Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Surfactants
2.3. Cultivation Conditions to Assess the Biodegradability of BSA, Saponin and Tween 20 in Scenedesmus obliquus Cultures
2.4. Growth Measurements in Scenedesmus obliquus Cultures
2.5. Gram Staining
2.6. Inverted Optical Microscope
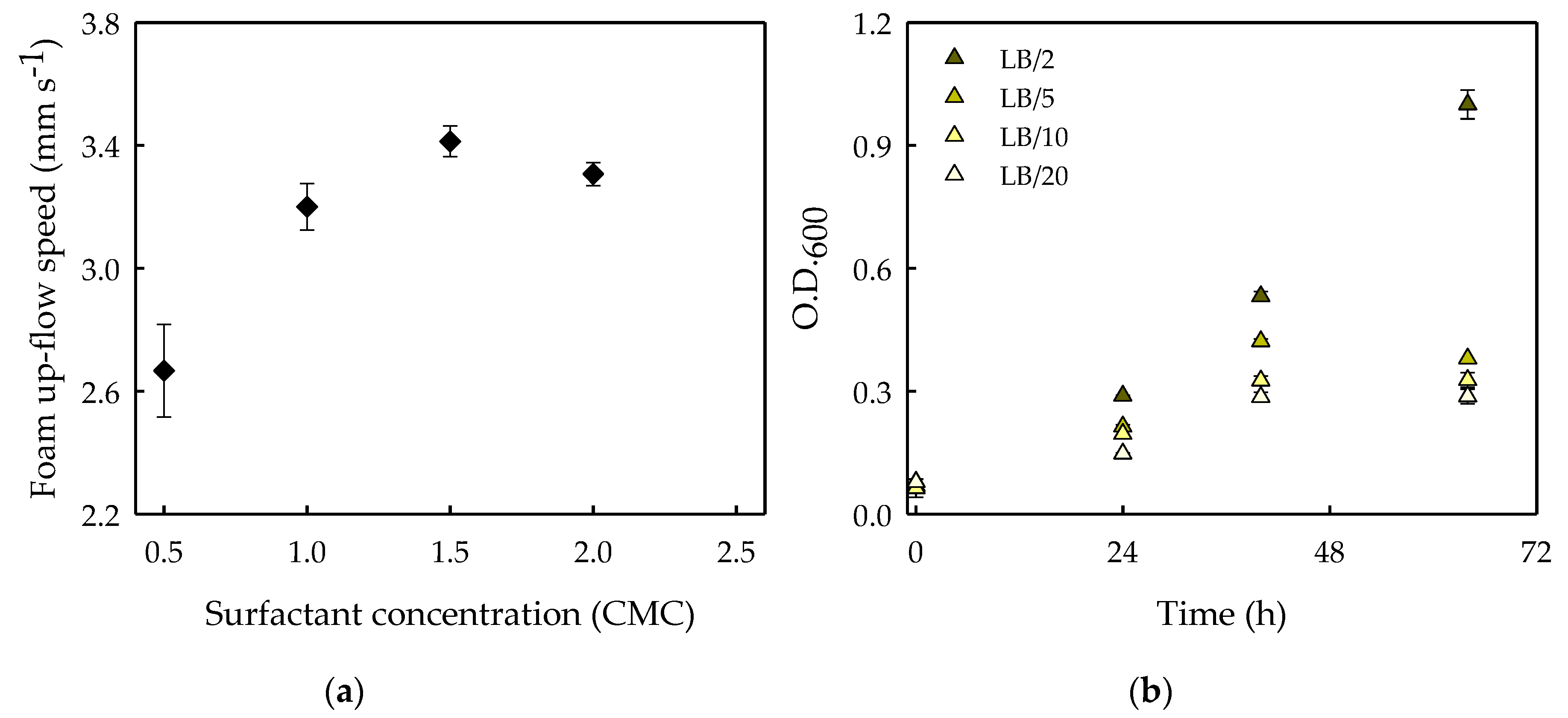
2.7. Assessment of the Effects of Surfactant Biodegradation on Foamability
3. Results and Discussion
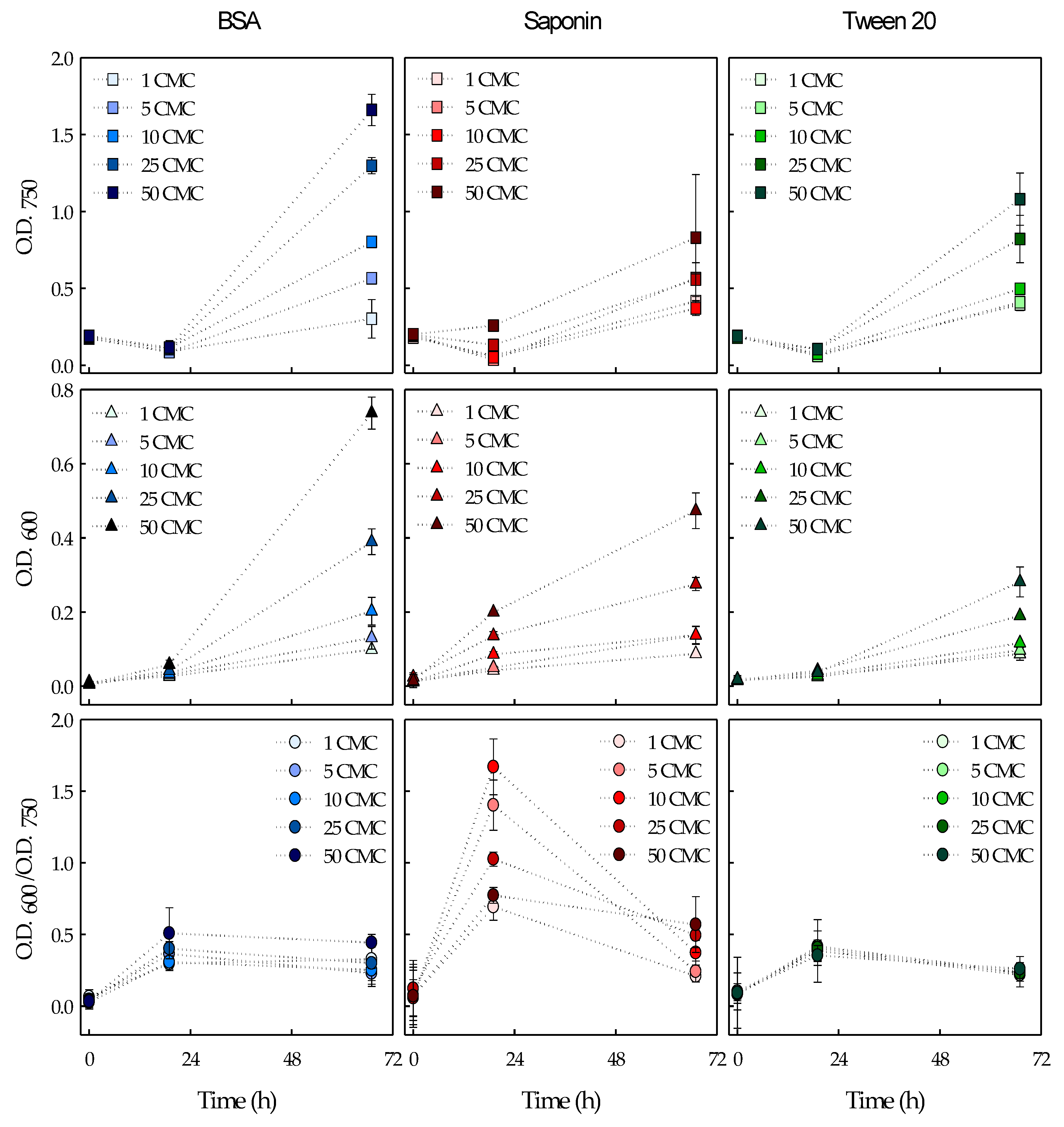
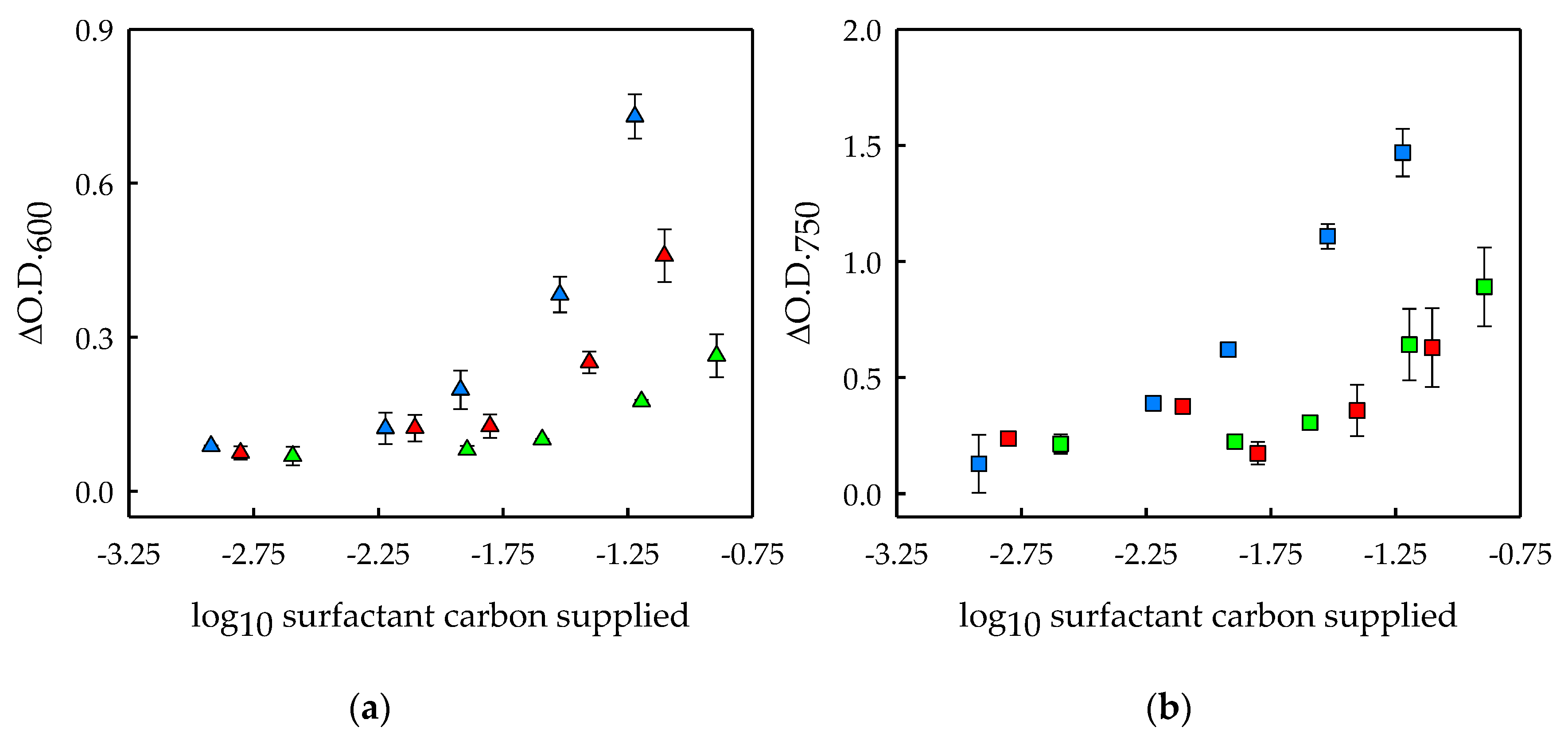
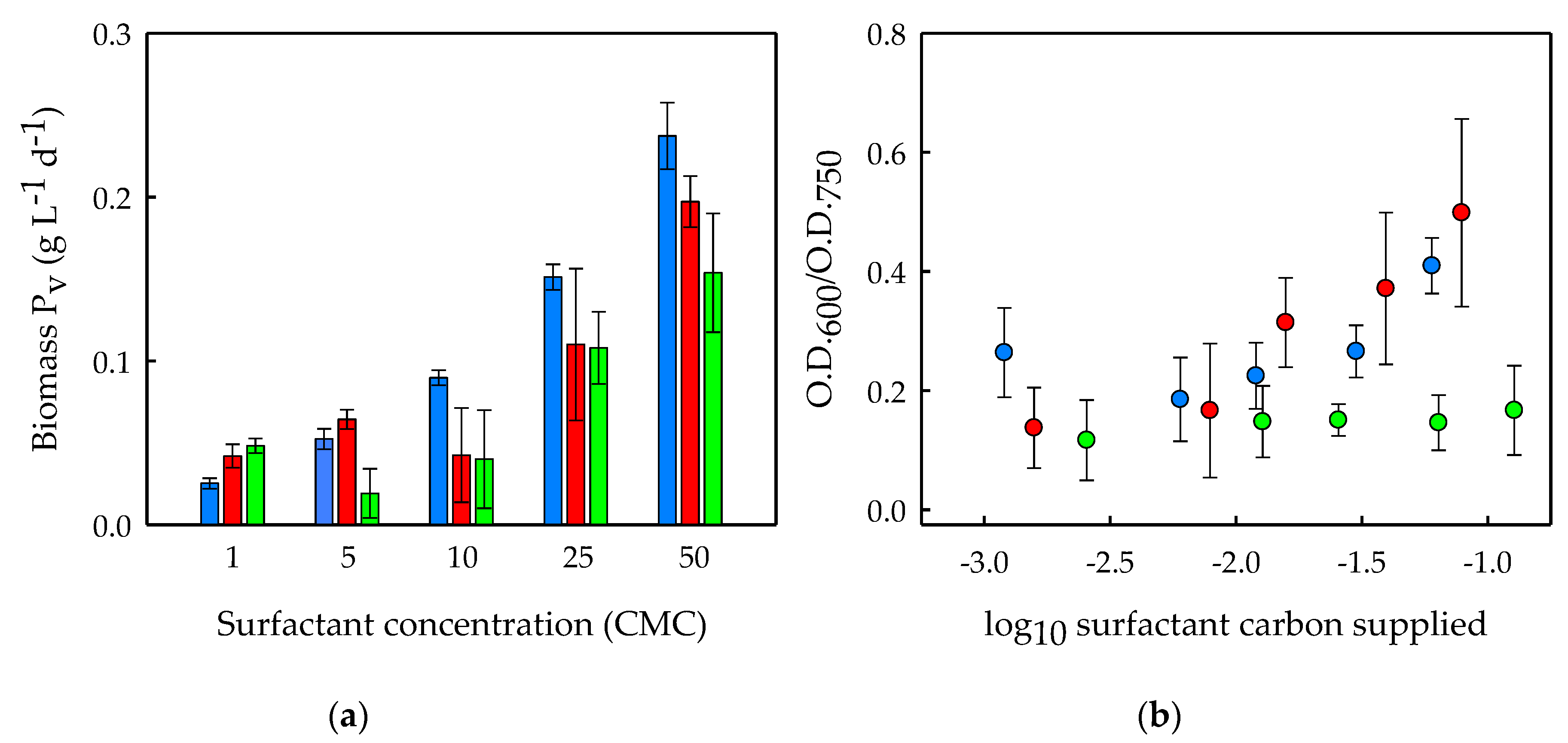
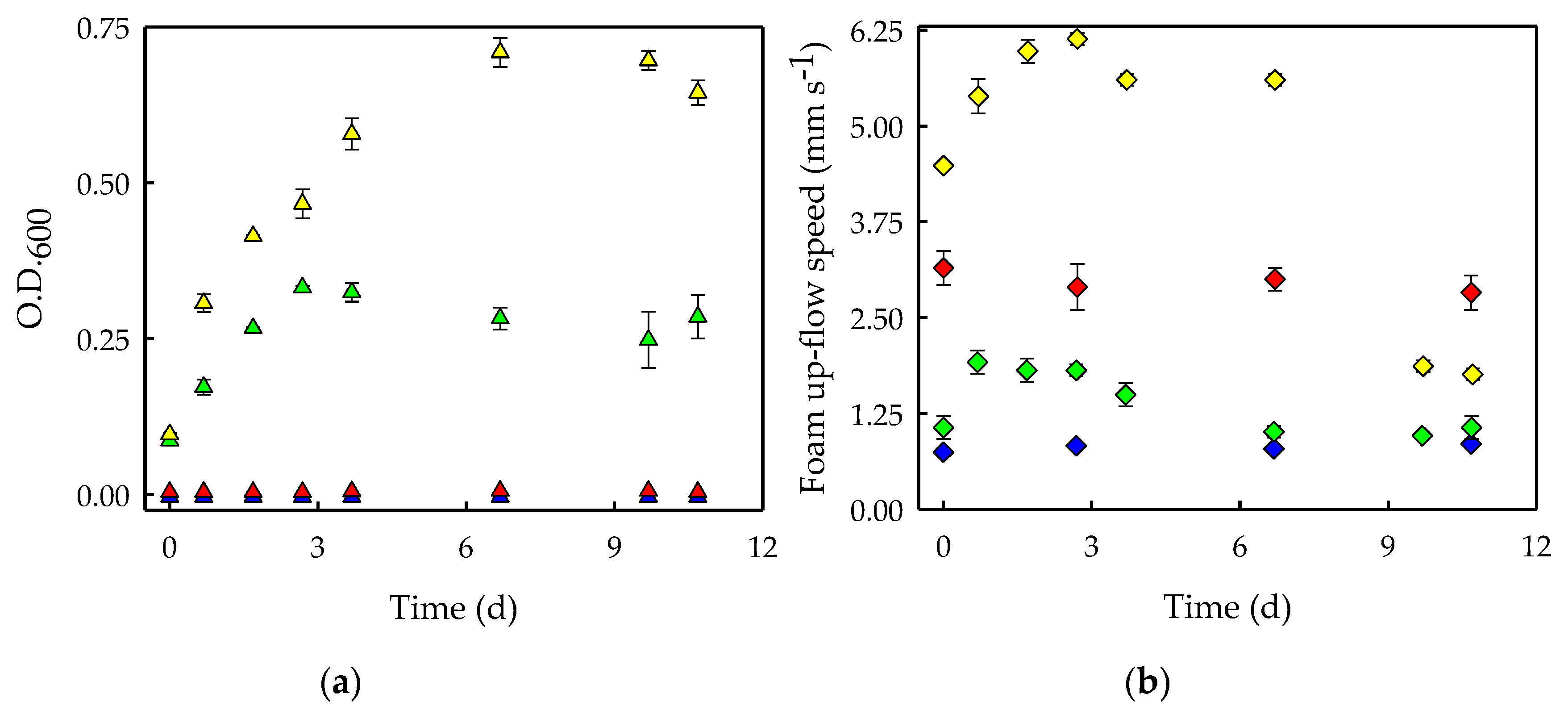
3.1. Surfactants Biodegradation in Microalgal Cultures
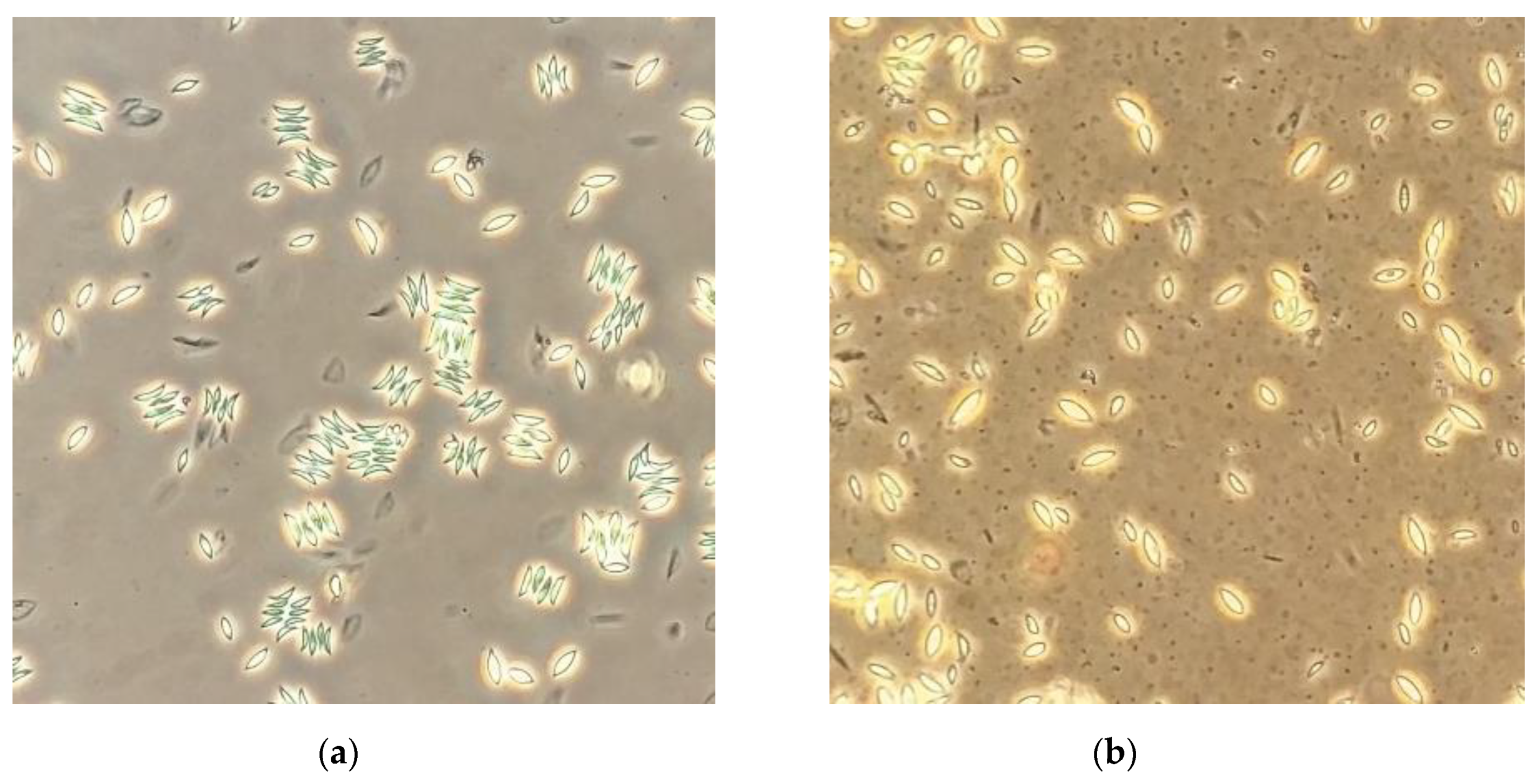
3.2. Morphological Changes in S. obliquus Cultures at High Saponin Concentrations
3.3. Effects of Surfactant Biodegradation on Foaming Capacity
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forján, E.; Navarro, F.; Cuaresma, M.; Vaquero, I.; Ruíz-Domínguez, M.C.; Gojkovic, Ž.; Vázquez, M.; Márquez, M.; Mogedas, B.; Bermejo, E.; et al. Microalgae: Fast-growth sustainable green factories. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1705–1755. [Google Scholar] [CrossRef]
- Ruiz, J.; Olivieri, G.; De Vree, J.; Bosma, R.; Willems, P.; Reith, J.H.; Eppink, M.H.M.; Kleinegris, D.M.M.; Wijffels, R.H.; Barbosa, M.J. Towards industrial products from microalgae. Energy Environ. Sci. 2016, 9, 3036–3043. [Google Scholar] [CrossRef] [Green Version]
