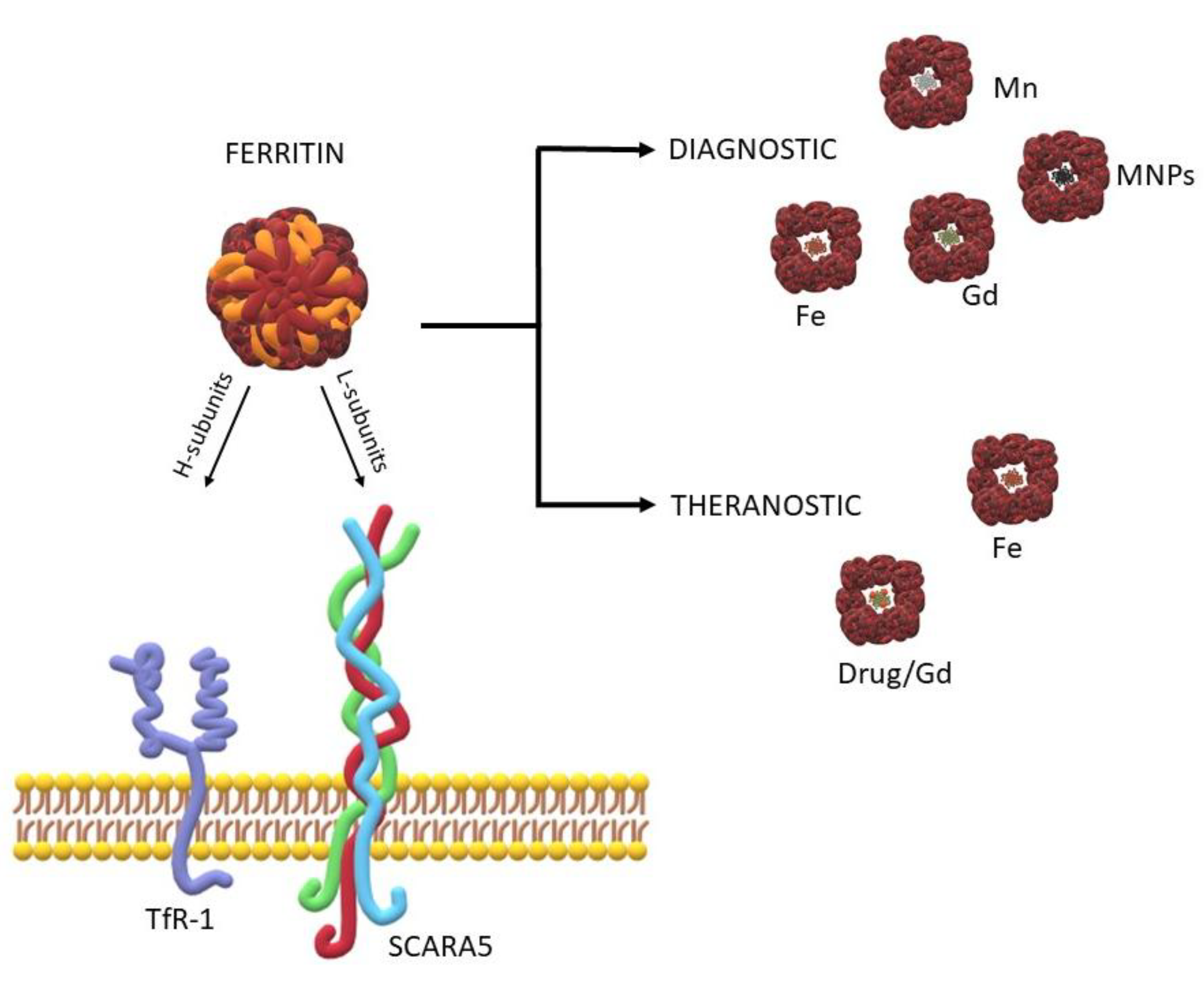
Ferritin: A Platform for MRI Contrast Agents Delivery


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
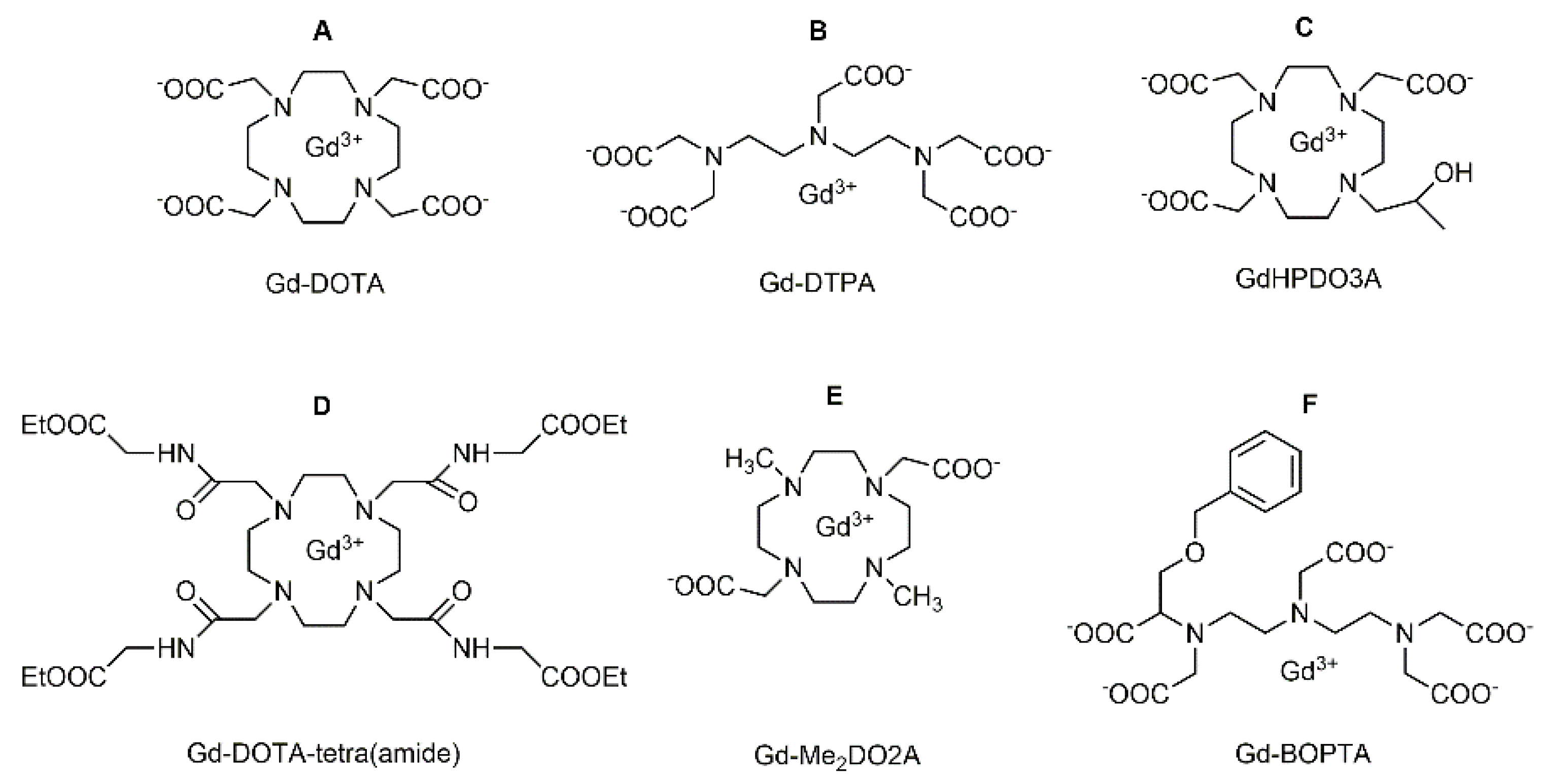
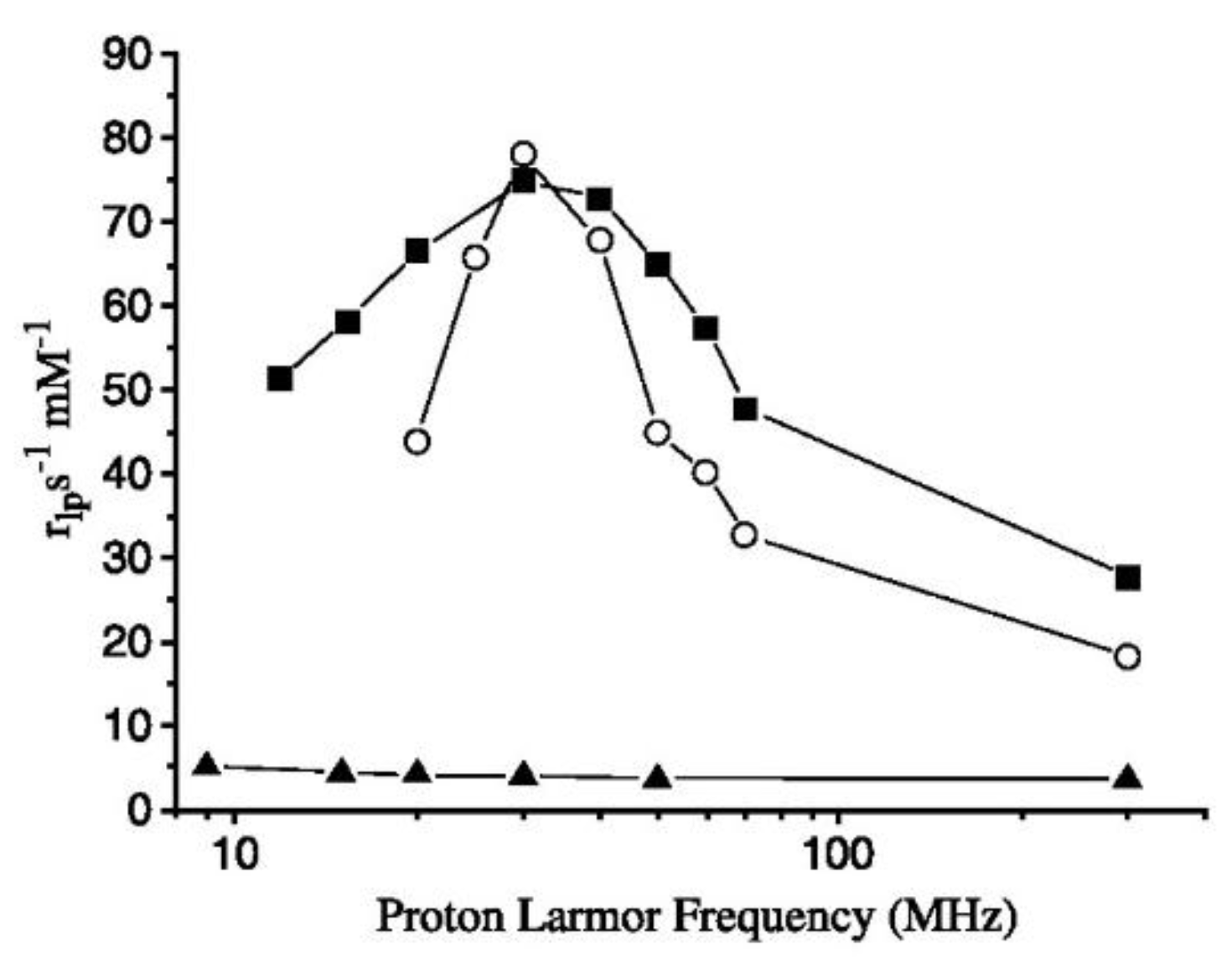
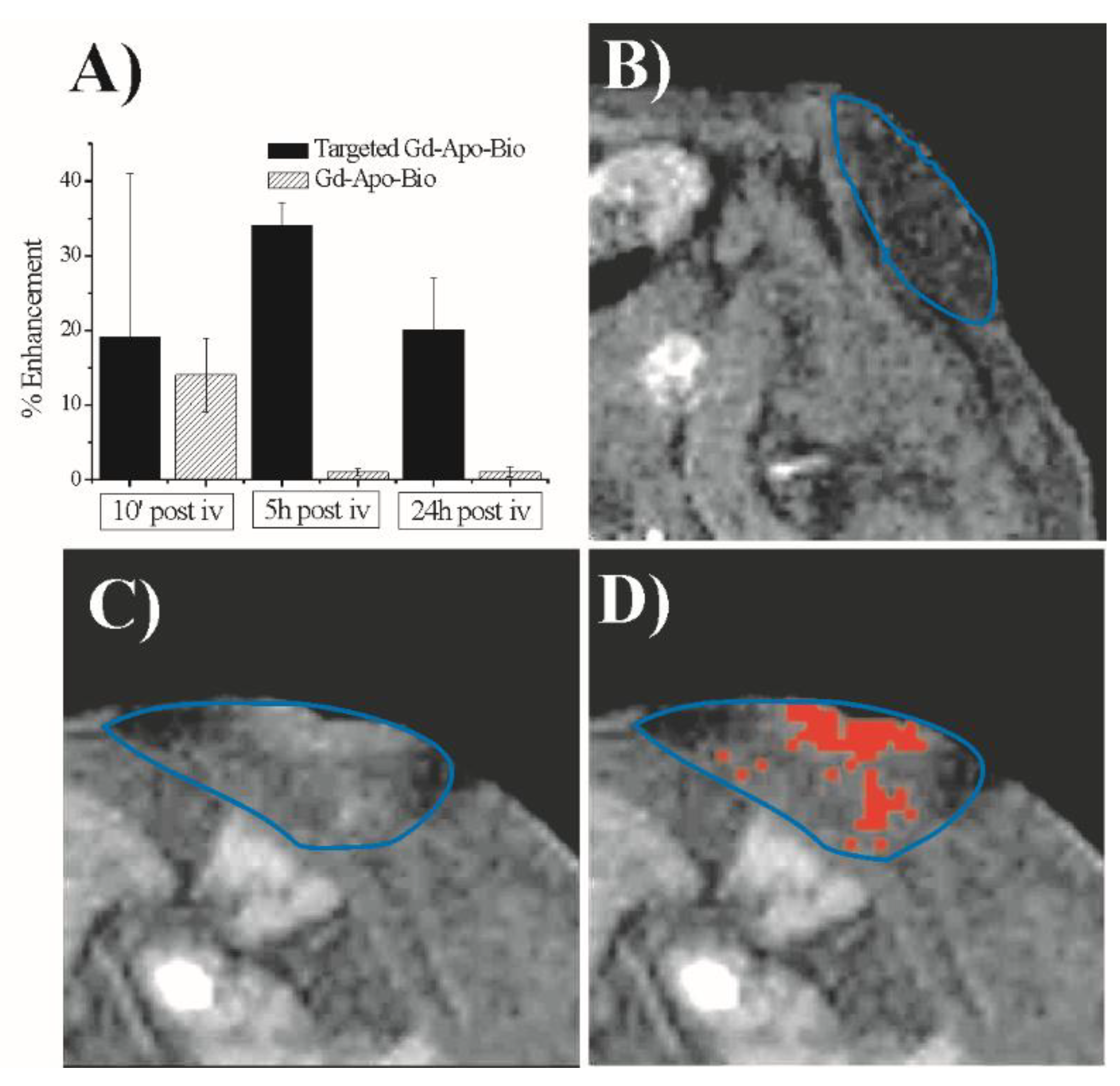
2. Gd-Loaded Apoferritin
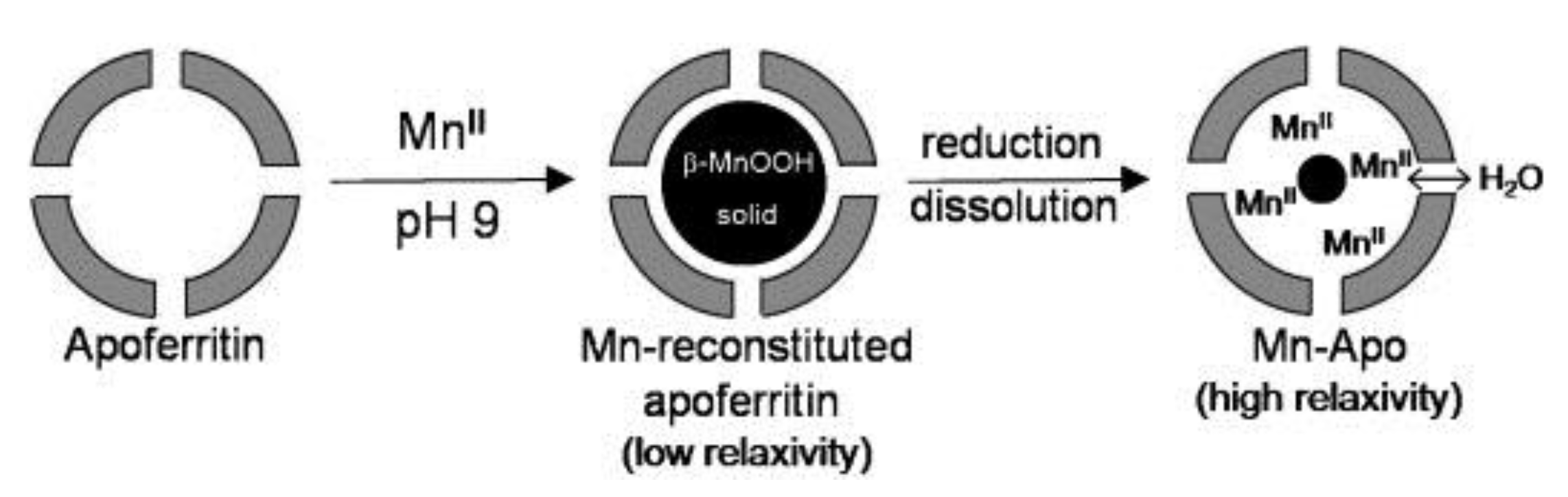
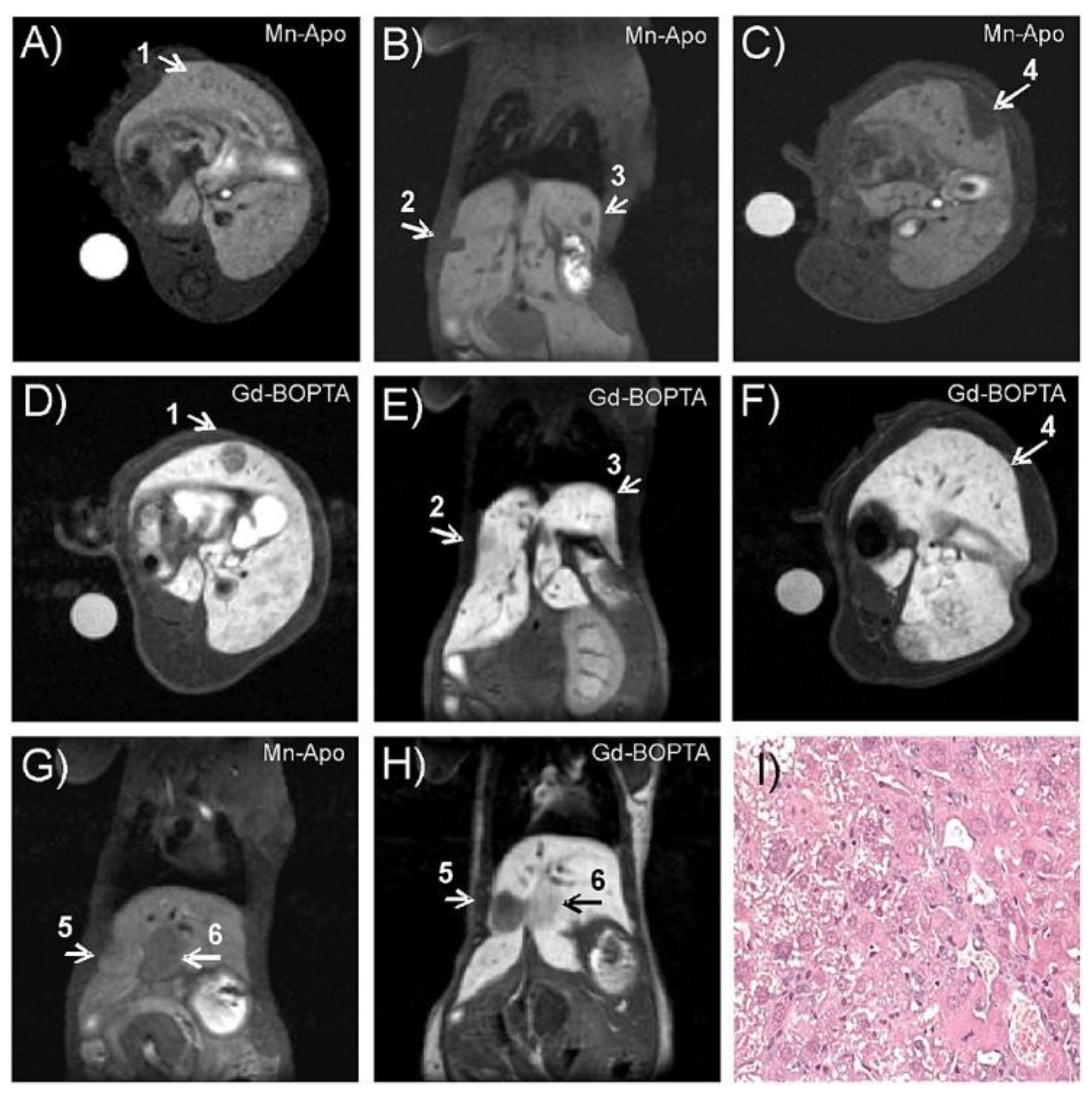
3. Mn-Loaded Apoferritin
4. Magnetoferritin
5. Ferritin as an Endogenous MRI Reporter for Gene Expression Imaging
6. Theranostic Use of Ferritin
7. Conclusions
Funding
Conflicts of Interest
References
- Wahsner, J.; Gale, E.M.; Rodríguez-Rodríguez, A.; Caravan, P. Chemistry of MRI Contrast Agents: Current Challenges and New Frontiers. Chem. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Terreno, E.; Dastrù, W.; Delli Castelli, D.; Gianolio, E.; Geninatti Crich, S.; Longo, D.; Aime, S. Advances in metal-based probes for MR molecular imaging applications. Curr. Med. Chem. 2010, 17, 3684–3700. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, C.S.; Tóth, E. MRI probes for sensing biologically relevant metal ions. Future Med. Chem. 2010, 2, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Fasano, M.; Curry, S.; Terreno, E.; Galliano, M.; Fanali, G.; Narciso, P.; Notari, S.; Ascenzi, P. The extraordinary ligand binding properties of human serum albumin. IUBMB Life 2005, 57, 787–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, D.L.; Arena, F.; Consolino, L.; Minazzi, P.; Geninatti-Crich, S.; Giovenzana, G.B.; Aime, S. Gd-AAZTA-MADEC, an improved blood pool agent for DCE-MRI studies on mice on 1 T scanners. Biomaterials 2016, 75, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaxton, C.S.; Rink, J.S.; Naha, P.C.; Cormode, D.P. Lipoproteins and lipoprotein mimetics for imaging and drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 116–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geninatti Crich, S.; Alberti, D.; Szabo, I.; Deagostino, A.; Toppino, A.; Barge, A.; Ballarini, F.; Bortolussi, S.; Bruschi, P.; Protti, N.; et al. MRI-guided neutron capture therapy by use of a dual gadolinium/boron agent targeted at tumour cells through upregulated low-density lipoprotein transporters. Chem. Eur. J. 2011, 17, 8479–8486. [Google Scholar] [CrossRef] [PubMed]
- Arosio, P.; Ingrassia, R.; Cavadini, P. Ferritins: A family of molecules for iron storage, antioxidation and more. Biochim. Biophys. Acta 2009, 1790, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Chiou, B.; Connor, J.R. Emerging and Dynamic Biomedical Uses of Ferritin. Pharmaceuticals 2018, 11, 124. [Google Scholar] [CrossRef] [PubMed]
