Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions



Abstract
:1. Introduction
2. Industrial Bioprocesses: Current Issues
2.1. Feedstock
2.2. Inhibitory Compounds from the Feedstock
2.3. Changes in the pH Value
2.4. Water Demand
2.5. Metabolic Balance
2.6. Product Inhibition
2.7. Continuous Cultivation of Microorganisms
3. Industrial Bioprocesses: Possible Solutions
3.1. Cell Factories
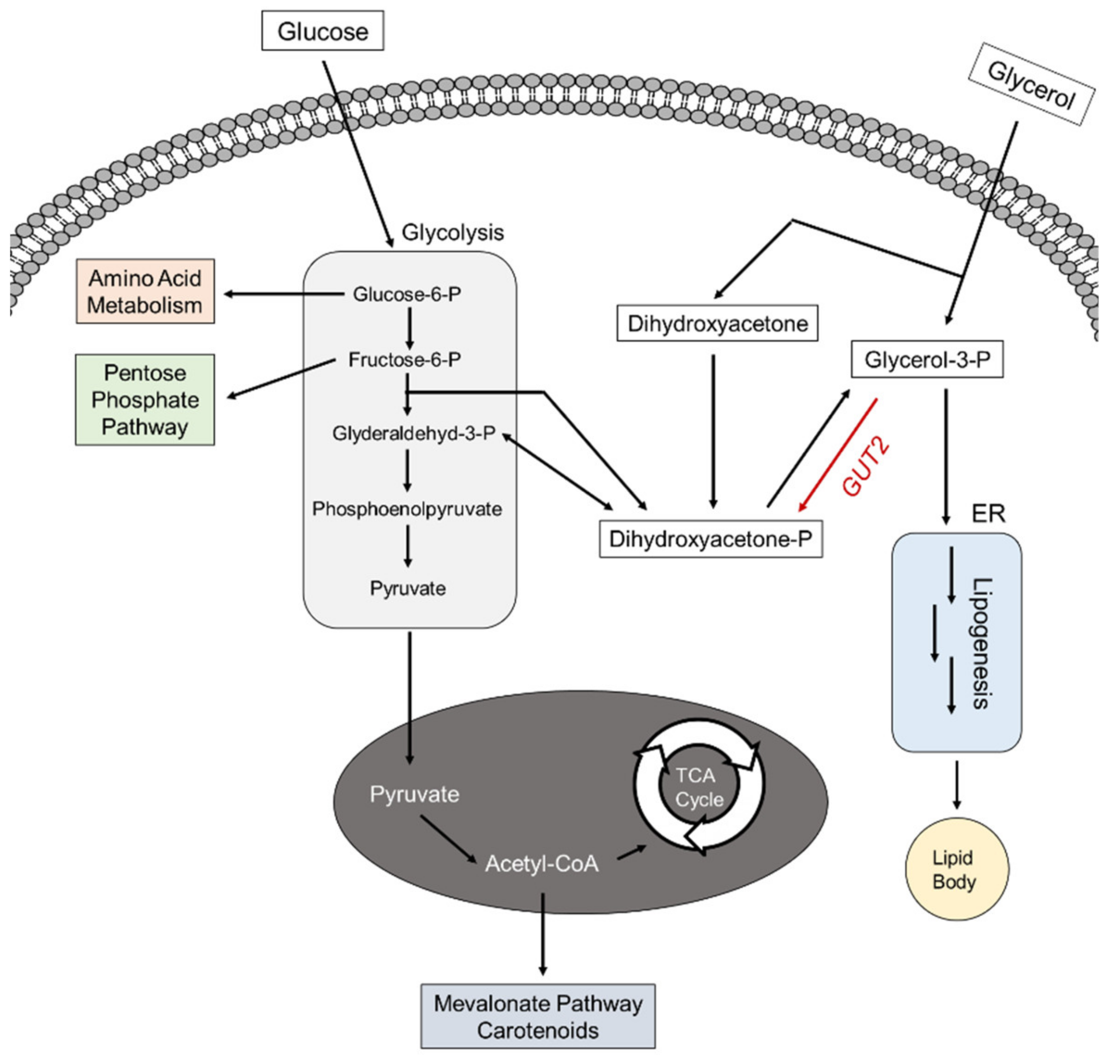
3.1.1. Glycerol as Feedstock: Yarrowia lipolytica
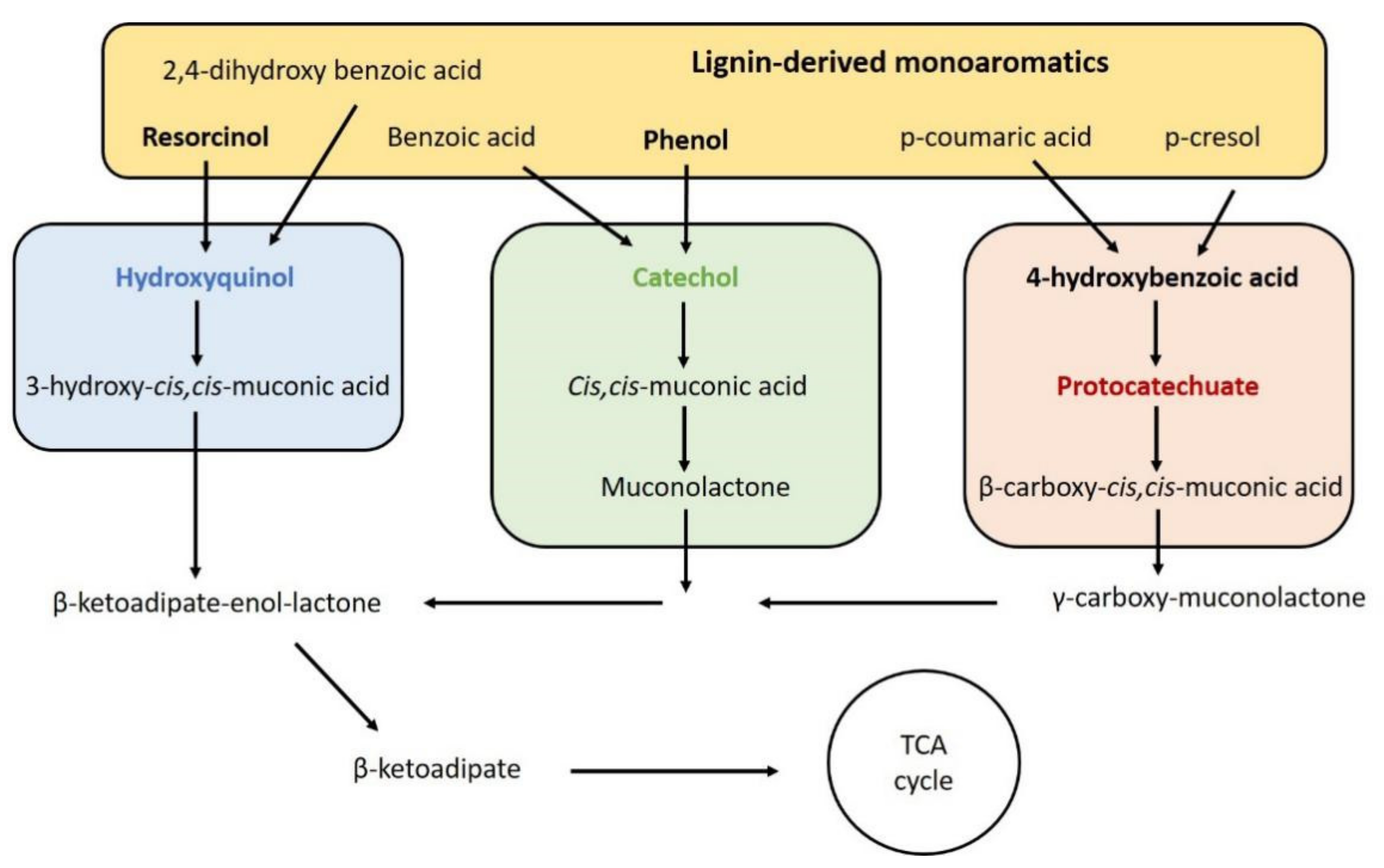
3.1.2. Tolerance and Conversion of Lignocellulosic Inhibitors: Trichosporon oleaginosus
3.1.3. Tolerance to a Broad Range of pH Values: Ustilago cynodontis
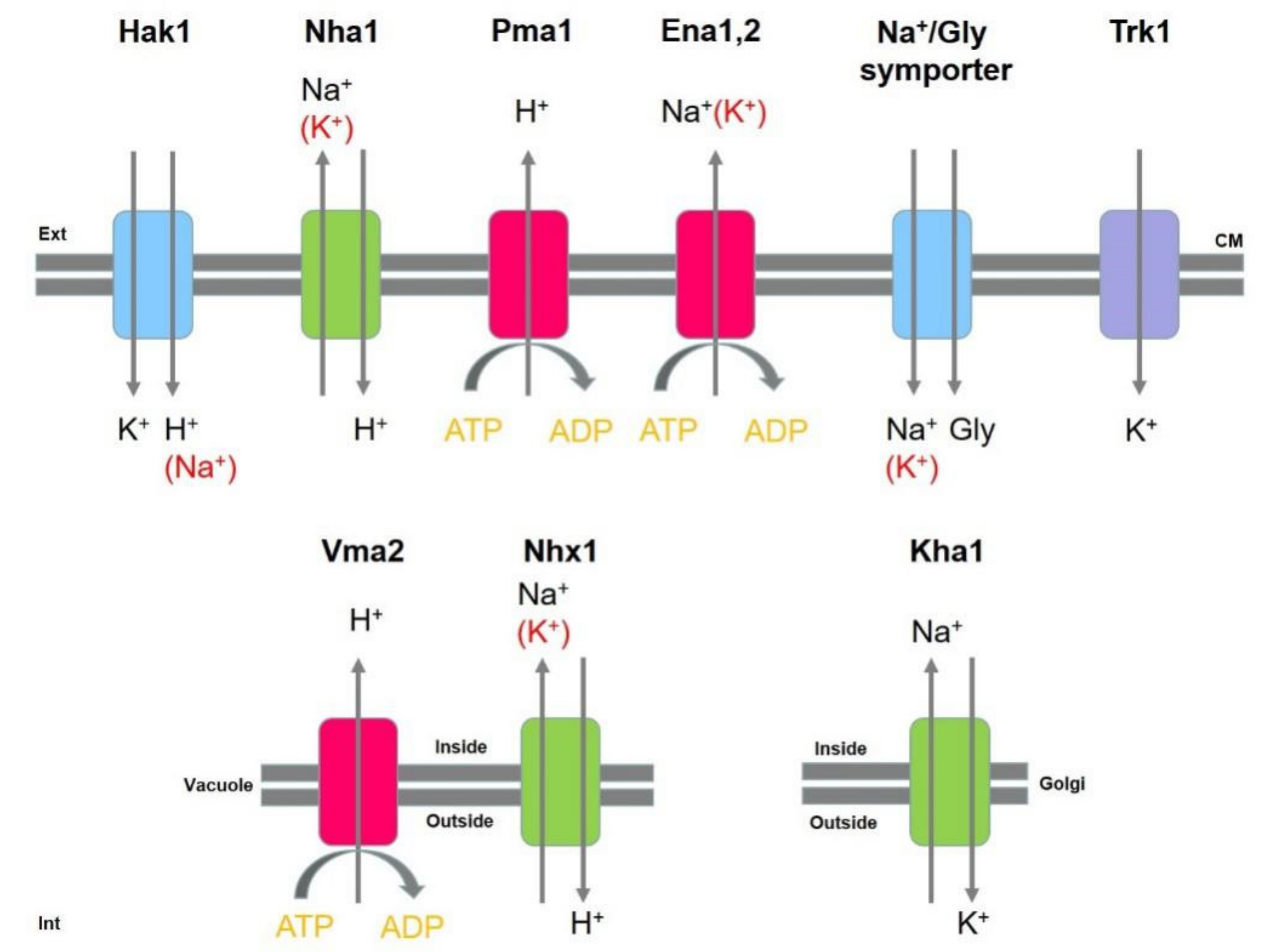
3.1.4. Use of Non-Pure Water: Debaryomyces hansenii
3.2. Bioreactor Configurations
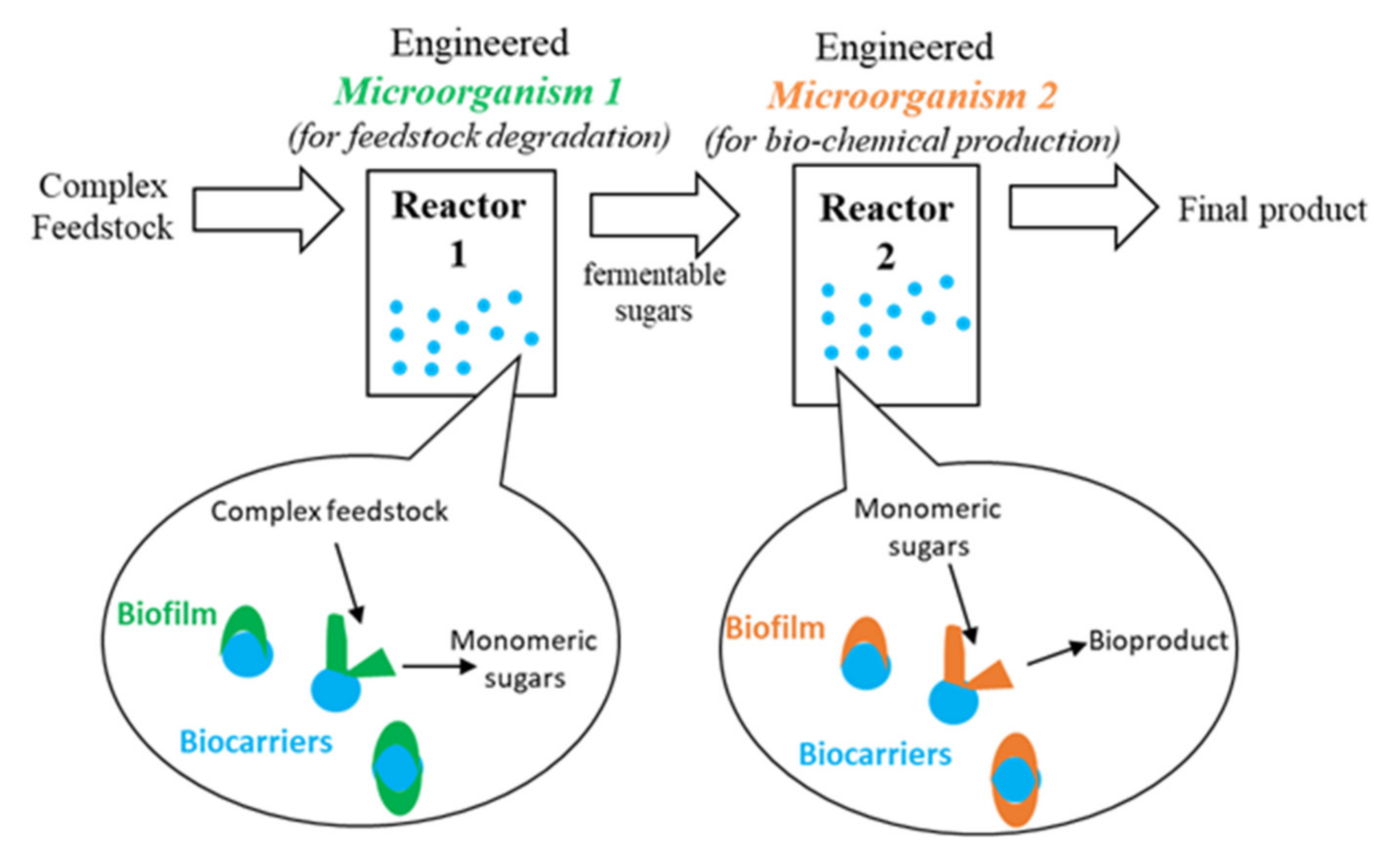
3.2.1. Sequential Bioreactors
- (1)