- Acién Fernández, F.G.; Fernández Sevilla, J.M.; Molina Grima, E. Photobioreactors for the production of microalgae. Rev. Environ. Sci. Bio/Technology 2013, 12, 131–151. [Google Scholar] [CrossRef]
- Norsker, N.H.; Barbosa, M.J.; Vermuë, M.H.; Wijffels, R.H. Microalgal production—A close look at the economics. Biotechnol. Adv. 2011, 29, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Janoska, A.; Lamers, P.P.; Hamhuis, A.; van Eimeren, Y.; Wijffels, R.H.; Janssen, M. A liquid foam-bed photobioreactor for microalgae production. Chem. Eng. J. 2017, 313, 1206–1214. [Google Scholar] [CrossRef] [Green Version]
- Janoska, A.; Barten, R.; de Nooy, S.; van Rijssel, P.; Wijffels, R.H.; Janssen, M. Improved liquid foam-bed photobioreactor design for microalgae cultivation. Algal Res. 2018, 33, 55–70. [Google Scholar] [CrossRef]
- Janoska, A.; Andriopoulos, V.; Wijffels, R.H.; Janssen, M. Potential of a liquid foam-bed photobioreactor for microalgae cultivation. Algal Res. 2018, 36, 193–208. [Google Scholar] [CrossRef]
- Janoska, A.; Vázquez, M.; Janssen, M.; Wijffels, R.H.; Cuaresma, M.; Vílchez, C. Surfactant selection for a liquid foam-bed photobioreactor. Biotechnol. Prog. 2018, 34, 711–720. [Google Scholar] [CrossRef]