- Jutz, G.; van Rijn, P.; Santos, M.B.; Böker, A.P. Ferritin: A versatile building block for bionanotechnology. Chem. Rev. 2015, 115, 1653–1701. [Google Scholar] [CrossRef] [PubMed]
- Theil, E.C. Ferritin: The protein nanocage and iron biomineral in health and in disease. Inorg. Chem. 2013, 52, 12223–12233. [Google Scholar] [CrossRef] [PubMed]
- Todorich, B.; Zhang, X.; Connor, J.R. H-ferritin is the major source of iron for oligodendrocytes. Glia 2011, 59, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.C.; Chau, L.A. Uptake of ferritin by isolated rat hepatocytes. Effect of metabolic inhibitors and iron. Clin. Investig. Med. Méd. Clin. Exp. 1993, 16, 15–21. [Google Scholar]
- Blight, G.D.; Morgan, E.H. Ferritin and iron uptake by reticulocytes. Br. J. Haematol. 1983, 55, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Moss, D.; Powell, L.W.; Arosio, P.; Halliday, J.W. Characterization of the ferritin receptors of human T lymphoid (MOLT-4) cells. J. Lab. Clin. Med. 1992, 119, 273–279. [Google Scholar] [PubMed]
- Sakamoto, S.; Kawabata, H.; Masuda, T.; Uchiyama, T.; Mizumoto, C.; Ohmori, K.; Koeffler, H.P.; Kadowaki, N.; Takaori-KondoH, A. H-Ferritin is Preferentially Incorporated by Human Erythroid Cells through Transferrin Receptor 1 in a Threshold-Dependent Manner. PLoS ONE 2015, 10, e0139915. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Paragas, N.; Ned, R.M.; Qiu, A.; Viltard, M.; Leete, T.; Drexler, I.R.; Chen, X.; Sanna-Cherchi, S.; Mohammed, F.; et al. Scara5 is a ferritin receptor mediating non-transferrin iron delivery. Dev. Cell 2009, 16, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Seaman, W.E.; Di, X.; Wang, W.; Willingham, M.; Torti, F.M.; Torti, S.V. Iron uptake mediated by binding of H-ferritin to the TIM-2 receptor in mouse cells. PLoS ONE 2011, 6, e23800. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Fang, J.C.; Ryan, J.C.; Niemi, E.C.; Lebrón, J.A.; Björkman, P.J.; Arase, H.; Torti, F.M.; Torti, S.V.; Nakamura, M.C.; et al. Binding and uptake of H-ferritin are mediated by human transferrin receptor-1. Proc. Natl. Acad. Sci. USA 2010, 107, 3505–3510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhang, L.; Knez, M. Comparison of two endogenous delivery agents in cancer therapy: Exosomes and ferritin. Pharmacol. Res. 2016, 110, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Knovich, M.A.; Storey, J.A.; Coffman, L.G.; Torti, S.V. Ferritin for the Clinician. Blood Rev. 2009, 23, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, X.; Dong, D.; Zhang, B.; Xue, Y.; Shang, P. Transferrin receptor 1 in cancer: A new sight for cancer therapy. Am. J. Cancer Res. 2018, 8, 916–931. [Google Scholar] [PubMed]
