
Optimum Design of N Continuous Stirred-Tank Bioreactors in Series for Fermentation Processes Based on Simultaneous Substrate and Product Inhibition

Abstract
:1. Introduction
- The cells generated in the first few reactors will continue to metabolize and produce ethanol in the latter reactors.
- In multistage CSTRs, cells are exposed to suitable growth conditions in the first few reactors (low product concentration and high substrate concentration) compared to a single CSTR where high inhibitory product concentration and low substrate concentration are present.
- In multistage CSTRs, inhibitory product concentration increases gradually from one reactor to the other, while in single CSTR, a high concentration of inhibitory product is present from the beginning (i.e., reactors in series reduces the effect of product inhibition). Recently, Wang et al. [21] experimentally evaluated two CSTRs in series with cell recycling after each reactor for the production of ethanol. This system was found to be effective in ethanol production because of the high cell concentration (cell recycling) and due to a certain degree of relief in substrate and ethanol inhibition.
2. Methodology
2.1. Kinetic Equation with Substrate and Product Inhibition
2.2. Continuous Stirred-Tank Reactors in Series with Total Minimum Volume
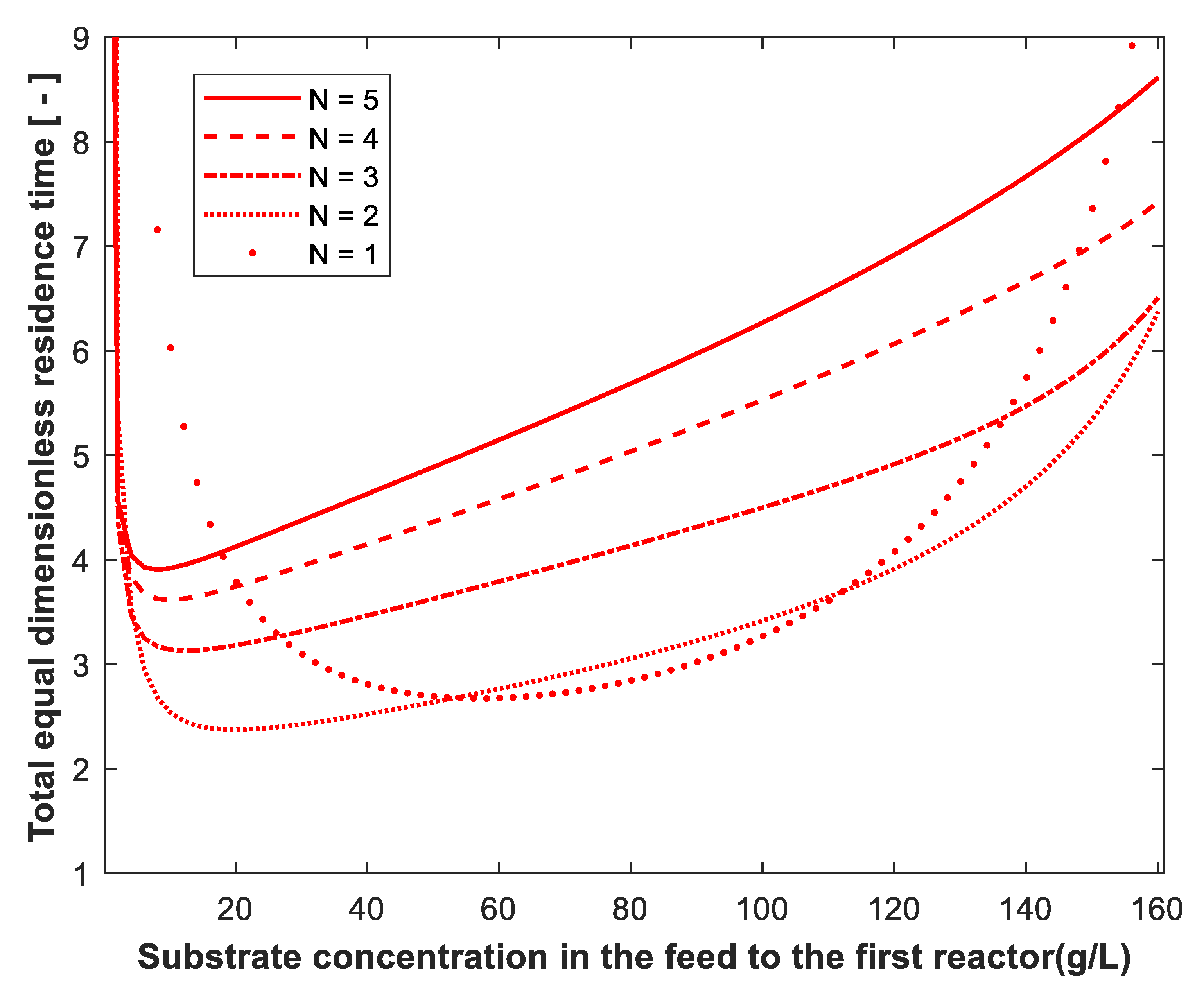
2.3. Continuous Stirred-Tank Reactors of Equal Size in Series
2.4. Percentage Reduction in Total Residence Time Using the Optimum Design Criteria
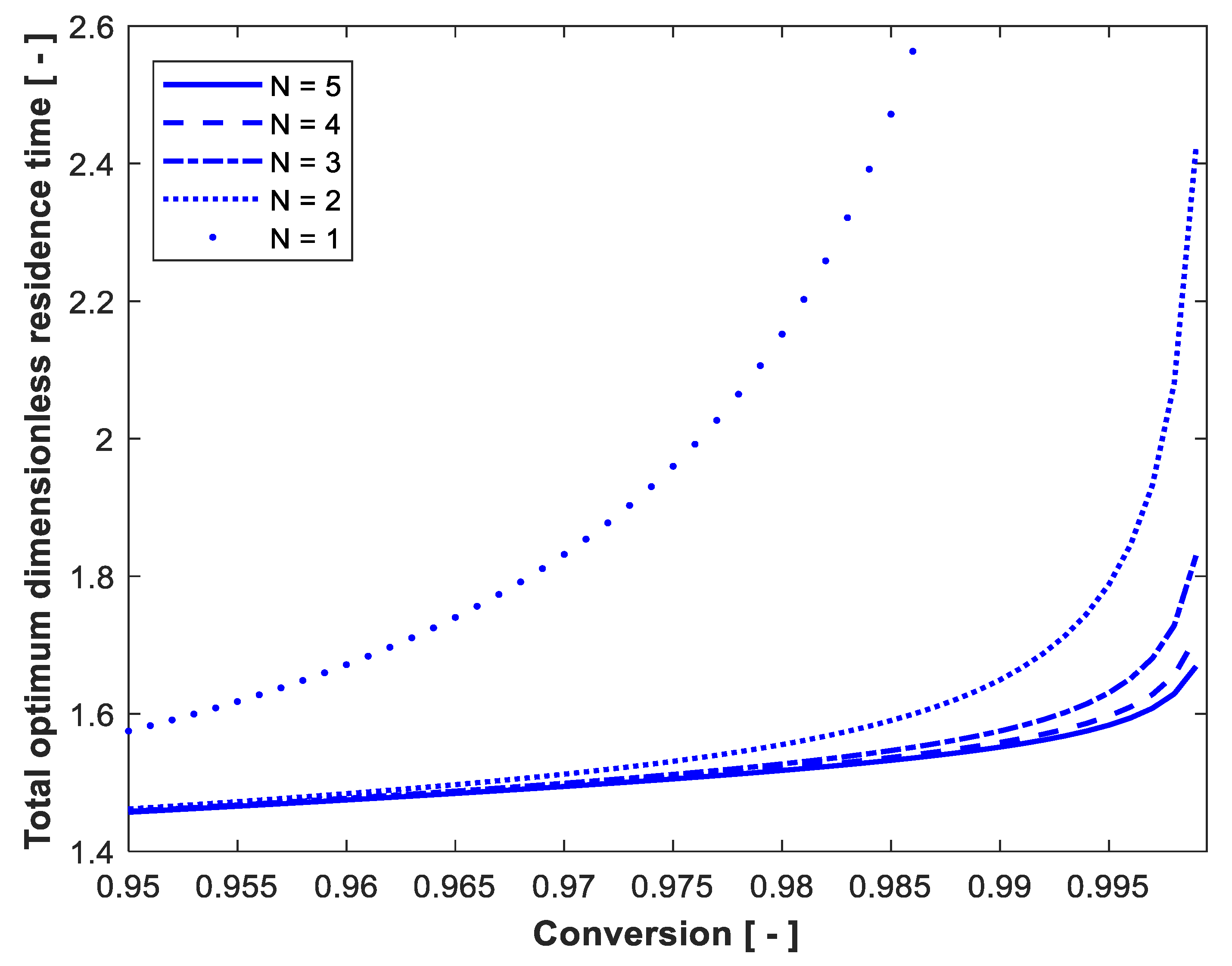
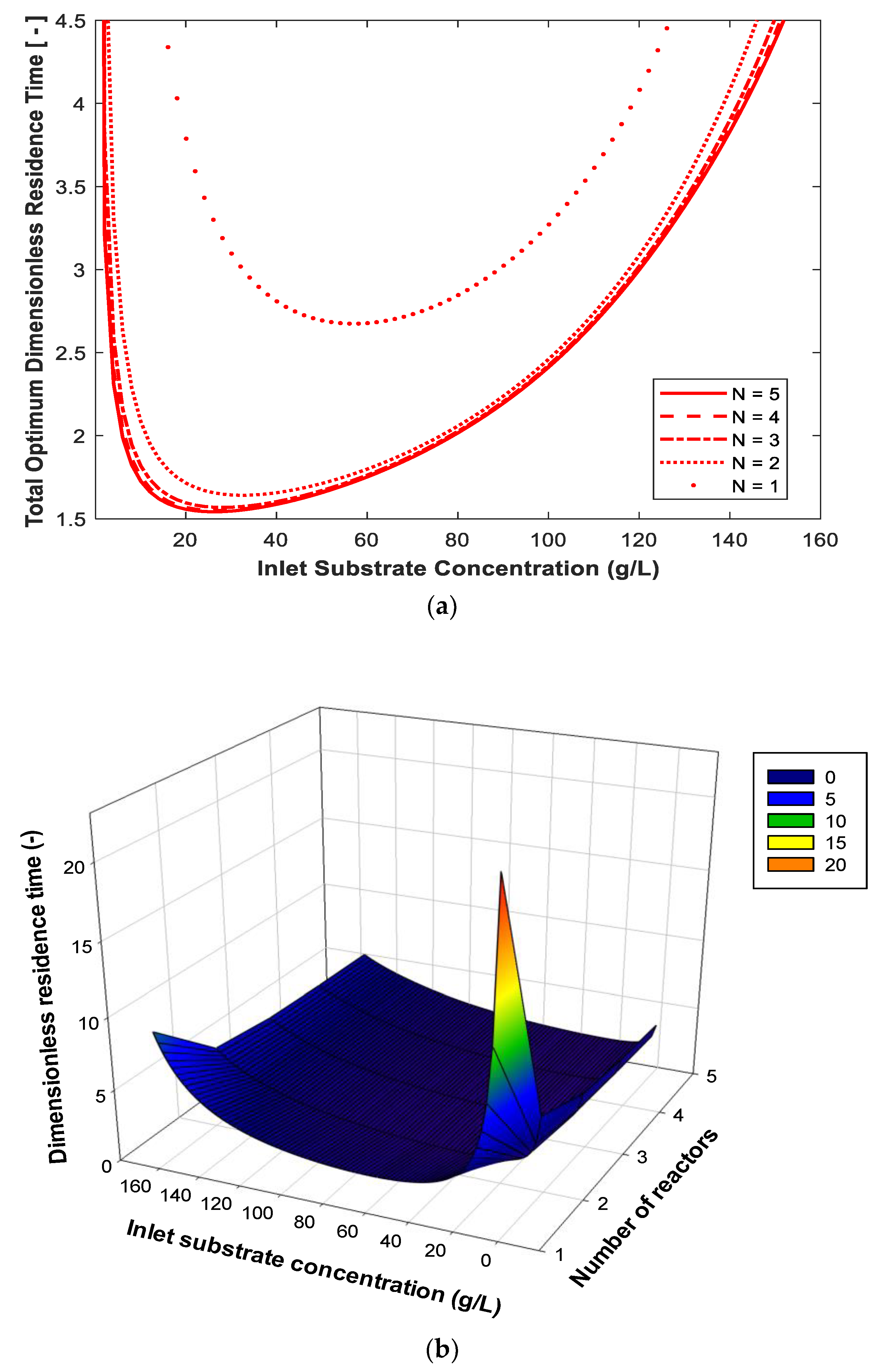
3. Results and Discussion
4. Conclusions
Funding
Conflicts of Interest
Nomenclature
| constant defined as | |
| constant defined as | |
| constant defined as | |
| liquid flow rate (L/h) | |
| Monod constant (g/L) | |