- Use of a feedstock containing a carbon source more preferable to the microorganism than the hydrolysis product. For example, a feedstock containing or supplemented with crude glycerol could be used for certain microorganisms with a preference for glycerol. Provided an appropriate dilution rate is used, the resulting glucose will not be metabolized in reactor 1 and hence will be transferred to reactor 2 for use in product formation.
- (2)
- Through careful characterization of substrate consumption and cellulose hydrolysis kinetics in the first reactor, the dilution rate can be optimized such that recovery of the sugar product is maximized while ensuring minimal carbon source availability for substrate degradation by the first microorganism.
3.2.2. Integrated Downstream Process Units
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cherubini, F.; Jungmeier, G.; Wellisch, M.; Willke, T.; Skiadas, I.V.; Van Ree, R.; De Jong, E. Toward a common classification approach for biorefinery systems. Biofuels Bioprod. Biorefining 2009, 3, 534–546. [Google Scholar] [CrossRef]
- Hingsamer, M.; Jungmeier, G. Biorefineries. In The Role of Bioenergy in the Emerging Bioeconomy; Lago, C., Caldes, N., Lecon, Y., Eds.; Elsevier Inc.: Madrid, Spain, 2019; pp. 179–222. [Google Scholar]
- Caullet, C.; Le Notre, J. Vegetable oil Biorefineries. In Industrial Biorefinery & White Biotechnology; Pandey, A., Hofer, R., Larroche, C., Eds.; Elsevier Inc.: London, UK, 2015; pp. 247–270. [Google Scholar]
- Waites, M.J.; Morgan, N.L.; Rockey, J.S.; Higton, G. Industrial Microbiology: An Introduction; Blackwell Science Ltd.: Oxford, UK, 2001. [Google Scholar]
- Novo Nordisk. Available online: https://www.novonordisk.com/ (accessed on 14 April 2020).
- Kjeldsen, T.; Balschmidt, P.; Diers, I.; Hach, M.; Kaarsholm, N.C.; Ludvigsen, S. Expression of insulin in yeast: The importance of molecular adaptation for secretion and conversion. Biotechnol. Genet. Eng. Rev. 2001, 18, 89–121. [Google Scholar] [CrossRef] [PubMed]
- Biosynthia. Available online: https://www.biosyntia.com/ (accessed on 14 April 2020).