- Huang, J.; Zheng, D.L.; Qin, F.S.; Cheng, N.; Chen, H.; Wan, B.B.; Wang, Y.P.; Xiao, H.S.; Han, Z.G. Genetic and epigenetic silencing of SCARA5 may contribute to human hepatocellular carcinoma by activating FAK signaling. J. Clin. Investig. 2010, 120, 223–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, K.; Su, F.; Liu, L.; Lv, X.; Zhang, J.; Zhang, Y.; Liu, B. SCARA5 plays a critical role in the progression and metastasis of breast cancer by inactivating the ERK1/2, STAT3, and AKT signaling pathways. Mol. Cell Biochem. 2017, 435, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Khoshnejad, M.; Parhiza, H.; Shuvaeva, V.V.; Dmochowskib, I.J.; Muzykantova, V.R. Ferritin-based drug delivery systems: Hybrid nanocarriers for vascular immunotargeting. J. Control. Release 2018, 282, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Khoshnejad, M.; Greineder, C.F.; Pulsipher, K.W.; Villa, C.H.; Altun, B.; Pan, D.C.; Tsourkas, A.; Dmochowski, I.J.; Muzykantov, V.R. Ferritin Nanocages with Biologically Orthogonal Conjugation for Vascular Targeting and Imaging. Bioconj. Chem. 2018, 29, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Aime, S.; Cabella, C.; Colombatto, S.; Geninatti Crich, S.; Gianolio, E.; Maggioni, F. Insights into the use of paramagnetic Gd(III) complexes in MR-molecular imaging investigations. J. Magn. Reson. Imaging 2002, 16, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Chasteen, N.D. Molecular diffusion into horse spleen ferritin: A nitroxide radical spin probe study. Biophys. J. 1996, 71, 1587–1595. [Google Scholar] [CrossRef]
- Belletti, D.; Pederzoli, F.; Forni, F.; Vandelli, M.A.; Tosi, G.; Ruozi, B. Protein cage nanostructure as drug delivery system: Magnifying glass on apoferritin. Expert Opin. Drug Deliv. 2017, 14, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Aime, S.; Frullano, L.; Geninatti Crich, S. Compartmentalization of a gadolinium complex in the apoferritin cavity: A route to obtain high relaxivity contrast agents for magnetic resonance imaging. Angew. Chem. Int. Ed. Engl. 2002, 41, 1017–1019. [Google Scholar] [CrossRef]
