
The Role of Bacterial, Dentinal, Salivary, and Neutrophil Degradative Activity in Caries Pathogenesis




Abstract
:
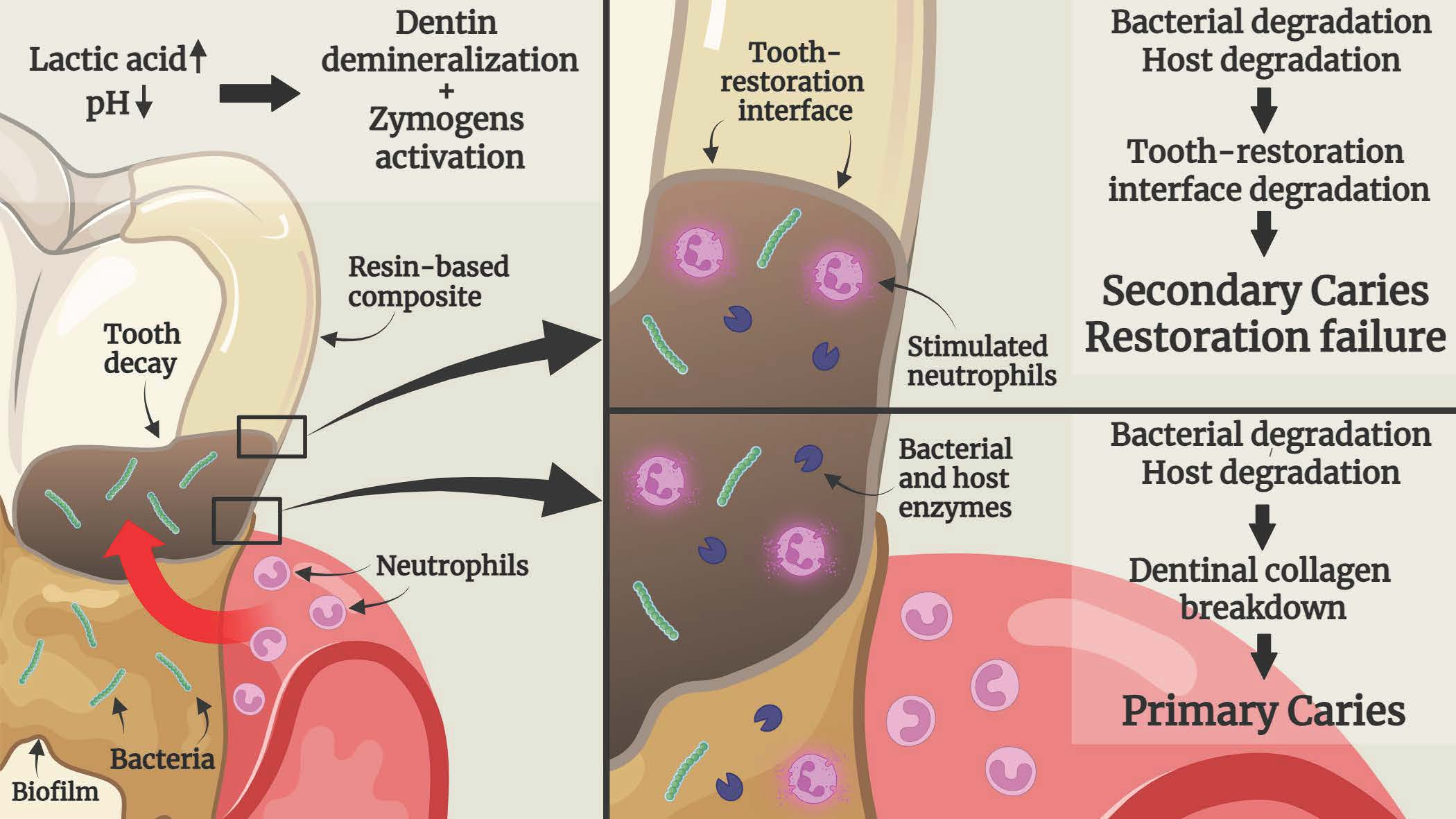{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
3. Caries Classifications
3.1. Primary Caries: Coronal and Root Caries
3.2. Secondary (Recurrent) Caries
4. Bacterial, Dentinal, and Salivary Dentin Degradative Activity
4.1. Bacterial Degradative Activity towards Dentin
4.2. Dentinal Endogenous Degradative Activity towards Dentin
4.3. Salivary Degradative Activity towards Dentin
5. Immune System Degradative Activity
5.1. Neutrophils
5.1.1. Neutrophils’ Degradative Activity in Periodontitis
5.1.2. Neutrophils’ Degradative Activity towards Dentin
6. Resin-based Composites
6.1. RBC Chemistry and Composition
6.2. The Adhesive System
6.3. The Hybrid Layer
6.4. RBC Degradation
6.4.1. Bacterial Degradative Activity towards RBC
6.4.2. Salivary Degradative Activity towards RBC
6.4.3. Neutrophils Degradative Activity towards RBC
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. Dental caries. Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef]
- Van Chuyen, N.; Van Du, V.; Son, H.A. The prevalence of dental caries and associated factors among secondary school children in rural highland Vietnam. BMC Oral Health 2021, 21, 1–7. [Google Scholar] [CrossRef]
- Bagramian, R.A.; Garcia-Godoy, F.; Volpe, A.R. The global increase in dental caries. A pending public health crisis. Am. J. dent 2009, 22, 3–8. [Google Scholar]
- US Department of Health and Human Services Oral Health Coordinating Committee. US Department of Health and Human Services oral health strategic framework, 2014–2017. Public Health Rep. 2016, 131, 242–257. [Google Scholar] [CrossRef]
- Ferro, K.J.; Morgano, S.M. The Glossary of Prosthodontic Terms: Ninth Edition. J. Prosthet. Dent. 2017, 117, e1–e105. [Google Scholar] [CrossRef]
- Aqawi, M.; Sionov, R.V.; Steinberg, D. Anti-bacterial properties of cannabigerol toward Streptococcus mutans. Front. Microbiol. 2021, 12, 922. [Google Scholar] [CrossRef]
- Boisen, G.; Davies, J.R.; Neilands, J. Acid tolerance in early colonizers of oral biofilms. BMC Microbiol. 2021, 21, 1–9. [Google Scholar] [CrossRef]
- Takahashi, N.; Nyvad, B. Caries ecology revisited: Microbial dynamics and the caries process. Caries Res. 2008, 42, 409–418. [Google Scholar] [CrossRef]