- Novozymes. Available online: https://www.novozymes.com/en (accessed on 14 April 2020).
- LanzaTech. Available online: https://www.lanzatech.com/ (accessed on 14 April 2020).
- Marcellin, E.; Behrendorff, J.B.Y.H.; Nagaraju, S.; DeTissera, S.; Segovia, S.; Palfreyman, R.W.; Daniell, J.; Licona-Cassani, C.; Quek, L.-E.; Speight, R.; et al. Low carbon fuels and commodity chemicals from waste gases – systematic approach to understand energy metabolism in a model acetogen. Green Chem. 2016, 18, 3020–3028. [Google Scholar] [CrossRef] [Green Version]
- DSM. Available online: https://www.dsm.com/corporate/home.html (accessed on 15 April 2020).
- Cargill. Available online: https://www.cargill.com/home (accessed on 15 April 2020).
- Lonza. Available online: https://www.lonza.com/ (accessed on 15 April 2020).
- Workman, M.; Andersen, M.R.; Thykaer, J. Integrated Approaches for Assessment of Cellular Performance in Industrially Relevant Filamentous Fungi. Ind. Biotechnol. 2013, 9, 337–344. [Google Scholar] [CrossRef]
- Klímek, P.; Wimmer, R.; Mishra, P.K.; Kúdela, J. Utilizing brewer’s-spent-grain in wood-based particleboard manufacturing. J. Clean. Prod. 2017, 141, 812–817. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; Rehmann, L.; Marzocchella, A. Deep Eutectic Solvents pretreatment of agro-industrial food waste. Biotechnol. Biofuels 2018, 11, 37. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; De La Feld, M.; Marzocchella, A. Renewable feedstocks for biobutanol production by fermentation. New Biotechnol. 2017, 39, 135–140. [Google Scholar] [CrossRef]
- Berlowska, J.; Cieciura-Wloch, W.; Kalinowska, H.; Kręgiel, D.; Borowski, S.; Pawlikowska, E.; Binczarski, M.; Witonska, I.A. Enzymatic Conversion of Sugar Beet Pulp: A Comparison of Simultaneous Saccharification and Fermentation and Separate Hydrolysis and Fermentation for Lactic Acid Production. Food Technol. Biotechnol. 2018, 56, 188–196. [Google Scholar] [CrossRef]
- Prasad, S.; Singh, A.; Joshi, H. Ethanol as an alternative fuel from agricultural, industrial and urban residues. Resour. Conserv. Recycl. 2007, 50, 1–39. [Google Scholar] [CrossRef]
- Ameram, N.; Muhammad, S.; Yusof, N.A.A.N.; Ishak, S.; Ali, A.; Shoparwe, N.F.; Ter, T.P. Chemical composition in sugarcane bagasse: Delignification with sodium hydroxide. Malays. J. Fundam. Appl. Sci. 2019, 15, 232–236. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; De La Feld, M.; Marzocchella, A. Agro Food Wastes and Innovative Pretreatments to Meet Biofuel Demand in Europe. Chem. Eng. Technol. 2019, 42, 954–961. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; Marzocchella, A. Pre-treatment and enzymatic hydrolysis of lettuce residues as feedstock for bio-butanol production. Biomass- Bioenergy 2017, 96, 172–179. [Google Scholar] [CrossRef]
- RTaherzadeh, M.; Bolton, K.; Wong, J.; Pandey, A. Sustainable Resource Recovery and Zero Waste Approaches; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Rajendran, K.; Lin, R.; Wall, D.M.; Murphy, J.D. Influential Aspects in Waste Management Practices. Sustainable Resource Recovery and Zero Waste Approaches 2019, 65–78. [Google Scholar] [CrossRef]
- McCoy, M. GLYCERIN SURPLUS. Chem. Eng. News Arch. 2006, 84, 7. [Google Scholar] [CrossRef]
- Rodrigues, A.; Bordado, J.; Dos Santos, R.G. Upgrading the Glycerol from Biodiesel Production as a Source of Energy Carriers and Chemicals—A Technological Review for Three Chemical Pathways. Energies 2017, 10, 1817. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Barrett, D.M.; Delwiche, M.J.; Stroeve, P. Methods for Pretreatment of Lignocellulosic Biomass for Efficient Hydrolysis and Biofuel Production. Ind. Eng. Chem. Res. 2009, 48, 3713–3729. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, Q.; Cheng, G. Deconstruction of corncob by steam explosion pretreatment: Correlations between sugar conversion and recalcitrant structures. Carbohydr. Polym. 2017, 156, 351–356. [Google Scholar] [CrossRef]
- Kim, D. Physico-Chemical Conversion of Lignocellulose: Inhibitor Effects and Detoxification Strategies: A Mini Review. Molecul 2018, 23, 309. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Franden, M.A.; Yang, Q.; Chou, Y.-C.; Zhang, M.; Pienkos, P.T. Identification of Inhibitors in Lignocellulosic Slurries and Determination of Their Effect on Hydrocarbon-Producing Microorganisms. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Kucharska, K.; Rybarczyk, P.; Hołowacz, I.; Łukajtis, R.; Glinka, M.; Kamiński, M. Pretreatment of Lignocellulosic Materials as Substrates for Fermentation Processes. Mol. 2018, 23, 2937. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Bao, J. Lignocellulose Pretreatment Using Acid as Catalyst. In Handbook of Biorefinery Research and Technology; Park, J., Ed.; Springer: Dordrecht, The Netherlands, 2018. [Google Scholar]
- Chin, J.Y.; Ab Rahim, S.K.E.; Abdullah, N.S. Studies on Alkaline Pretreatment of Sugarcane Bagasse and Rice Straw Hydrolysis for the Recovery of Reducing Sugar. Int. J. Curr. Res. Sci. Eng. Technol. 2018, 1, 26–31. [Google Scholar] [CrossRef]
- Novozymes. Bioenergy Application sheet Novozyme Cellic CTec3 HS. Available online: https://www.novozymes.com/en (accessed on 14 April 2020).
