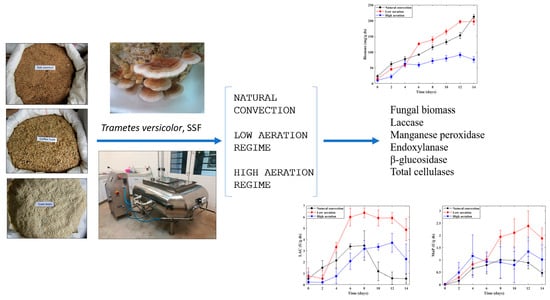
3.1. Solid State Cultivation of T. versicolor in a Fixed-Bed Bioreactor
In this work, SSF process was performed in the novel bioreactor described in
Section 2.3 (
Figure 1) using
T. versicolor to evaluate the lignocellulolytic enzymes production under two levels of forced aeration and no air injection. In
Figure 2, it is observed how the colonization of the substrates was developed by
T. versicolor under the three aeration levels evaluated at 14 incubation days. For LAR, the formation of a denser and more homogeneous mycelium on the substrate beds was noticed; these beds were completely colonized on the eighth fermentation day, equivalent to a substrate colonization rate (SCR) of 1.25 kg/day colonized substrate. The substrate was colonized on the ninth day when the HAR was applied (SCR of 1.11 kg/day), and on the 12th day when no air was injected into the bioreactor (SCR of 0.83 kg/day). In a previous work [
7], an SCR of 0.75 kg/day was determined during
T. versicolor vegetative phase in similar culture media packed in 3-kg bag units; this indicates the efficiency of the process in the pilot bioreactor used. It should be noted that the purpose of
T. versicolor cultivation in bags is the production of its fruiting bodies, while the objective of the culture in the bioreactor was to obtain lignocellulolytic enzymes.
Throughout the cultivation, an average 60% moisture in the substrates and 90–95% relative humidity within the bioreactor were maintained. This indicates that the air injection and circulation system inside the bioreactor, along with the relative humidity control system, kept the substrate moisture stable, which is crucial for the fungal biomass development. The results obtained showed that LAR increased the formation of exudates, which are related to water synthesis from the fungal aerobic respiratory metabolism. The water formed leaches soluble and potentially valuable substances through the substrate bed by gravity, which are collected in the lower part of the bioreactor.
The heat transfer is one of the aspects generating the greatest difficulties during the operation of solid-state large-scale fermenters, so temperature control inside the beds is one of the main concerns arising during the work of such bioreactors. Overheating the beds can dry the substrate and cause damage to the hyphae of the fungus, which significantly restricts the synthesis of valuable products, such as enzymes. The design of the bioreactor used, allowed for adequate temperature control compared to conventional tray bioreactors. In the latter, the bed height cannot be very high (between 5 and 8 cm) to avoid the accumulation of metabolic heat inside the substrate, which is removed only by contact with the air circulating between the trays [
30]. In contrast, the substrate boxes provided with cooling tubes in the pilot bioreactor employed do allow more efficient heat removal. In this sense, although the air injection into this bioreactor was intended to activate
T. versicolor biomass production and its ligninolytic enzymes, it helps to remove the heat from the beds, as well. HAR did show a slight change in temperature reaching average values between 27 °C and 30 °C, compared to the NC regime that reached an average bed temperature of 33 °C at the third day after the bioreactor inoculation. Therefore, the beds under NC were cooled by recirculating cold water; thus, it was possible to restore the temperature to the previously defined value of 30 °C in a time of 125 min.
During SSF, microbial cells take in oxygen directly from the air. However, several operational factors affect the oxygen transfer rate, such as substrate porosity, material layer thickness, substrate moisture, reactor design, and mechanical agitation, if present [
31]. In addition, the technology of solid-state fermentation in tray bioreactors is not fully developed. In this regard, the usage of forced ventilation is not common, and the transport of oxygen and carbon dioxide mostly depends on the diffusion, which generates important constraints to the transfer of mass and heat during the cultivation process. In general, for bioreactors operating under natural convection regimen, oxygen uptake by filamentous fungi is much higher than oxygen supply in the bioreactor. This fact diminishes the availability of oxygen that may be dissolved in the biofilm formed on the surface of the solid substrate particles, generating an oxygen concentration gradient during the aerobic fermentation. In this work, the application of two regimes of forced aeration seeks to avoid this issue. In fact, an improved behavior of
T. versicolor was obtained by using the low aeration regime for producing cell biomass during the vegetative growth phase, as well as a higher lignin degradation in the culture medium (see
Section 3.4). It is likely that the LAR supplied an appropriate level of oxygen required by the mushroom for its vegetative growth phase, as well as the proper balance between oxygen and carbon dioxide needed for this phase. In this regard, very high oxygen levels can retard the growth of several types of microorganisms, among which several species of filamentous fungi are found, as has been empirically demonstrated in different investigations [
15,
18,
32]. Nevertheless, it is still necessary to understand what is modified in the cellular metabolism of fungi that experience this effect.