| dimensionless Monod constant | |
| inhibition constant (g/L) | |
| dimensionless inhibition constant | |
| number of CSTRs in series | |
| substrate concentration (g/L) | |
| product concentration (g/L) | |
| maximum or critical product concentration (g/L) | |
| reactor volume, L | |
| cell concentration, g/L | |
| cell yield coefficient, g biomass/g substrate | |
| product yield coefficient, g product/g substrate | |
| Greek Symbols | |
| dimensionless substrate concentration | |
| residence time, h | |
| dimensionless residence time | |
| specific growth rate, h−1 | |
| maximum specific growth rate, h−1 | |
| degree of substrate conversion | |
| Subscripts | |
| refers to the ith reactor | |
| refers to the Nth reactor | |
| initial | |
| total | |
| equal | |
| optimum | |
References
- Nguyen, V.D.; Auresenia, J.; Kosuge, H.; Tan, R.R.; Brondial, Y. Vacuum fermentation integrated with separation process for ethanol production. Biochem. Eng. J. 2011, 55, 208–214. [Google Scholar] [CrossRef]
- Ghose, T.K.; Tyagi, R.D. Rapid ethanol fermentation of cellulose hydrolysate. II. Product and substrate inhibition and optimization of fermenter design. Biotechnol. Bioeng. 1979, 21, 1401–1420. [Google Scholar] [CrossRef]
- Moulin, G.; Boze, H.; Galzy, P. Inhibition of alcoholic fermentation by substrate and ethanol. Biotechnol. Bioeng. 1980, 22, 2375–2381. [Google Scholar] [CrossRef]
- Van den Heuvel, J.C.; Beeftink, H.H. Kinetic effects of simultaneous inhibition by substrate and product. Biotechnol. Bioeng. 1988, 31, 718–724. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, D.; Lin, Y.; Wang, X.; Kong, H.; Tanaka, S. Substrate and product inhibition on yeast performance in ethanol fermentation. Energy Fuels 2015, 29, 1019–1027. [Google Scholar] [CrossRef]