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar] [CrossRef]
- Hetényi, K.; Nemeth, A.; Sevella, B. Role of pH-regulation in lactic acid fermentation: Second steps in a process improvement. Chem. Eng. Process. Process. Intensif. 2011, 50, 293–299. [Google Scholar] [CrossRef]
- Emmenegger, M.F.; Pfister, S.; Koehler, A.; De Giovanetti, L.; Arena, A.P.; Zah, R. Taking into account water use impacts in the LCA of biofuels: An Argentinean case study. Int. J. Life Cycle Assess. 2011, 16, 869–877. [Google Scholar] [CrossRef]
- Chiu, Y.-W.; Suh, S.; Pfister, S.; Hellweg, S.; Koehler, A. Measuring ecological impact of water consumption by bioethanol using life cycle impact assessment. Int. J. Life Cycle Assess. 2011, 17, 16–24. [Google Scholar] [CrossRef]
- Hossain, G.; Liu, L.; Du, G. Industrial Bioprocesses and the Biorefinery Concept. Current Developments in Biotechnology and Bioengineering 2017, 3–27. [Google Scholar] [CrossRef]
- Yang, M.; Wang, J.; Nan, Y.; Zhang, J.; Li, L.; Liu, G.; Vepsäläinen, J.; Kuittinen, S.; Pappinen, A. Effect of salts formed by neutralization for the enzymatic hydrolysis of cellulose and acetone–butanol–ethanol fermentation. RSC Adv. 2019, 9, 33755–33760. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, C.; Frost, A.T.; Ramos-Moreno, L.; Martinez, J.L. A Physiological Characterization in Controlled Bioreactors Reveals a Novel Survival Strategy for Debaryomyces hansenii at High Salinity and Confirms its Halophilic Behavior. Biorxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Conway, P.M.; Cervenka, N.D.; Cui, J.; Maloney, M.; Olson, D.G.; Lynd, L.R. Metabolic engineering of Clostridium thermocellum for n-butanol production from cellulose. Biotechnol. Biofuels 2019, 12, 186. [Google Scholar] [CrossRef]
- Song, X.; Li, Y.; Wu, Y.; Cai, M.; Liu, Q.; Gao, K.; Zhang, X.; Bai, Y.; Xu, H.; Qiao, M. Metabolic engineering strategies for improvement of ethanol production in cellulolytic Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18, 18–26. [Google Scholar] [CrossRef]
- Kumar, R.; Tabatabaei, M.; Karimi, K.; Horváth, I.S. Recent updates on lignocellulosic biomass derived ethanol—A review. Biofuel Res. J. 2016, 3, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Shih-Hsin, H.; Kengo, S.; Riaan den, H.; Kentaro, I.; Chiaki, O.; Willem, H.Z.; Tomohisa, H. Engineering of a novel cellulose-adherent cellulolytic Saccharomyces cerevisiae for cellulolitic biofuel production. Sci. Rep. 2016, 6, 1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wu, D.; Lin, Y.; Wang, X.; Kong, H.; Tanaka, S. Substrate and Product Inhibition on Yeast Performance in Ethanol Fermentation. Energy Fuels 2015, 29, 1019–1027. [Google Scholar] [CrossRef]
- Wen, Z.; Ledesma-Amaro, R.; Lin, J.; Jiang, Y.; Yang, S. Improved n-Butanol Production from Clostridium cellulovorans by Integrated Metabolic and Evolutionary Engineering. Appl. Environ. Microbiol. 2019, 85, 7–24. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Yun, J.; Zhang, H.; Magocha, T.A.; Zabed, H.M.; Xue, Y.; Fokum, E.B.; Sun, W.; Qi, X. Genetically Engineered Strains: Application and Advances for 1,3-Propanediol Production from Glycerol. Food Technol. Biotechnol. 2017, 56, 3–15. [Google Scholar] [CrossRef]
- Mezzina, M.P.; Álvarez, D.S.; Egoburo, D.E.; Peña, R.D.; Nikel, P.I.; Pettinari, M.J. A New Player in the Biorefineries Field: Phasin PhaP Enhances Tolerance to Solvents and Boosts Ethanol and 1,3-Propanediol Synthesis in Escherichia coli. Appl. Environ. Microbiol. 2017, 83, e00662-17. [Google Scholar] [CrossRef] [Green Version]
- Davy, A.M.; Kildegaard, H.F.; Andersen, M.R. Cell Factory Engineering. Cell Syst. 2017, 4, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Karagiosis, S.A.; Baker, S.E. Fungal Cell Factories. Food and Industrial Bioproducts and Bioprocessing 2012, 205–219. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Bailey, J. Toward a science of metabolic engineering. Science 1991, 252, 1668–1675. [Google Scholar] [CrossRef]
- Baeshen, N.A.; Baeshen, M.N.; Sheikh, A.; Bora, R.S.; Ahmed, M.M.M.; Ramadan, H.A.; Saini, K.S.; Redwan, E.M. Cell factories for insulin production. Microb. Cell Factories 2014, 13, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomgardner, M. Cargill, DSM start up stevia factory. C&EN Glob. Enterp. 2019, 97, 14–15. [Google Scholar] [CrossRef]
- Wickerham, L.J.; Kurtzman, C.P.; Herman, A.I. Sexual Reproduction in Candida lipolytica. Am. Assoc. Adv. Sci. 1970, 167, 1141. [Google Scholar] [CrossRef] [PubMed]
- Timoumi, A.; Cléret, M.; Bideaux, C.; Guillouet, S.E.; Allouche, Y.; Molina-Jouve, C.; Fillaudeau, L.; Gorret, N. Dynamic behavior of Yarrowia lipolytica in response to pH perturbations: Dependence of the stress response on the culture mode. Appl. Microbiol. Biotechnol. 2016, 101, 351–366. [Google Scholar] [CrossRef]
- Workman, M.; Holt, P.; Thykaer, J. Comparing cellular performance of Yarrowia lipolytica during growth on glucose and glycerol in submerged cultivations. AMB Express 2013, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Beopoulos, A.; Mrozova, Z.; Thevenieau, F.; Le Dall, M.-T.; Hapala, I.; Papanikolaou, S.; Chardot, T.; Nicaud, J.-M. Control of Lipid Accumulation in the Yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 7779–7789. [Google Scholar] [CrossRef] [Green Version]