- Ying, G.-G. Distribution, Behavior, Fate, and Effects of Surfactants and Their Degradation Products in the Environment. Environ. Int. 2004, 77–109. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes, J.L.; Garbayo, I.; Cuaresma, M.; Montero, Z.; González-Del-Valle, M.; Vílchez, C. Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar. Drugs 2016, 14, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, Z.; Su, J.; Tian, Y.; Ning, X.; Hong, H.; Zheng, T. Lysis of a red-tide causing alga, Alexandrium tamarense, caused by bacteria from its phycosphere. Biol. Control 2010, 52, 123–130. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Hernandez, J.P.; Morey, T.; Bashan, Y. Microalgae growth-promoting bacteria as “helpers” for microalgae: A novel approach for removing ammonium and phosphorus from municipal wastewater. Water Res. 2004, 38, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Choix, F.J.; de-Bashan, L.E.; Bashan, Y. Enhanced accumulation of starch and total carbohydrates in alginate-immobilized Chlorella spp. induced by Azospirillum brasilense: I. Autotrophic conditions. Enzym. Microb. Technol. 2012, 51, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, M.; Fuentes, J.L.; Hincapié, A.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Selection of microalgae with potential for cultivation in surfactant-stabilized foam. Algal Res. 2018, 31, 216–224. [Google Scholar] [CrossRef]