- Vasalatiy, O.; Zhao, P.; Zhang, S.; Aime, S.; Sherry, A.D. Catalytic effects of apoferritin interior surface residues on water proton exchange in lanthanide complexes. Contrast Media Mol. Imaging 2006, 1, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fries, P.H.; Belorizky, E. Enhancement of the water proton relaxivity by trapping Gd3+ complexes in nanovesicles. J. Chem. Phys. 2010, 133, 024504. [Google Scholar] [CrossRef] [PubMed]
- Makino, A.; Harada, H.; Okada, T.; Kimura, H.; Amano, H.; Saji, H.; Hiraoka, M.; Kimura, S. Effective encapsulation of a new cationic gadolinium chelates into apoferritin and its evaluation as an MRI contrast agent. Nanomedicine 2011, 7, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, P.; Valero, E.; Gálvez, N.; Domínguez-Vera, J.M.; Marinone, M.; Poletti, G.; Corti, M.; Lascialfari, A. MRI relaxation properties of water-soluble apoferritin-encapsulated gadolinium oxide-hydroxide nanoparticles. Dalton Trans. 2009, 5, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Geninatti Crich, S.; Bussolati, B.; Tei, L.; Grange, C.; Esposito, G.; Lanzardo, S.; Camussi, G.; Aime, S. Magnetic resonance visualization of tumor angiogenesis by targeting neural cell adhesion molecules with the highly sensitive gadolinium-loaded apoferritin probe. Cancer Res. 2006, 66, 9196–9201. [Google Scholar] [CrossRef] [PubMed]
- Thakral, C.; Alhariri, J.; Abraham, J.L. Long-term retention of gadolinium in tissues from nephrogenic systemic fibrosis patient after multiple gadolinium-enhanced MRI scans: Case report and implications. Contrast Media Mol. Imaging 2007, 2, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Caruthers, S.D.; Senpan, A.; Schmieder, A.H.; Wickline, S.A.; Lanza, G.M. Revisiting an old friend: Manganese-based MRI contrast agents. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2011, 3, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Kálmán, F.K.; Geninatti-Crich, S.; Aime, S. Reduction/dissolution of a beta-MnOOH nanophase in the ferritin cavity to yield a highly sensitive, biologically compatible magnetic resonance imaging agent. Angew. Chem. Int. Ed. Engl. 2010, 49, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, F.C.; Douglas, T.; Levi, S.; Arosio, P.; Mann, S. Reconstitution of manganese oxide cores in horse spleen and recombinant ferritins. J. Inorg. Biochem. 1995, 58, 59–68. [Google Scholar] [CrossRef]