- Larroude, M.; Celinska, E.; Back, A.; Thomas, S.; Nicaud, J.-M.; Ledesma-Amaro, R. A synthetic biology approach to transform Yarrowia lipolytica into a competitive biotechnological producer of β-carotene. Biotechnol. Bioeng. 2017, 115, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.; Jackson, E.N.; Zhu, Q. Sustainable source of omega-3 eicosapentaenoic acid from metabolically engineered Yarrowia lipolytica: From fundamental research to commercial production. Appl. Microbiol. Biotechnol. 2015, 99, 1061–1062. [Google Scholar] [CrossRef] [Green Version]
- Juszczyk, P.; Rymowicz, W.; Kita, A.; Rywińska, A. Biomass production by Yarrowia lipolytica yeast using waste derived from the production of ethyl esters of polyunsaturated fatty acids of flaxseed oil. Ind. Crop. Prod. 2019, 138, 138. [Google Scholar] [CrossRef]
- Fell, J.W.; Boekhout, T.; Fonseca, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. Int. J. Syst. Evol. Micr. 2002, 50, 1351–1371. [Google Scholar] [CrossRef]
- Gujjari, P.; Coumes, K.; Zhou, J.J.; Suh, S.-O. Characterization of oleaginous yeasts revealed two novel species: Trichosporon cacaoliposimilis sp. nov. and Trichosporon oleaginosus sp. nov. Mycologia 2011, 103, 1110–1118. [Google Scholar] [CrossRef]
- Yaguchi, A.; Rives, D.; Blenner, M.A. New kids on the block: Emerging oleaginous yeast of biotechnological importance. AIMS Microbiol. 2017, 3, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Moon, N.J.; Hammond, E.; Glatz, B.A. Conversion of Cheese Whey and Whey Permeate to Oil and Single-Cell Protein. J. Dairy Sci. 1978, 61, 1537–1547. [Google Scholar] [CrossRef]
- Bednarski, W.; Leman, J.; Tomasik, J. Utilization of beet molasses and whey for fat biosynthesis by a yeast. Agric. Wastes 1986, 18, 19–26. [Google Scholar] [CrossRef]
- Christophe, G.; Deo, J.L.; Kumar, V.; Nouaille, R.; Fontanille, P.; Larroche, C. Production of Oils from Acetic Acid by the Oleaginous Yeast Cryptococcus curvatus. Appl. Biochem. Biotechnol. 2011, 167, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zheng, Y.; Dorgan, K.M.; Chen, S. Oil production by oleaginous yeasts using the hydrolysate from pretreatment of wheat straw with dilute sulfuric acid. Bioresour. Technol. 2011, 102, 6134–6140. [Google Scholar] [CrossRef]
- Zheng, Y.; Chi, Z.; Ahring, B.K.; Chen, S. Oleaginous yeast Cryptococcus curvatus for biofuel production: Ammonia’s effect. Biomass- Bioenergy 2012, 37, 114–121. [Google Scholar] [CrossRef]
- Park, G.W.; Chang, H.N.; Jung, K.; Seo, C.; Kim, Y.-C.; Choi, J.H.; Woo, H.C.; Hwang, I.-J. Production of microbial lipid by Cryptococcus curvatus on rice straw hydrolysates. Process. Biochem. 2017, 56, 147–153. [Google Scholar] [CrossRef]
- Yaguchi, A.; Robinson, A.; Mihealsick, E.; Blenner, M.A. Metabolism of aromatics by Trichosporon oleaginosus while remaining oleaginous. Microb. Cell Factories 2017, 16, 206. [Google Scholar] [CrossRef] [Green Version]
- Xiaolei, Z.; Jiaxin, C.; Di, W.; Ji, L.; Rajeshwar, D.T.; Rao, Y.S. Economical lipid production from Trichosporon oleaginosus via dissolved oxygen adjustment and crude glycerol addition. Bioresour. Technol. 2019, 273, 288–296. [Google Scholar]
- Chen, J.; Zhang, X.; Drogui, P.; Tyagi, R. The pH-based fed-batch for lipid production from Trichosporon oleaginosus with crude glycerol. Bioresour. Technol. 2018, 259, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Feldbrügge, M.; Kellner, R.; Schipper, K. The biotechnological use and potential of plant pathogenic smut fungi. Appl. Microbiol. Biotechnol. 2013, 97, 3253–3265. [Google Scholar] [CrossRef] [PubMed]
- Guevarra, E.D.; Tabuchi, T. Accumulation of itaconic, 2-hydroxyparaconic, itatartaric, and malic acids by strains of the genus Ustilago. Agric. Biol. Chem. 1990, 54, 2353–2358. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Konishi, M.; Fukuoka, T.; Imura, T.; Kitamoto, D. Identification of Ustilago cynodontis as a new producer of glycolipid biosurfactants, mannosylerythritol lipids, based on ribosomal DNA sequences. J. Oleo Sci. 2008, 57, 549–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, S. Biological amphiphiles (microbial surfactants). Curr. Opin. Colloid. Interface Sci. 2002, 7, 12–20. [Google Scholar] [CrossRef]
- Kitamoto, D.; Isoda, H.; Nakahara, T. Functional and potential application of glycolipids biosurfactants. J. Biosci. Bioeng. 2002, 94, 187–201. [Google Scholar] [CrossRef]
- Couturier, M.; Navarro, D.; Olivé, C.; Chevret, D.; Haon, M.; Favel, A.; Lesage-Meessen, L.; Henrissat, B.; Coutinho, P.M.; Berrin, J.-G. Post-genomic analyses of fungal lignocellulosic biomass degradation reveal the unexpected potential of the plant pathogen Ustilago maydis. BMC Genom. 2012, 13, 57. [Google Scholar] [CrossRef] [Green Version]
- Geiser, E.; Wierckx, N.; Zimmermann, M.; Blank, L.M. Identification of an endo-1,4-beta-xylanase of Ustilago maydis. BMC Biotechnol. 2013, 13, 59. [Google Scholar] [CrossRef] [Green Version]
- Willke, T.; Vorlop, K.-D. Biotechnological production of itaconic acid. Appl. Microbiol. Biotechnol. 2001, 56, 289–295. [Google Scholar] [CrossRef]
- Geilen, F.M.A.; Engendahl, B.; Harwardt, A.; Marquardt, W.; Klankermayer, J.; Leitner, W. Selective and Flexible Transformation of Biomass-Derived Platform Chemicals by a Multifunctional Catalytic System. Angew. Chem. 2010, 122, 5642–5646. [Google Scholar] [CrossRef]
