google Scholar] [CrossRef]
- Stein, J.R. Handbook of Phycological Methods: Culture Methods and Growth Measurements; Cambridge University Press: Cambridge, UK, 1973. [Google Scholar]
- Kim, T.-H.; Lee, Y.; Han, S.-H.; Hwang, S.-J. The effects of wavelength and wavelength mixing ratios on microalgae growth and nitrogen, phosphorus removal using Scenedesmus sp. for wastewater treatment. Bioresour. Technol. 2013, 130, 75–80. [Google Scholar] [CrossRef]
- Şimşek, G.K.; Cetin, A. Effect of Different Wavelengths of Light on Growth, Pigment Content and Protein Amount of Chlorella vulgaris. Fresenius Environ. Bull. 2017, 26, 7974–7980. [Google Scholar]
- Van Impe, J.F.M. Power and limitations of model based bioprocess optimization. Math. Comput. Simul. 1996, 42, 159–169. [Google Scholar] [CrossRef]
- Flynn, K.J.; Davidson, K.; Leftley, J.W. Carbon-nitrogen relations at whole-cell and free-amino-acid levels during batch growth of Isochrysis galbana (Prymnesiophyceae) under conditions of alternating light and dark. Mar. Biol. 1994, 118, 229–237. [Google Scholar] [CrossRef]
- Ben-Yaakov, S.; Guterman, H.; Vonshak, A.; Richmond, A. An automatic method for on-line estimation of the photosynthetic rate in open algal ponds. Biotechnol. Bioeng. 1985, 27, 1136–1145. [Google Scholar] [CrossRef]
- Guterman, H.; Ben-Yaakov, S.; Vonshak, A. Automatic on-line growth estimation method for outdoor algal biomass production. Biotechnol. Bioeng. 1989, 34, 143–152. [Google Scholar] [CrossRef]
- Becerra-Celis, G.; Tebbani, S.; Joannis-Cassan, C.; Isambert, A.; Siguerdidjane, H. Control Strategy for Continuous Microalgae Cultivation Process in a Photobioreactor. In Proceedings of the 17th IEEE International Conference on Control Applications, San Antonio, TX, USA, 3–5 September 2008; Institute of Electrical and Electronics Engineers: Piscataway, NJ, USA, 2008; pp. 684–689. [Google Scholar] [CrossRef]
- Mairet, F.; Muñoz-Tamayo, R.; Bernard, O. Adaptive control for optimizing microalgae production. IFAC Proc. Vol. 2013, 46, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Ifrim, G. Feedback Linearizing Control of Light-to-Microalgae Ratio in Artificially Lighted Photobioreactors. In Proceedings of the 12th IFAC Symposium on Computer Applications in Biotechnology, Mumbai, India, 16–18 December 2013; The International Federation of Automatic Control: Laxenburg, Austria, 2013; pp. 169–174. [Google Scholar] [CrossRef]
- Sewsynker-Sukai, Y.; Faloye, F.; Kana, E.B.G. Artificial neural networks: An efficient tool for modelling and optimization of biofuel production (a mini review). Biotechnol. Biotechnol. Equip. 2017, 31, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Juneja, A.; Murthy, G.S. Model predictive control coupled with economic and environmental constraints for optimum algal production. Bioresour. Technol. 2018, 250, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Ifrim, G.; Titica, M.; Barbu, M.; Ceanga, E.; Caraman, S. Optimization of a Microalgae Growth Process in Photobioreactors. IFAC-PapersOnLine 2016, 49, 218–223. [Google Scholar] [CrossRef]
- Deschênes, J.S.; Vande Wouwer, A. Dynamic Optimization of Biomass Productivity in Continuous Cultures of Microalgae Isochrysis galbana through Modulation of the Light Intensity. IFAC-PapersOnLine 2015, 48, 1093–1099. [Google Scholar] [CrossRef]
- Tebbani, S.; Titica, M.; Ifrim, G.; Barbu, M.; Caraman, S. Optimal operation of a lumostatic microalgae cultivation process. In Developments in Model-Based Optimization and Control—Distributed Control and Industrial Applications; Springer International Publishing: New York, NY, USA, 2015; pp. 209–235. [Google Scholar]
- Tebbani, S.; Titica, M.; Caraman, S. Control of the Light-to-Microalgae Ratio in a Photobioreactor. In Proceedings of the 18th International Conference on System Theory, Control and Computing, Sinaia, Romania, 17–19 October 2014; Institute of Electrical and Electronics Engineers: Piscataway, NJ, USA, 2014; pp. 393–398. [Google Scholar] [CrossRef]
- Camacho, E.F.; Bordons, C. Model based predictive controllers. In Model Predictive Control in the Process Industry, 1st ed.; Springer: London, UK, 1995; pp. 1–100. [Google Scholar] [CrossRef]