For a SSF process with filamentous fungi, Oostra et al. [
33] concluded that oxygen variation in the layer of aerial hyphae was relatively slight, but oxygen concentration severely changed in the layer of wet cells when forced air was injected. In addition, oxygen concentrations were reduced with the increase of the solid bed depth and the progress of the fermentation. Likewise, these authors determined that the oxygen content is dynamical and is influenced by the carbon dioxide generated by the microbial metabolism. Therefore, it is evident that oxygen supply inside the solid particles and hyphae are two key factors affecting the solid-state fermentation process, so more research efforts on this aspect should be conducted in the future, especially through the design and operation of novel types of fixed-bed bioreactors for the production of macromycetes and their metabolites with commercial interest as the one employed in this work.
3.2. Production of T. versicolor Biomass
T. versicolor biomass production in the solid-state pilot-scale bioreactor is illustrated in
Figure 3. During most fermentation time, the highest biomass production was obtained for LAR. A progressive growth of the fungal biomass (characteristic of a vigorous growth phase) was observed with a maximum concentration of 202.4 mg/g ds on the 14th cultivation day; this aeration regime showed statistically significant differences throughout cultivation compared to the other two treatments evaluated (
p-value = 0.0456). This can be explained by the stimulation provided by the low aeration level during the vegetative growth of the fungus, which allows an appropriate air exchange for its development. The other aeration regimes also showed progressive growth of the fungal biomass, reaching maximum concentrations of 91.04 mg/g ds for HAR and 204.7 mg/g ds for NC. However, HAR showed a very slow growth during the first two days (close to a latency phase), a growth phase until day 10, and a slow growth phase considered as a stationary phase between days 10 and 14; this behavior could be due to an excess of air exchange that can disperse the CO
2 levels necessary for the vegetative fungal growth. In contrast, biomass production under NC did not show stationary nor lag phases, which can be attributed to the stimulus occurring in the vegetative growth phase of several white rot fungi in the presence of higher levels of carbon dioxide, as indicated by Chang and Miles [
32] and in another previous work [
21].
T. versicolor growth curve for LAR reached the stationary phase towards the 12th day and the biomass concentrations were significantly higher than those of NC treatment from the sixth day until the 12th day; the final biomass concentration for LAR and NC were almost the same. Thus, low aeration level makes possible to shorten the fermentation time when this fungus is grown in the pilot bioreactor used in this work compared to the case when no air is injected. The colonization of solid substrates by white rot fungi is slower when it is not possible to control all operating conditions, which makes the processes vulnerable to contamination by competitors with the consequent increase in the costs; that is not the case of the bioreactor employed in which improved performance is attributed to the control of its operating conditions and the formulation of the culture medium used.
The biomass concentrations are similar to those obtained in a previous work [
5] for
T. versicolor grown on 12 solid media containing oak sawdust, coconut fiber, coffee husk, soy oil, corn bran, calcium carbonate, and copper(II) sulfate with different concentrations of these components; it is highlighted that the medium used in this work only contained oak sawdust, coffee husk, corn bran, and small amounts of sugar, copper(II) sulphate, manganese(II) sulfate, and calcium carbonate. In that work, the fungus was cultivated in 200-g polypropylene bags for 49 days. The biomass concentrations reached in most media on the 14th day were below the biomass content achieved in the present work. Only in one medium, the biomass content was significantly higher (about 260 mg/ds) on the 14th day in plastic bags; however, this was not the medium that reached the maximum fungal concentration after 49-day cultivation. As the main goal of the cultivation in the bioreactor employed is the production of ligninolytic enzymes, the fermentation process was finished on the 14th day. More research is needed to assess the level of fungal biomass that could be achieved in the fixed-bed pilot-scale bioreactor. In this regard, it is evident that LAR favors the substrate colonization by the fungus at a rate higher than that of the bags for the first 14 days of cultivation, as noted in
Section 3.1.