- Kidd, E.A.M.; Fejerskov, O. What constitutes dental caries? Histopathology of carious enamel and dentin related to the action of cariogenic biofilms. J. Dent. Res. 2004, 83, 35–38. [Google Scholar] [CrossRef]
- Featherstone, J.D.B. The continuum of dental caries—evidence for a dynamic disease process. J. Dent. Res. 2004, 83, 39–42. [Google Scholar] [CrossRef]
- Mazzoni, A.; Tjäderhane, L.; Breschi, L. Role of dentin MMPs in caries progression and bond stability. J. Dent. Res. 2014, 94, 241–251. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef]
- Boukpessi, T.; Menashi, S.; Chaussain, C. Endogenous enzymes in root caries. Karger Publ. 2017, 26, 35–42. [Google Scholar] [CrossRef]
- Finer, Y.; Jaffer, F.; Santerre, J.P. Mutual influence of cholesterol esterase and pseudocholinesterase on the biodegradation of dental composites. Biomaterials 2003, 25, 1787–1793. [Google Scholar] [CrossRef]
- Liu, Y.; Tjäderhane, L.; Tay, F.R. Limitations in bonding to dentin and experimental strategies to prevent bond degradation. J. Dent. Res. 2011, 90, 953–968. [Google Scholar] [CrossRef]
- Santerre, J.P.; Shajii, L.; Leung, B.W. Relation of dental composite formulations to their degradation and the release of hydrolyzed polymeric-resin-derived products. Crit. Rev. Oral Biol. Med. 2001, 12, 136–151. [Google Scholar] [CrossRef]
- Shokati, B.; Tam, L.E.; Santerre, J.P.; Finer, Y. Effect of salivary esterase on the integrity and fracture toughness of the dentin-resin interface. J. Biomed. Mater. Res. Part B Appl. Biomater. 2010, 94, 230–237. [Google Scholar] [CrossRef]
- Gitalis, R.; Zhou, L.; Finer, Y. Human neutrophils degrade methacrylate resin composites and tooth dentin. Acta Biomater. 2019, 88, 325–331. [Google Scholar] [CrossRef]
- Gitalis, R.; Bae, J.H.; Finer, Y. Human neutrophils compromise the restoration-tooth interface. Acta Biomater. 2020, 117, 283–293. [Google Scholar] [CrossRef]
- Hajishengallis, G. New developments in neutrophil biology and Periodontitis. Periodontology 2000 2020, 82, 78–92. [Google Scholar] [CrossRef]
- Shen, S.; Samaranayake, L.P.; Yip, H.K. In vitro growth, acidogenicity and cariogenicity of predominant human root caries flora. J. Dent. 2004, 32, 667–678. [Google Scholar] [CrossRef] [PubMed]