- Geninatti Crich, S.; Cutrin, J.C.; Lanzardo, S.; Conti, L.; Kálmán, F.K.; Szabó, I.; Lago, N.R.; Iolascon, A.; Aime, S. Mn-loaded apoferritin: A highly sensitive MRI imaging probe for the detection and characterization of hepatocarcinoma lesions in a transgenic mouse model. Contrast Media Mol. Imaging 2012, 7, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Geninatti Crich, S.; Cadenazzi, M.; Lanzardo, S.; Conti, L.; Ruiu, R.; Alberti, D.; Cavallo, F.; Cutrin, J.C.; Aime, S. Targeting ferritin receptors for the selective delivery of imaging and therapeutic agents to breast cancer cells. Nanoscale 2015, 7, 6527–6533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, I.; Geninatti Crich, S.; Alberti, D.; Kálmán, F.K.; Aime, S. Mn loaded apoferritin as an MRI sensor of melanin formation in melanoma cells. Chem. Commun. (Camb.) 2012, 48, 2436–2438. [Google Scholar] [CrossRef] [PubMed]
- Patrick, P.S.; Rodrigues, T.B.; Kettunen, M.I.; Lyons, S.K.; Neves, A.A.; Brindle, K.M. Development of Timd2 as a reporter gene for MRI. Magn. Reson. Med. 2016, 75, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Sana, B.; Poh, C.L.; Lim, S. A manganese-ferritin nanocomposite as an ultrasensitive T2 contrast agent. Chem. Commun. (Camb.) 2012, 48, 862–864. [Google Scholar] [CrossRef] [PubMed]
- Hocq, A.; Luhmer, M.; Saussez, S.; Louryan, S.; Gillis, P.; Gossuin, Y. Effect of magnetic field and iron content on NMR proton relaxation of liver, spleen and brain tissues. Contrast Media Mol. Imaging 2014, 10, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Gossuin, Y.; Muller, R.N.; Gillis, P.; Bartel, L. Relaxivities of human liver and spleen ferritin. Magn. Reson. Imaging 2005, 23, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Forge, D.; Port, M.; Roch, A.; Robic, C.; Vander Elst, L.; Muller, R.N. Magnetic iron oxide nanoparticles: Synthesis, stabilization, vectorization, physicochemical characterizations, and biological applications. Chem. Rev. 2008, 108, 2064–2110. [Google Scholar] [CrossRef] [PubMed]
- Unterweger, H.; Dézsi, L.; Matuszak, J.; Janko, C.; Poettler, M.; Jordan, J.; Bäuerle, T.; Szebeni, J.; Fey, T.; Boccaccini, A.; et al. Dextran-coated superparamagnetic iron oxide nanoparticles for magnetic resonance imaging: Evaluation of size-dependent imaging properties, storage stability and safety. Int. J. Nanomed. 2018, 13, 1899–1915. [Google Scholar] [CrossRef] [PubMed]
- López-Castro, J.D.; Maraloiu, A.V.; Delgado, J.J.; Calvino, J.J.; Blanchin, M.G.; Gálvez, N.; Domínguez-Vera, J.M. From synthetic to natural nanoparticle: Monitoring the biodegradation of SPIO into ferritin by electron microscopy. Nanoscale 2011, 3, 4597–4599. [Google Scholar] [CrossRef] [PubMed]