- Zentou, H.; Abidin, Z.Z.; Yunus, R.; Biak, D.R.A.; Issa, M.A.; Pudza, M.Y. A new model of alcoholic fermentation under a byproduct inhibitory effect. ACS Omega 2021, 6, 4137–4146. [Google Scholar] [CrossRef] [PubMed]
- Sunarno, J.N.; Prasertsan, P.; Suwan, W.D.; Kongjan, P.; Cheirslip, B. Mathematical modeling of ethanol production from glycerol by Enterobacter aerogenes concerning the influence of impurities, substrate and product concentration. Biochem. Eng. J. 2020, 155, 107471. [Google Scholar] [CrossRef]
- Sirisansaneeyakul, S.; Hipolito, C.N.; Kobayashi, G.; Lertsiri, S.; Luangpituksa, P.; Varavinit, S.; Ishizaki, A. Kinetic modeling of lactic acid fermentation from sago starch using Lactococcus IO-1. Annu. Rep. IC Biotechnol. 1998, 21, 504–524. [Google Scholar]
- Goncalves, L.M.D.; Xavier, A.M.R.B.; Almeida, J.S.; Carrondo, M.J.T. Concomitant substrate and product inhibition kinetics in lactic acid production. Enzym. Microb. Technol. 1991, 13, 314–319. [Google Scholar] [CrossRef]
- Balakrishnan, R.; Tadi, S.R.R.; Rajaram, S.K.; Mohan, N.; Sivaprakasam, S. Batch and fed-batch fermentation of optically pure D(-) lactic acid from kodo millet (Paspalum scrobiculatum) bran residue hydrolysate: Growth and inhibition kinetic modeling. Prep. Biochem. Biotechnol. 2020, 50, 365–378. [Google Scholar] [CrossRef]
- Moeller, L.; Strehlitz, B.; Aurich, A.; Zehnsdorf, A.; Bley, T. Optimization of citric acid production from glucose by Yarrowia lipolytica. Eng. Life Sci. 2007, 7, 504–511. [Google Scholar] [CrossRef]
- Lin, S.K.C.; Du, C.; Koutinas, A.; Wang, R.; Webb, C. Substrate and product inhibition kinetics in succinic acid production by Actinobacillus succinogenes. Biochem. Eng. J. 2008, 41, 128–135. [Google Scholar] [CrossRef]