The operation of solid-state fermenters for producing metabolites with macromycetes on an industrial scale presents difficulties when adequate bed compaction is not guaranteed. The compaction is due to the thickening of the hyphae during the macromycetes vegetative phase as a way to prepare the substrates for the subsequent fructifying (reproductive) phase. This causes a decrease in the porosity of the solid materials, as well as the change in the structure of the meso and micropores of the substrate particles, due to the degradation and consumption of their constituting biopolymers. These structural changes are necessary to obtain several metabolites during the vegetative phase but also affect the mass and heat transfer in the bed. Therefore, improved designs of solid-state bioreactors for macromycetes have to enhance these transfer phenomena, ensuring at the same time the substrate compaction. Fixed-bed bioreactors offer better operating conditions with lower costs because they guarantee the compaction of the substrates. Precisely, the pilot bioreactor used in this work allowed an adequate substrate compaction for the development of T. versicolor, which is evidenced in the fungal biomass formation data for different aeration levels. This bioreactor design surpasses traditional fixed-bed fermenters due to the automatic control of humidity and temperature, as well as the air flow regulation. These characteristics are crucial to overcome the disadvantages related to heat and mass transfer in conventional tray reactors.
The RS variation during SSF in the fixed-bed bioreactor showed the same trend that occurred in previous works under NC conditions, not only with this macromycete but also with white rot fungi, such as
Pleurotus ostreatus and
Lentinus edodes [
7], and with
Grifola frondosa [
34]. The RS profiles over time (see
Figure 3) show a variation in a range between 4 and 25 mg/g ds for the three regimes. The maximum RS values reached for the three treatments were 23.92 mg/g ds on the 12th day for LAR, 13.11 mg/g ds on the 10th day for HAR, and 9.27 mg/g ds on the 6th day for NC. These values coincide with the start of the stationary phase for the two forced aeration regimes, and with the middle of the growth phase for the NC treatment. It is presumed that the RS are generated at a constant rate throughout the fermentation as a result of the degradation of carbohydrates present in the substrate, so that their variation in time is due to the consumption of simple sugars by the fungi as an energy source necessary for their growth. Additionally, selective lignin-degrading fungi, such as
T. versicolor, use carbohydrates other than cellulose at the beginning of fermentation to obtain their energy source, so the fungus had to first degrade the hemicellulose and starch present in the substrate [
35].
3.3. Production of Lignocellulolytic Enzymes
The activities of laccase, manganese peroxidase, endoxylanase, β-glucosidase, and total cellulases from
T. versicolor during the 14 days of SSF in the pilot bioreactor did not show a defined time trend for any of the three regimes studied. The maximum enzymatic activities reached during the SSF process are presented in
Table 1. In this work, we sought to especially evaluate the air flowrate effect in the fixed-bed bioreactor on the production of LAC and MnP from
T. versicolor grown on agro-industrial waste. The maximum LAC activity for LAR was 6.37 U/g ds, approximately twice the maximum activity obtained for the other two regimes for 14 fermentation days. It is worth noting the considerable increase in the activity of this enzyme during 8 fermentation days compared to the activity attained under NC. This behavior was expected due to the dependence on oxygen that laccase has for its expression.
The progress of the LAC activity with time for the three regimes studied is shown in
Figure 4. The LAC activity for LAR showed a rapid growth during the first six days of cultivation, and then stabilized in values between 4.9 and 6.4 U/g ds in the next six days with a slight decrease towards the 14th day. It is evident that both the absence of air injected into the bioreactor and the HAR lead to lower LAC values, which coincided practically during the first 8 days; however, from the tenth day, the LAC titers were significantly reduced to values close to zero for NC. Therefore, the positive effect of the air injection at low level on the LAC activity using the novel bioreactor reported in this work is very noticeable. This can be explained considering that LAC uses oxygen as a substrate, which makes it directly dependent on this compound. NC regime implies lower oxygen levels during fermentation and, consequently, lower LAC values. However, HAR led to lower LAC titers (
Figure 4), which could be due to the correspondence between the LAC synthesis and the primary metabolism of the fungus for biomass formation. As noted above, this process of cell formation requires moderate CO
2 levels, so a high aeration frequency removes these levels, decreasing the metabolic activity of the fungus in its vegetative phase and, therefore, the LAC values.
MnP activity showed no statistically significant differences for any of the three regimes studied. The maximum enzymatic activity obtained from MnP was reported on the 12th day of fermentation for LAR (see
Table 1). This value was approximately double the maximum activities obtained for HAR and NC. The MnP activity is markedly lower than those of the other enzymes measured (see
Table 1), probably due to causes associated with the extraction methods used for its determination. This enzyme is very sensitive to deactivation when subjected to physical or chemical treatments [
7]. This fact could have provoked greater standard deviations in its enzymatic activity, which implied that the analysis of variance showed that there were no statistically significant differences between the treatments. This is also highlighted by Postemsky et al. [
36], who evaluated the influence of enzyme extraction methods on the result of the enzymatic activity assays, finding large differences in the results obtained according to the selection of the extraction method.