- Ferreira, A.; Ribeiro, B.; Marques, P.A.S.S.; Ferreira, A.F.; Dias, A.P.; Pinheiro, H.M.; Reis, A.; Gouveia, L. Scenedesmus obliquus mediated brewery wastewater remediation and CO2 biofixation for green energy purposes. J. Clean. Prod. 2017, 165, 1316–1327. [Google Scholar] [CrossRef]
- Wang, R.; Xue, S.; Zhang, D.; Zhang, Q.; Wen, S.; Kong, D.; Yan, C.; Cong, W. Construction and characteristics of artificial consortia of Scenedesmus obliquus-bacteria for S. obliquus growth and lipid production. Algal Res. 2015, 12, 436–445. [Google Scholar] [CrossRef]
- Cuaresma, M.; Janssen, M.; Vílchez, C.; Wijffels, R.H. Productivity of Chlorella sorokiniana in a short light-path (SLP) panel photobioreactor under high irradiance. Biotechnol. Bioeng. 2009, 104, 352–359. [Google Scholar] [CrossRef]
- Clarkson, J.R.; Cui, Z.F.; Darton, R.C. Protein denaturation in foam: II. Surface activity and conformational change. J. Colloid Interface Sci. 1999, 215, 333–338. [Google Scholar] [CrossRef]
- Helenius, A.; McCaslin, D.R.; Fries, E.; Tanford, C. Properties of detergents. Methods Enzymol. 1979, 56, 734–749. [Google Scholar] [CrossRef] [PubMed]
- Gram, C. Ueber die isolirte Farbung der Schizomyceten in Schnitt-und Trockenpraparaten. Fortschr. Med. 1884, 2, 185–189. [Google Scholar]