- Ferone, M.; Raganati, F.; Olivieri, G.; Salatino, P.; Marzocchella, A. Continuous succinic acid fermentation by Actinobacillus succinogenes. Assessment of growth and succinic acid production kinetics. Appl. Biochem. Biotechnol. 2019, 187, 782–799. [Google Scholar] [CrossRef] [PubMed]
- Vlysidis, A.; Binns, M.; Webb, C. Utilization of glycerol to platform chemicals within the biorefinery concept: A case for succinate production. Chem. Eng. Trans. 2009, 18, 537–542. [Google Scholar]
- Van Niel, E.W.J.; Claassen, P.A.M.; Stams, A.J.M. Substrate and product inhibition of hydrogen by the extreme thermophile, Caldicellulosiruptor saccharolyticus. Biotechnol. Bioeng. 2003, 81, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yin, Y.; Wang, J. Recent advance in inhibition of dark fermentative hydrogen production. Int. J. Hydrogen Energy 2021, 46, 5053–5073. [Google Scholar] [CrossRef]
- Lopez, S.J.; Sola, C.; Lema, J.M. Substrate and product inhibition significance in the kinetics of sucrose hydrolysis by invertase. Biotechnol. Bioeng. 1982, 24, 2721–2724. [Google Scholar]
- Mukesh, D.; Salunke, R.; Banerji, A.A.; Bevinakatti, H.S. Modeling of lipase catalyzed transesterification reactions. Biocatalysis 1992, 6, 151–159. [Google Scholar] [CrossRef]
- Horvat, P.; Spoljaric, I.V.; Lopar, M.; Atlic, A.; Koller, M.; Braunegg, G. Mathematical modelling and process optimization of a continuous 5-stage bioreactor cascade for production of poly[-(R)-3-hydroxybutyrate] by Cupriavidusnecator. Bioprocess Biosyst. Eng. 2013, 36, 1235–1250. [Google Scholar] [CrossRef]
- Hill, G.A.; Robinson, C.W. Minimum tank volumes for CFST bioreactors in series. Can. J. Chem. Eng. 1989, 67, 818–824. [Google Scholar] [CrossRef]
- Wang, F.S.; Li, C.C.; Lin, Y.S.; Lee, W.C. Enhanced ethanol production by continuous fermentation in a two-tank system with cell recycling. Process Biochem. 2013, 48, 1425–1428. [Google Scholar] [CrossRef]