- Valero, E.; Tambalo, S.; Marzola, P.; Ortega-Muñoz, M.; López-Jaramillo, F.J.; Santoyo-González, F.; de Dios López, J.; Delgado, J.J.; Calvino, J.J.; Cuesta, R.; et al. Magnetic Nanoparticles-Templated Assembly of Protein Subunits: A New Platform for Carbohydrate-Based MRI. Nanoprobes J. Am. Chem. Soc. 2011, 133, 4889–4895. [Google Scholar] [CrossRef] [PubMed]
- Masaki, U.; Masahiro, T.; Cunningham, C.H.; Suzuki, Y.; Willits, D.A.; Willis, A.F.; Yang, P.C.; Tsao, P.S.; McConnell, M.C.; Young, M.J.; et al. A Human Ferritin Iron Oxide Nano-composite Magnetic Resonance Contrast Agent. Magn. Reson. Med. 2008, 60, 1073–1081. [Google Scholar]
- Cai, Y.; Wang, Y.; Xua, H.; Cao, C.; Zhue, R.; Tang, X.; Zhang, T.; Pan, Y. Positive Magnetic Resonance Angiography by Ultrafine Ferritin-based Iron Oxide Nanoparticles. Nanoscale 2018. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Ziv, K.; Plaks, V.; Harmelin, A.; Neeman, M. Ferritin nanoparticles as magnetic resonance reporter gene. Nanomed. Nanobiotechnol. 2009, 1, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Dafni, H.; Meir, G.; Harmelin, A.; Neeman, M. Ferritin as an endogenous MRI reporter for noninvasive imaging of gene expression in C6 glioma tumors. Neoplasia 2005, 7, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Vandsburger, M.H.; Radoul, M.; addadi, Y.; Mpofu, S.; Cohen, B.; Eliam, R.; Neeman, M. Quantitative bioexponential MR imaging relaxometry reveals the dynamic recruitment of ferritin-expressing fibroblasts to the angiogenic rim of tumors. Radiology 2013, 268, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewska, A.; Nowakowski, A.; Janowski, M.; Bulte, J.W.; Gilad, A.A.; Walczak, P.; Lukomska, B. Pre- and postmortem imaging of transplanted cells. Int. J. Nanomed. 2015, 10, 5543–5559. [Google Scholar]
- Iordanova, B.; Ahrens, E.T. In vivo magnetic resonance imaging of ferritin-based reporter visualizes native neuroblast migration. Neuroimage 2012, 59, 1004–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Cai, J.; Liu, B.; Zhong, Y.; Qin, Y. Cellular magnetic resonance imaging contrast generated by the ferritin heavy chain genetic reporter under the control of a Tet-On switch. Stem Cell Res. Ther. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; He, R.; Zhang, Y.; Wu, R.; Xiao, Y. Adenoviral vector mediated ferritin over-expression in mesenchymal stem cells detected by 7T MRI in vitro. PLoS ONE 2017, 12, e0185260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Duan, X.; Lu, L.; Zhang, X.; Chen, M.; Mao, J.; Cao, M.; Shen, J. In vivo long-term tracking of neuronal stem cells transplanted into an acute ischemic stroke model with reporter gene-based bimodal MR and optical imaging. Cell Transp. 