- Griffiths, M.J.; Garcin, C.; van Hille, R.P.; Harrison, S.T.L. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.A.; Curtis, B.S.; Curtis, W.R. Improving accuracy of cell and chromophore concentration measurements using optical density. BMC Biophys. 2013, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Jurado, E.; Fernndez-Serrano, M.; Ros, F.; Lechug, M. Aerobic Biodegradation of Surfactants. Biodegrad.-Life Sci. 2013. [Google Scholar] [CrossRef] [Green Version]
- Mølgaard, P.; Chihaka, A.; Lemmich, E.; Furu, P.; Windberg, C.; Ingerslev, F.; Halling-Sørensen, B. Biodegradability of the molluscicidal saponins of Phytolacca dodecandra. Regul. Toxicol. Pharmacol. 2000, 32, 248–255. [Google Scholar] [CrossRef]
- Cadoret, A.; Conrad, A.; Block, J.C. Availability of low and high molecular weight substrates to extracellular enzymes in whole and dispersed activated sludges. Enzym. Microb. Technol. 2002, 31, 179–186. [Google Scholar] [CrossRef]
- Confer, D.R.; Logan, B.E. Molecular weight distribution of hydrolysis products during the biodegradation of model macromolecules in suspended and biofilm cultures. II. Dextran and dextrin. Water Res. 1997, 31, 2137–2145. [Google Scholar] [CrossRef]
- Mosquera-Corral, A.; Montràs, A.; Heijnen, J.J.; Van Loosdrecht, M.C.M. Degradation of polymers in a biofilm airlift suspension reactor. Water Res. 2003, 37, 485–492. [Google Scholar] [CrossRef]
- Wang, C.; Lan, C.Q. Effects of shear stress on microalgae—A review. Biotechnol. Adv. 2018, 36, 986–1002. [Google Scholar] [CrossRef]