The time profile of the MnP enzymatic titers for LAR showed an increasing behavior during most of the fermentation (until day 12) compared to NC (
Figure 4). For the latter case, a maximum value of the MnP activity was observed towards the middle of the cultivation process with a gradual decrease to values close to zero towards day 14. MnP activity for HAR presented notable oscillations throughout the fermentation, but in general the values obtained were between the values corresponding to the other two treatments. MnP is a hydrogen peroxide dependent enzyme, which it uses as a substrate. The fungus requires the oxygen presence to form hydrogen peroxide through several pathways involving the action of glucose-oxidase and glyoxal oxidase [
15], making MnP indirectly dependent on oxygen. In this work, LAR during
T. versicolor cultivation led to a greater MnP synthesis according to most data obtained from the samples taken (see
Figure 4b). Meanwhile, HAR led to enzymatic MnP titers comparable to those of NC, which can be explained again by the correspondence between biomass formation and moderate CO
2 levels in the bioreactor. Therefore, the forced aeration exerts an important influence on the synthesis of ligninolytic enzymes when a stationary fixed-bed bioreactor as that employed in this work is used.
The enzymatic activity curves showed noticeable decreases at different times, especially for NC and HAR. This fact could be attributed to changes in the substrate composition during the fermentation, since new substances that inhibit enzymatic activities can be produced; to the inhibiting by-products of the reactions occurring during fermentation; and to the production of isoenzymatic variants of the same enzymes at different times and different operating conditions causing changes in enzymatic titers. This reduction in ligninolytic activities with time have also been detected by several researchers, who report some of these hypotheses as possible causes of decreasing said enzymatic activities [
34,
37].
There is a considerable variation of the LAC and MnP activities disclosed in the literature. Zhu and Zhang [
38] reported LAC and MnP activities of
T. versicolor incubated on mixtures of lignocellulosic materials of 45.1 U/g ds and 1.35 U/g ds, respectively, when control parameters for heat removal in laboratory-scale beds were optimized. Postemsky et al. [
36] reported LAC titers of 10.93 U/g ds on a rice straw and husks and 16.44 U/g ss on a substrate based on sunflower seed hulls in the full colonization phase for
Ganoderma lucidum grown in a pilot-scale bioreactor. These values are higher than those obtained in this work, although it should be considered that the substrate used was not supplemented with phenolic inductors or oils, such as those used in works, like that of Postemsky et al. [
36].
For the hydrolytic enzymatic activities measured in this work (ENX, βG, and TC), there were mostly no statistically significant differences between the three treatments studied, probably because the action of these enzymes does not depend on oxygen. ENX was the only one that exhibited the highest enzymatic activity (9.05 U/g ds on the tenth day of culture) under NC against the two treatments with forced aeration, obtaining statistically significant differences (
p-value = 0.0005). The other enzymatic activities showed their maximum values at the LAR (see
Table 1).
Figure 5 shows the time profile of the ENX titers for the three regimes. The ENX profiles obtained do not show a clear trend with time. In fact, the data of this activity oscillate in a range of about 20% of its average value over time for NC and about 50% for the two forced aeration levels. These variations are comparable with the standard deviations of the mean value of the experimental ENX measurements, which is related to the random errors of the analytical procedure employed (shown in
Figure 5 through error bars). Nevertheless, higher values can be evidenced under NC compared to the activities under forced aeration. This behavior denotes that this hydrolase does not exhibit oxygen dependence for its expression. Different authors [
15,
39] have pointed out that hemicellulose is the first polysaccharide attacked by white rot fungi in natural environments from which they obtain the most available energy source at the beginning of the substrate degradation in the form of simple sugars, such as xylose, arabinose, and mannose. These monosaccharides are integrated into the primary fungal metabolism for cell biomass synthesis without oxygen involvement. This could explain why forced aeration levels do not increase the synthesis of ENX.
In the case of LAR, the maximum activities of the cellulase group evaluated as TC and of the βG reached values of 11.58 U/g ds and 350.28 U/g ds, respectively (see
Table 1). Likewise, the standard deviations of the data obtained were quite high and many of them overlapped, which could be due to the experimental errors of corresponding laboratory procedures. It should be noted that the cultivation time studied in the fixed-bed bioreactor was 14 days, a time chosen to especially evaluate the production of ligninolytic enzymes. However, the synthesis of cellulases is induced by the presence of cellulose available in the medium.