2017, 26, 1648–1662. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Ono, K.; Suzuki, H.; Eguchi, M.; Yamaguchi, S.; Sawada, M. Visualization of Arc promoter-driven neuronal activity by magnetic resonance imaging. Neurosci. Lett. 2017, 666, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Noury, M.; Lopez, J. Nanomedicine and personalised medicine: Understanding the personalisation of health care in the molecular era. Sociol. Health Illn. 2016, 20, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Mankoff, D.A. A definition of Molecular Imaging. J. Nucl. Med. 2007, 48, 18–21. [Google Scholar]
- Lammers, T.; Aime, S.; Hennink, W.E.; Storm, G.; Kiessling, F. Theranostic Nanomedicine. Acc. Chem. Res. 2011, 44, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Caruthers, S.D.; Wickline, S.A.; Lanza, G.M. Nanotechnological applications in medicine. Curr. Opin. Biotechnol. 2007, 18, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Lammers, T.; Hennink, W.E.; Storm, G. Tumour-targeted nanomedicines: Principles and practice. Br. J. Cancer 2008, 99, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Truffi, M.; Fiandra, L.; Sorrentino, L.; Monieri, M.; Corsi, F.; Mazzucchelli, S. Ferritin nanocages: A biological platform for drug delivery, imaging and theranostics in cancer. Pharmacol. Res. 2016, 107, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Mazzucchelli, S.; Mazzucchelli, S.; Bellini, M.; Fiandra, L.; Truffi, M.; Rizzuto, M.A.; Sorrentino, L.; Longhi, E.; Nebuloni, M.; Prosperi, D.; et al. Nanometronomic treatment of 4T1 breast cancer with nanocaged doxorubicin prevents drug resistance and circumvents cardiotoxicity. Oncotarget 2017, 8, 8383–8396. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Tang, W.; Chen, H.; Lin, X.; Todd, T.; Wang, G.; Cowger, T.; Chen, X.; Xie, J. RGD-modified nanoparticles for efficient drug delivery to tumors. ACS Nano 2013, 7, 4830–4837. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, X.; Diao, H.; Zhang, J.; Li, H.; Sun, H.; Guo, Z. Encapsulation of platinum anticancer drugs by apoferritin. Chem. Commun. 2007, 33, 3453–3455. [Google Scholar] [CrossRef] [PubMed]
- Falvo, E.; Tremante, E.; Fraioli, R.; Leonetti, C.; Zamparelli, C.; Boffi, A.; Morea, V.; Ceci, P.; Giacomini, P. Antibody-drug conjugates: Targeting melanoma with cisplatin encapsulated in protein-cage nanoparticles based on human ferritin. Nanoscale 2013, 5, 12278–12285. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, G.; Pica, A.; Petruk, G.; Pane, F.; Amoresano, A.; Cilibrizzi, A.; Vilar, R.; Monti, D.M.; Merlino, A. Preparation, structure, cytotoxicity and mechanism of action of ferritin-Pt(II) terpyridine compound nanocomposites. Nanomedicine (Lond.) 2018. [Google Scholar] [CrossRef] [PubMed]