- Raymond, J.W.; Rogers, T.N.; Shonnard, D.R.; Kline, A.A. A review of structure-based biodegradation estimation methods. J. Hazard. Mater. 2001, 84, 189–215. [Google Scholar] [CrossRef]
- Yeh, D.H.; Pennell, K.D.; Pavlostathis, S.G. Toxicity and biodegradability screening of nonionic surfactants using sediment-derived methanogenic consortia. Water Sci. Technol. 1998, 38, 55–62. [Google Scholar] [CrossRef]
- Chang, Y.C.; Huang, S.C.; Chen, K.F. Evaluation of the effects of nanoscale zero-valent iron (nZVI) dispersants on intrinsic biodegradation of trichloroethylene (TCE). Water Sci. Technol. 2014, 69, 2357–2363. [Google Scholar] [CrossRef] [PubMed]
- Flynn, K.; Butler, I. Nitrogen sources for the growth of marine microalgae: Role of dissolved free amino acids. Mar. Ecol. Prog. Ser. 1986, 34, 281–304. [Google Scholar] [CrossRef]
- Isleten-Hosoglu, M.; Gultepe, I.; Elibol, M. Optimization of carbon and nitrogen sources for biomass and lipid production by Chlorella saccharophila under heterotrophic conditions and development of Nile red fluorescence based method for quantification of its neutral lipid content. Biochem. Eng. J. 2012, 61, 11–19. [Google Scholar] [CrossRef]
- Xiong, W.; Li, X.; Xiang, J.; Wu, Q. High-density fermentation of microalga Chlorella protothecoides in bioreactor for microbio-diesel production. Appl. Microbiol. Biotechnol. 2008, 78, 29–36. [Google Scholar] [CrossRef]