- Klement, T.; Büchs, J. Itaconic acid – A biotechnological process in change. Bioresour. Technol. 2013, 135, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Tehrani, H.H.; Saur, K.; Tharmasothirajan, A.; Blank, L.M.; Wierckx, N. Process engineering of pH tolerant Ustilago cynodontis for efficient itaconic acid production. Microb. Cell Factories 2019, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tehrani, H.H.; Tharmasothirajan, A.; Track, E.; Blank, L.M.; Wierckx, N. Engineering the morphology and metabolism of pH tolerant Ustilago cynodontis for efficient itaconic acid production. Metab. Eng. 2019, 54, 293–300. [Google Scholar] [CrossRef]
- Norkrans, B. Studies on marine occurring yeasts: Growth related to pH, NaCl concentration and temperature. Arch. Microbiol. 1966, 54, 374–392. [Google Scholar] [CrossRef]
- Breuer, U.; Harms, H. Debaryomyces hansenii—An extremophilic yeast with biotechnological potential. Yeast 2006, 23, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Almagro, A.; Prista, C.; Castro, S.; Quintas, C.; Madeira-Lopes, A.; Ramos, J.; Loureiro-Dias, M. Effects of salts on Debaryomyces hansenii and Saccharomyces cerevisiae under stress conditions. Int. J. Food Microbiol. 2000, 56, 191–197. [Google Scholar] [CrossRef]
- Papouskova, K.; Sychrová, H. The co-action of osmotic and high temperature stresses results in a growth improvement of Debaryomyces hansenii cells. Int. J. Food Microbiol. 2007, 118, 1–7. [Google Scholar] [CrossRef]
- Navarrete, C.; Siles, A.; Martãnez, J.L.; Calero, F.; Ramos, J.; Martãnez, J.L.; Ramos, J. Oxidative stress sensitivity inDebaryomyces hansenii. FEMS Yeast Res. 2009, 9, 582–590. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, L.; Norkrans, B. On the mechanism of salt tolerance. Production of glycerol and heat during growth of Debaryomyces hansenii. Arch Microbiol 1976, 110, 177–183. [Google Scholar] [CrossRef]
- Adler, L.; Blomberg, A.; Nilsson, A. Glycerol metabolism and osmoregulation in the salt-tolerant yeast Debaryomyces hansenii. J. Bacteriol. 1985, 162, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadanho, M.; Almeida, J.; Sampaio, J. Assessment of yeast diversity in a marine environment in the south of Portugal by microsatellite-primed PCR. Antonie van Leeuwenhoek 2003, 84, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Butinar, L.; Zalar, P.; Frisvad, J.C.; Gunde-Cimerman, N. The genus Eurotium: Members of indigenous fungal community in hypersaline waters of salterns. FEMS Microbiol. Ecol. 2005, 51, 155–166. [Google Scholar] [CrossRef]
- Ramos, J.; Melero, Y.; Ramos-Moreno, L.; Michán, C.; Cabezas, L. Debaryomyces hansenii Strains from Valle De Los Pedroches Iberian Dry Meat Products: Isolation, Identification, Characterization, and Selection for Starter Cultures. J. Microbiol. Biotechnol. 2017, 27, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, M.L.D.C.; Delile, A.; Spinnler, H.-E.; Bonnarme, P.; Landaud, S. Comparison of volatile sulphur compound production by cheese-ripening yeasts from methionine and methionine–cysteine mixtures. Appl. Microbiol. Biotechnol. 2007, 75, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Cano-García, L.; Belloch, C.; Flores, M. Impact of Debaryomyces hansenii strains inoculation on the quality of slow dry-cured fermented sausages. Meat Sci. 2014, 96, 1469–1477. [Google Scholar] [CrossRef] [Green Version]
- Prista, C.; Loureiro-Dias, M.; Montiel, V.; García, R.; Ramos, J. Mechanisms underlying the halotolerant way of Debaryomyces hansenii. FEMS Yeast Res. 2005, 5, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Velkova, K.; Sychrová, H. The Debaryomyces hansenii NHA1 gene encodes a plasma membrane alkali-metal-cation antiporter with broad substrate specificity. Gene 2006, 369, 27–34. [Google Scholar] [CrossRef]
- Almagro, A.; Prista, C.; Benito, B.; Loureiro-Dias, M.C.; Ramos, J. Cloning and expression of two genes coding for sodium pumps in the sal-tolerant yeast Debaryomyces hansenii. J. Bacteriol. 2001, 183, 3251–3255. [Google Scholar] [CrossRef] [Green Version]
- Montiel, V.; Ramos, J.; Ramos, J. Intracellular Na+ and K+ distribution in Debaryomyces hansenii. Cloning and expression in Saccharomyces cerevisiae of DhNHX1. FEMS Yeast Res. 2007, 7, 102–109. [Google Scholar] [CrossRef]
- Carcãa-Salcedo, R.; Montiel, V.; Calero, F.; Ramos, J.; Ramos, J.; Salcedo, R.G. Characterization ofDhKHA1, a gene coding for a putative Na+transporter fromDebaryomyces hansenii. FEMS Yeast Res. 2007, 7, 905–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, J.L.; Sychrová, H.; Ramos, J. Monovalent cations regulate expression and activity of the Hak1 potassium transporter in Debaryomyces hansenii. Fungal Genet. Boil. 2011, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Prista, C.; González-Hernández, J.C.; Ramos, J.; Loureiro-Dias, M. Cloning and characterization of two K+ transporters of Debaryomyces hansenii. Microbiol. 2007, 153, 3034–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, C.; Da Costa, M.S.; Van Uden, N. Osmoregulatory active sodium-glycerol co-transport in the halotolerant yeast Debaryomyces hansenii. Yeast 1990, 6, 187–191. [Google Scholar] [CrossRef]
- Dujon, B.; Sherman, D.J.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuvéglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef]