T. versicolor first attacks lignin to subsequently release cellulose from solid substrates, so that the majority action of these enzymes occurs at a stage after the initial lignin degradation. Considering that oak sawdust is the main component of the substrate employed in this work, it is worth to point out that Bari et al. [
40] have demonstrated that lignin degradation precedes the degradation of polysaccharides during the oak wood decay by
T. versicolor. This explains why TC are not very high during the cultivation time studied in this work; with longer fermentation times, the synthesis of cellulases will be greater, as will the percentage of cellulose degradation in the medium. On the other hand, βG titers obtained are high compared to those reported in the available literature for 14 days of cultivation [
41,
42]. These high values could be related to the potential presence of structural analogues of cellobiose, the natural substrate of this enzyme, in the components of the solid substrate.
Fixed-bed bioreactors have exhibited higher performance for producing lignocellulolytic enzymes with macromycetes. The production of ligninases by
T. versicolor in the bioreactor used in this investigation varying the aeration levels, showed consistency with the studies conducted by other researchers who also proposed various bioreactor models with the same purpose. Rosales et al. [
43] indicated that the use of a tray bioreactor for laccase production using
Trametes hirsuta guarantees the best conditions for the process. LAC enzymatic activities reached in that tray bioreactor were three times higher than those obtained in a tubular bioreactor, the other equipment design assessed by these authors. This can be explained considering that the agitation generated in the tubular bioreactor caused mechanical stress to the fungus, reducing the enzyme production.
3.4. Degradation of the Main Biopolymers of the Lignocellulosic Biomass
The degradation percentages of lignin, cellulose, and hemicellulose achieved at the end of the SSF evaluated in this work are presented in
Table 2. Lignin degradation and consumption of carbohydrate fibers by the extracellular enzymatic consortium of
T. versicolor has been reported elsewhere; in particular, lignin degradation is mostly attributed to the constant LAC and MnP secretion [
44]. The highest degradation of lignin was achieved for low aeration regime. In this regard, it is worth to note that this regime is the one with the highest titers of LAC and MnP (see
Figure 4a) during the cultivation process. This suggests that high percentages of lignin degradation are directly related to the synthesis of LAC and MnP. In this sense, the leading action of laccase could explain most of the primary degradation of lignin, especially if taking into account that this enzyme was expressed the most under the low aeration regime due to its enhanced synthesis when moderate oxygen levels are available. HAR is not appropriate for lignin degradation considering that an elevated air flowrate may lead to the dispersion of the necessary levels of CO
2 during the cultivation. This dispersion, in turn, may reduce the primary metabolism of the fungus, i.e., the biomass growth and the synthesis of the related ligninases needed for the primary degradation of the lignocellulosic substrates are reduced under high aeration conditions, as shown in
Figure 3a and
Figure 4.
The low degradation percentages for cellulose and hemicellulose depicted in
Table 2 are explained by the short operating time selected for this work as mentioned above. In this sense, it is necessary to conduct more research to ascertain whether the high aeration regime can lead to higher cellulose degradation or not when the fermentation time is prolonged. On the other hand, the hemicellulose degradation did not show significant differences among the aeration regimes studied in this work.
When comparing the metabolite production by SSF and SmF, the lower water content and the existence of a gas phase between the particles in the former are highlighted. Regarding this, the thermal conductivity of the air is very poor during SSF compared to that of water in the case of SmF. For this reason, it is important to study the influence of aeration regime on enzyme production in SSF processes. Unlike the small number of published works on the effect of oxygen on lignocellulolytic enzymes produced by SSF on a pilot scale, studies on SmF with oxygen injection for cultivating white rot fungi and industrial dye discoloration are more frequent in the scientific literature [
45,
46]. Thus, the importance of the data generated in this research is highlighted. Based on the results obtained, it was evident that the SSF of agro-industrial waste using
T. versicolor in the fixed-bed bioreactor with natural convection and forced aeration, reached an efficient colonization of the substrates, a high production of fungal biomass, and high lignocellulolytic enzymatic titers. This enhanced performance is related to the satisfactory control of the temperature, substrate moisture, and relative humidity within the equipment, as well as the regulation of the aeration regime. Finally, the fungus
T. versicolor is an attractive organism able to degrade a wide range of lignocellulosic materials in order to valorize different agricultural and agro-industrial residues through the production not only of lignocellulolytic enzymes but also of other value-added products, like bioactive polysaccharides [
5], antimicrobial agents [
10], biofuels [
1], and packaging materials [
47], among others.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}