- Wall, J.B.; Hill, G.A. Optimum CFST bioreactor design. Experimental study using batch growth parameters for S. cerevisiae producing ethanol. Can. J. Chem. Eng. 1992, 70, 148–152. [Google Scholar] [CrossRef]
- Kalogeris, E.; Antzoulatos, O.; Mamma, D.; Hatzinikolaou, D.G.; Christakopoulos, P.; Kekos, D. Application of different processes for the biodegradation of 1,3-dichloro-2-propanol by the bacterium Pseudomonas putida DSM 437. Chem. Biochem. Eng. Q. 2007, 21, 297–305. [Google Scholar]
- Nishiwaki, A.; Dunn, I.J. Analysis of the performance of a two-stage fermenter with recycle for continuous ethanol production using different kinetic models. Biochem. Eng. J. 1999, 4, 37–44. [Google Scholar] [CrossRef]
- Nishiwaki, A.; Dunn, I.J. Analysis of a two-stage fermenter with recycle for continuous ethanol production. Chem. Eng. Comm. 1998, 168, 207–227. [Google Scholar] [CrossRef]
- Harmand, J.; Rapaport, A.; Trofino, A. Optimal design of interconnected bioreactors: New results. AIChE J. 2003, 49, 1433–1450. [Google Scholar] [CrossRef]
- Bayrock, D.P.; Ingledew, W.M. Ethanol production in multistage continuous, single stage continuous, Lactobacillus-contaminated continuous, and batch fermentations. World J. Microbiol. Biotechnol. 2005, 21, 83–88. [Google Scholar] [CrossRef]
- Abu Reesh, I.M. Optimal design for CSTRs in series using reversible Michaelis-Menten reactions. Bioprocess Eng. 1996, 15, 257–264. [Google Scholar]
- Abu Reesh, I.M. Optimal design of continuously stirred tank reactors in series using Michaelis-Menten kinetics with competitive product inhibition: Theoritical analysis. Desalination 2005, 180, 119–132. [Google Scholar] [CrossRef]
- MATLAB, Version (R2020b); The MathWorks Inc.: Natick, MA, USA, 2019.