| Formula | Concentration [g/L] | Concentration [g/L] | |
|---|---|---|---|
| 0.025 | micro-nutrients 1 | ||
| 0.075 | 8.829 | ||
| 0.025 | 1.441 | ||
| 0.750 | 6.093 | ||
| 0.075 | 1.571 | ||
| 0.176 | 0.400 | ||
| micro-nutrients 1 | 1 mL | 11.439 | |
| micro-nutrients 2 | 1 mL | micro-nutrients 2 | |
| micro-nutrients 3 | 1 mL | 63.651 | |
| 31.026 | |||
| micro-nutrients 3 | |||
| 4.976 | |||
| 10 mL/L | |||
| Vitamins | |||
| Vitamin B1 (thiamine ) | 8.934 | ||
| Vitamin H (biotine) | 0.025 | ||
| Vitamin B12 (cyanocobalamine) | 0.018 |
| Initial Conditions | Input Variables | ||||||
|---|---|---|---|---|---|---|---|
| Run | |||||||
| 1 | 14.6 | 10.1 | 0.025 | 33.2 | 100 | 100 | 0 |
| 2 | 14.6 | 12.3 | 0.030 | 36.5 | 100 | 150 | 0 |
| 3 | 15.5 | 30.7 | 0.061 | 41.3 | 200 | 200 | 0 |
| 4 | 15.5 | 18.7 | 0.069 | 39.4 | 200 | 250 | 0 |
| 5 | 13.1 | 60.3 | 0.026 | 44.8 | 100 | 220 | 0 |
| 6 | 13.1 | 76.7 | 0.023 | 47.5 | 100 | 350 | 0 |
| 7 | 11.0 | 225.0 | 0.016 | 49.9 | 250 | 520 | 0 |
| 8 | 11.0 | 255.0 | 0.017 | 52.0 | 150 | 600 | 0 |
| 9 | 13.3 | 183.8 | 0.022 | 42.6 | 200 | 750 | 0 |
| Parameter | Estimated Value | CV (%) | CI |
|---|---|---|---|
| 1.69 | 7.0 | 0.23 | |
| 17.69 | 10.0 | 3.5 | |
| 0.0104 | 4.0 | 0.0008 | |
| 0.1172 | 8.2 | 0.0189 | |
| 0.0573 | 6.4 | 0.0072 | |
| 1.36 | 17.3 | 0.46 | |
| 94.3 | 14.6 | 27.0 | |
| 0.0779 | 2.1 | 0.0032 | |
| 1.158 | 1.6 | 0.037 | |
| 184 | 7.7 | 28 | |
| 12.6 | 37.0 | 9.1 | |
| 0.72 | 23.8 | 0.34 | |
| 15.1 | 45.9 | 13.6 | |
| 10.6 | 24.5 | 5.1 |
| 1 | 8.5 | 0.019 | 28.35 | 104 |
| 2 | 9.8 | 0.035 | 28.11 | 97 |
| 3 | 36.4 | 0.047 | 38.08 | 202 |
| 4 | 14.8 | 0.053 | 48.55 | 209 |
| 5 | 56.3 | 0.026 | 36.98 | 92 |
| 6 | 92.1 | 0.020 | 37.19 | 103 |
| 7 | 249.9 | 0.019 | 45.40 | 261 |
| 8 | 249.0 | 0.018 | 45.26 | 157 |
| 9 | 141.7 | 0.017 | 37.38 | 193 |
| Parameter | [26] 1 | [29] 2 | Current Study 2 | |||
|---|---|---|---|---|---|---|
| Value | CV (%) | Value | CV (%) | Value | CV (%) | |
| 1.7 | - | 1.47 | 5.9 | 1.69 | 7.0 | |
| 0.0012 | - | 0.09 | 39.3 | 17.69 | 10.0 | |
| 0.050 | - | 0.011 | 13.7 | 0.0104 | 4.0 | |
| 0.25 | - | 0.099 | 2.3 | 0.1172 | 8.2 | |
| 0.073 | - | 0.7 | 15.8 | 0.0573 | 6.4 | |
| 1.4 | - | 0.22 | 0.1 | 1.36 | 17.3 | |
| 295 | - | 700 | 10.5 | 94.3 | 14.6 | |
| 0.0081 | - | 0.028 | 21.8 | 0.0779 | 2.1 | |
| 0.57 | - | 1.10 | 3.2 | 1.158 | 1.6 | |
| 63 | - | 1970 | 6.0 | 184 | 7.7 | |
| 16.2 | - | 17.3 | 8.8 | 12.6 | 37.0 | |
| 0.087 | - | 0.33 | 10.1 | 0.72 | 23.8 | |
| 0 | - | 0.14 | 19.2 | 15.1 | 45.9 | |
| - | - | 1.22 | 2.8 | 10.6 | 24.5 | |
| 0.0 | 1.5 | ||
| 0 | 1500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorrini, F.A.; Zamudio Lara, J.M.; Biagiola, S.I.; Figueroa, J.L.; Hernández Escoto, H.; Hantson, A.-L.; Vande Wouwer, A. Experimental Study of Substrate Limitation and Light Acclimation in Cultures of the Microalgae Scenedesmus obliquus—Parameter Identification and Model Predictive Control. Processes 2020, 8, 1551. https://doi.org/10.3390/pr8121551
Gorrini FA, Zamudio Lara JM, Biagiola SI, Figueroa JL, Hernández Escoto H, Hantson A-L, Vande Wouwer A. Experimental Study of Substrate Limitation and Light Acclimation in Cultures of the Microalgae Scenedesmus obliquus—Parameter Identification and Model Predictive Control. Processes. 2020; 8(12):1551. https://doi.org/10.3390/pr8121551
Chicago/Turabian StyleGorrini, Federico Alberto, Jesús Miguel Zamudio Lara, Silvina Inés Biagiola, José Luis Figueroa, Héctor Hernández Escoto, Anne-Lise Hantson, and Alain Vande Wouwer. 2020. "Experimental Study of Substrate Limitation and Light Acclimation in Cultures of the Microalgae Scenedesmus obliquus—Parameter Identification and Model Predictive Control" Processes 8, no. 12: 1551. https://doi.org/10.3390/pr8121551
APA StyleGorrini, F. A., Zamudio Lara, J. M., Biagiola, S. I., Figueroa, J. L., Hernández Escoto, H., Hantson, A. -L., & Vande Wouwer, A. (2020). Experimental Study of Substrate Limitation and Light Acclimation in Cultures of the Microalgae Scenedesmus obliquus—Parameter Identification and Model Predictive Control. Processes, 8(12), 1551. https://doi.org/10.3390/pr8121551