- Spier, M.R.; Vandenberghe, L.P.; Medeiros, A.B.; Soccol, R. Application of different types of bioreactor in bioprocesses. Bioreactors: Design, Properties and Applications. P.G. Antolli and Z. Liu. Nova Science Publishers 2011. pp. 55-90; Nova Science Publishers: New York, NY, USA, 2011; pp. 55–90. [Google Scholar]
- Guerriero, G.; Berni, R.; Muñoz-Sánchez, J.A.; Apone, F.; Abdel-Salam, E.M.; Qahtan, A.A.; Alatar, A.; Cantini, C.; Cai, G.; Hausman, J.-F.; et al. Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists. Genes 2018, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Bharathiraja, B.; Sudharsana, T.; Jayamuthunagai, J.; Pravenkumar, R.; Chozhavendhan, S.; Iyyappan, J. Biogas production—A review on composition, fuel properties, feed stock and principles of anaerobic digestion. Renew Sust Ener. Rev. 2018, 90, 570–582. [Google Scholar] [CrossRef]
- Raganati, F.; Olivieri, G.; Procentese, A.; Russo, M.E.; Salatino, P.; Marzocchella, A. Butanol production by bioconversion of cheese whey in a continuous packed bed reactor. Bioresour. Technol. 2013, 138, 259–265. [Google Scholar] [CrossRef]
- Raganati, F.; Procentese, A.; Olivieri, G.; Russo, M.E.; Gotz, P.; Salatino, P.; Marzocchella, A. Butanol production by Clostridium acetobutylicum in a series of packed bed biofilm reactors. Chem. Eng. Sci. 2016, 152, 678–688. [Google Scholar] [CrossRef]
- Dauglis, A.J. Integreted fermentation and recovery processes. Curr. Opin. Biotechnol. 1994, 5, 192–195. [Google Scholar] [CrossRef]
- Ezeji, T.C.; Karcher, P.M.; Qureshi, N.; Blaschek, H.P. Improving performance of a gas stripping-based recovery system to remove butanol from Clostridium beijerinckii fermentation. Bioprocess Biosyst. Eng. 2005, 27, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Rochón, E.; Ferrari, M.; Lareo, C. Integrated ABE fermentation-gas stripping process for enhanced butanol production from sugarcane-sweet sorghum juices. Biomass- Bioenergy 2017, 98, 153–160. [Google Scholar] [CrossRef]
- Gaykawad, S.S.; Zha, Y.; Punt, P.J.; Van Groenestijn, J.W.; Van Der Wielen, L.A.; Straathof, A.J.J. Pervaporation of ethanol from lignocellulosic fermentation broth. Bioresour. Technol. 2013, 129, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Hu, S.; Chen, C.; Wang, Y.; Zhang, C.; Miao, Q.; Qin, P.; Tan, T. Immobilized ethanol fermentation coupled to pervaporation with silicalite-1/polydimethylsiloxane/polyvinylidene fluoride composite membrane. Bioresour. Technol. 2016, 220, 124–131. [Google Scholar] [CrossRef]
- Aljundi, I.H.; Belovich, J.M.; Talu, O. Adsorption of lactic acid from fermentation broth and aqueous solutions on Zeolite molecular sieves. Chem. Eng. Sci. 2005, 60, 5004–5009. [Google Scholar] [CrossRef] [Green Version]
- Raganati, F.; Procentese, A.; Olivieri, G.; Russo, M.E.; Salatino, P.; Marzocchella, A. Bio-butanol separation by adsorption on various materials: Assessment of isotherms and effects of other ABE-fermentation compounds. Sep. Purif. Technol. 2018, 191, 328–339. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Company | Feedstock | Organism | Process | Product | Reference |
|---|---|---|---|---|---|
| Novo Nordisk | Glucose | S. cerevisiae | Fermentation | Human insulin | [5] |
| Biosynthia | Renewable sugars * | E. coli | Fermentation | Vitamins | [6,7] |
| Novozymes | * | Aspergillus oryzae Fungi Bacteria | Enzyme production | Bulk enzymes for food, beverages, agriculture, bioenergy and household care | [8] |
| LanzaTech | Dihydrogen, carbon monoxide and carbon dioxide | Clostridium autoethanogenum | Syngas fermentation | Ethanol | [9,10] |
| Avansya (DSM/Cargill) | Crude sugars * | S. cerevisiae | Recombinant yeast fermentation | Steviol glycosides | [11,12] |
| Lonza | * | Mammalian cells Pichia, Bacillus, E. coli | * | Pharmaceuticals, biopharmaceuticals, fine chemicals, enzymes | [13] |
| Feedstock | Lignin Content (%) | Availability | Amount (Mt y−1) | Transport Cost | Reference |
|---|---|---|---|---|---|
| Brewer’ Spent Grains | 15–27 | All year round | 8 | − 1 | [15,16] |
| Agricultural residues | 5–15 | Seasonal | 150 | + 2 | [17,19] |
| Potato peel | 30 | All year round | 0.5 | − | [17,19] |
| Fresh cut sector | 15–20 | All year round | 1.4 | − | [21,22] |
| Sugar beet pulp | 2 | Seasonal | 7 | − | [17,18] |
| Sugarcane bagasse | 25–28 | Seasonal | 10 | − | [20] |
| Coffee silverskin | 27 | All year round | 0.1 | − | [16,21] |
| OFMSW | 5 | All year round | 1000 | − | [23,24] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarrete, C.; Jacobsen, I.H.; Martínez, J.L.; Procentese, A. Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions. Processes 2020, 8, 768. https://doi.org/10.3390/pr8070768
Navarrete C, Jacobsen IH, Martínez JL, Procentese A. Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions. Processes. 2020; 8(7):768. https://doi.org/10.3390/pr8070768
Chicago/Turabian StyleNavarrete, Clara, Irene Hjorth Jacobsen, José Luis Martínez, and Alessandra Procentese. 2020. "Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions" Processes 8, no. 7: 768. https://doi.org/10.3390/pr8070768
APA StyleNavarrete, C., Jacobsen, I. H., Martínez, J. L., & Procentese, A. (2020). Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions. Processes, 8(7), 768. https://doi.org/10.3390/pr8070768