- Dourado, A.; Goma, G.; Albuquerque, U.; Sevely, Y. Modeling and static optimization of the ethanol production in a cascade reactor. I. Modeling. Biotechnol. Bioeng. 1987, 29, 187–194. [Google Scholar] [CrossRef]
- Dourado, A.; Calvet, J.L.; Sevely, Y.; Goma, G. Modeling and static optimization of the ethanol production in a cascade reactor. II. Static optimization. Biotechnol. Bioeng. 1987, 29, 195–203. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetics and Stoichiometric Coefficients | Symbol | Value and Units |
|---|---|---|
| Maximum specific growth rate | 0.4 | |
| Monod constant | 0.48 | |
| Inhibition constant | 205.2 | |
| Maximum product concentration | 87 | |
| Cell (biomass) yield coefficient | 0.1 | |
| Product yield coefficient | 0.48 |
| Fermentation Process | N Reactors/Equal or Unequal Size | Reactor Performance/Volume | Ref. |
|---|---|---|---|
| Ethanol fermentation by the yeast S. cerevisiae, 30 °C, pH of 4, bagasse hydrolysate as substrate. | 2 equal-size reactors in series | Volume reduction using 2 CSTRs compared to one reactor is 58%. | [2] |
| Ethanol fermentation by S. cerevisiae using glucose as substrate. | 1 and 5 equal-size reactors | Ethanol productivity in 5 reactors in series is 4 times that of a single CSTR or batch reactor having the same volume. | [27] |
| Ethanol fermentation in 2 CSTRs in series with cell recycling/separator after each stage of reactor. Numerical study using 4 kinetic models. | 2 unequal-size reactors | 2-CSTRs in series with recycling have volume of 25% less than one chemostat and 30% higher ethanol productivity. | [24] |
| Ethanol fermentation by S. cerevisiae on glucose as substrate in continuous stirred-tank reactors. | 3 decreasing volume CSTRs in series | Experimental results confirmed the optimal design predictions for substrate, product and biomass concentrations for each of the 3 unequal-size reactors in series. | [22] |
| Minimum total volume of CSTRs in series based on substrate and product inhibition kinetic model obtained from [2]. | Up to 5 reactors in series. Equal and optimum design. | For feed substrate concentration of 50 g/L and 99% conversion. The percentage reduction in the total volume using the optimum design compared to equal-size reactors is 35%, 54%, 62% and 66% for 2, 3, 4 and 5 reactors in series, respectively. | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abu Reesh, I.M. Optimum Design of N Continuous Stirred-Tank Bioreactors in Series for Fermentation Processes Based on Simultaneous Substrate and Product Inhibition. Processes 2021, 9, 1419. https://doi.org/10.3390/pr9081419
Abu Reesh IM. Optimum Design of N Continuous Stirred-Tank Bioreactors in Series for Fermentation Processes Based on Simultaneous Substrate and Product Inhibition. Processes. 2021; 9(8):1419. https://doi.org/10.3390/pr9081419
Chicago/Turabian StyleAbu Reesh, Ibrahim M. 2021. "Optimum Design of N Continuous Stirred-Tank Bioreactors in Series for Fermentation Processes Based on Simultaneous Substrate and Product Inhibition" Processes 9, no. 8: 1419. https://doi.org/10.3390/pr9081419
APA StyleAbu Reesh, I. M. (2021). Optimum Design of N Continuous Stirred-Tank Bioreactors in Series for Fermentation Processes Based on Simultaneous Substrate and Product Inhibition. Processes, 9(8), 1419. https://doi.org/10.3390/pr9081419