- Tian-Yuan, Z.; Yin-Hu, W.; Lin-Lan, Z.; Xiao-Xiong, W.; Hong-Ying, H. Screening heterotrophic microalgal strains by using the Biolog method for biofuel production from organic wastewater. Algal Res. 2014, 6, 175–179. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Mishra, S. Enhanced biofuel production potential with nutritional stress amelioration through optimization of carbon source and light intensity in Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 179, 565–572. [Google Scholar] [CrossRef]
- Tuchman, N.C.; Schollett, M.A.; Rier, S.T.; Geddes, P. Differential heterotrophic utilization of organic compounds by diatoms and bacteria under light and dark conditions. Hydrobiologia 2006, 561, 167–177. [Google Scholar] [CrossRef]
- Azizullah, A.; Richter, P.; Häder, D.P. Toxicity assessment of a common laundry detergent using the freshwater flagellate Euglena gracilis. Chemosphere 2011, 84, 1392–1400. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, Y.; Huang, W.; Xu, J.; Wang, Z.; Xu, J.; Yuan, Z. Enhanced Accumulation of Carbohydrate and Starch in Chlorella zofingiensis Induced by Nitrogen Starvation. Appl. Biochem. Biotechnol. 2014, 174, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Greses, S.; Christian, J.; Aguado, D.; Ferrer, J.; Seco, A. Microbial community characterization during anaerobic digestion of Scenedesmus spp. under mesophilic and thermophilic conditions. Algal Res. 2017, 27, 121–130. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surfactant | Type | CMC (g L−1) | Molecular Weight (g mol−1) | Carbon Content (% w/w) | Chemical Composition |
|---|---|---|---|---|---|
| BSA | Non-catalytic protein, anionic character | 0.03 [20] | 2754.11 | 53.64 | Bovine serum albumin derived from cows |
| Saponin | Non-ionic | 0.05 (P.I.) | 486.69 | 42.73 | Glycoside + Triterpene |
| Tween 20 | Non-ionic | 0.06 [21] | 1228.00 | 56.73 | Polyoxyethylene (20) sorbitanmonolaurate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, M.; Castilla-Alcántara, J.C.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures. Processes 2020, 8, 1640. https://doi.org/10.3390/pr8121640
Vázquez M, Castilla-Alcántara JC, Garbayo I, Vílchez C, Cuaresma M. Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures. Processes. 2020; 8(12):1640. https://doi.org/10.3390/pr8121640
Chicago/Turabian StyleVázquez, María, José Carlos Castilla-Alcántara, Inés Garbayo, Carlos Vílchez, and María Cuaresma. 2020. "Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures" Processes 8, no. 12: 1640. https://doi.org/10.3390/pr8121640
APA StyleVázquez, M., Castilla-Alcántara, J. C., Garbayo, I., Vílchez, C., & Cuaresma, M. (2020). Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures. Processes, 8(12), 1640. https://doi.org/10.3390/pr8121640