- Cutrin, J.C.; Geninatti Crich, S.; Burghelea, D.; Dastrù, W.; Aime, S. Curcumin/Gd Loaded Apoferritin: A Novel “Theranostic” Agent To Prevent Hepatocellular Damage in Toxic Induced Acute Hepatitis. Mol. Pharm. 2013, 10, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Laura Conti, L.; Lanzardo, S.; Ruiu, R.; Cadenazzi, M.; Cavallo, F.; Aime, S.; Geninatti Crich, S. L-Ferritin targets breast cancer stem cells and delivers therapeutic and imaging agents. Oncotarget 2016, 7, 66713–66727. [Google Scholar] [Green Version]
- Frank, N.Y.; Schatton, T.; Frank, M.H. The therapeutic promise of the cancer stem cell concept. J. Clin. Investig. 2010, 120, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turino, L.N.; Ruggiero, M.R.; Stefanìa, R.; Cutrin, J.C.; Aime, S.; Geninatti Crich, S. Ferritin Decorated PLGA/Paclitaxel Loaded Nanoparticles Endowed with an Enhanced Toxicity Toward MCF-7 Breast Tumor Cells. Bioconj. Chem. 2017, 28, 1283–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutrin, J.C.; Alberti, D.; Bernacchioni, C.; Ciambellotti, S.; Turano, P.; Luchinat, C.; Geninatti-Crich, S.; Aime, S. Cancer cell death induced by ferritins and the peculiar role of their labile iron pool. Oncotarget 2018, 9, 27974–27984. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.V.; Torti, F.M. Iron and cancer: More ore to be mined. Nat. Rev. Cancer 2013, 342, 342–355. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruggiero, M.R.; Alberti, D.; Bitonto, V.; Geninatti Crich, S. Ferritin: A Platform for MRI Contrast Agents Delivery. Inorganics 2019, 7, 33. https://doi.org/10.3390/inorganics7030033
Ruggiero MR, Alberti D, Bitonto V, Geninatti Crich S. Ferritin: A Platform for MRI Contrast Agents Delivery. Inorganics. 2019; 7(3):33. https://doi.org/10.3390/inorganics7030033
Chicago/Turabian StyleRuggiero, Maria Rosaria, Diego Alberti, Valeria Bitonto, and Simonetta Geninatti Crich. 2019. "Ferritin: A Platform for MRI Contrast Agents Delivery" Inorganics 7, no. 3: 33. https://doi.org/10.3390/inorganics7030033
APA StyleRuggiero, M. R., Alberti, D., Bitonto, V., & Geninatti Crich, S. (2019). Ferritin: A Platform for MRI Contrast Agents Delivery. Inorganics, 7(3), 33. https://doi.org/10.3390/inorganics7030033