- AlQranei, M.S.; Balhaddad, A.A.; Melo, M.A.S. The burden of root caries: Updated perspectives and advances on management strategies. Gerodontology 2021, 38, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Askar, H.; Krois, J.; Schwendicke, F. Secondary caries: What is it, and how it can be controlled, detected, and managed? Clin. Oral Investig. 2020, 24, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Mjör, I.A.; Toffentti, F. Secondary caries: A literature review with case reports. Quintessence Int. 2000, 31, 165–179. [Google Scholar]
- Ferracane, J.L. Models of caries formation around dental composite restorations. J. Dent. Res. 2017, 96, 364–371. [Google Scholar] [CrossRef]
- Jokstad, A. Secondary caries and microleakage. Dent. Mater. 2016, 32, 11–25. [Google Scholar] [CrossRef]
- Schwendicke, F.; Kern, M.; Paris, S. Marginal integrity and secondary caries of selectively excavated teeth in vitro. J. Dent. 2014, 42, 1261–1268. [Google Scholar] [CrossRef]
- Campos, P.E.G.A.; Barceleiro, M.O.; Martins, L.R.M. Evaluation of the cervical integrity during occlusal loading of class II restorations. Oper. Dent. 2008, 33, 59–64. [Google Scholar] [CrossRef]
- Marashdeh, M.Q.; Gitalis, R.; Finer, Y. Endodontic pathogens possess collagenolytic properties that degrade human dentine collagen matrix. Int. Endod. J. 2019, 52, 416–423. [Google Scholar] [CrossRef]
- Huang, B.; Stewart, C.A.; McCulloch, C.A.; Santerre, J.P.; Cvitkovitch, D.G.; Finer, Y. Streptococcus mutans Proteases Degrade Dentinal Collagen. Dent. J. 2022, 10, 223. [Google Scholar] [CrossRef]
- Tezvergil-Mutluay, A.; Mutluay, M.; Pashley, D.H. Effect of phosphoric acid on the degradation of human dentin matrix. J. Dent. Res. 2013, 92, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Buzalaf, M.A.R.; Hannas, A.R.; Delbem, A.C.B. pH-cycling models for in vitro evaluation of the efficacy of fluoridated dentifrices for caries control: Strengths and limitations. J. Appl. Oral Sci. 2010, 18, 316–334. [Google Scholar] [CrossRef] [PubMed]
- Ashwini, A.; Dineshkumar, T.; Rajkumar, K. Dentin degradonomics–The potential role of salivary MMP-8 in dentin caries. J. Clin. Exp. Dent. 2020, 12, e108–e115. [Google Scholar] [CrossRef]
- Hedenbjörk-Lager, A.; Bjørndal, L.; Ericson, D. Caries correlates strongly with salivary levels of matrix metalloproteinase-8. Caries Res. 2015, 49, 1–8. [Google Scholar] [CrossRef]
- Charadram, N.; Farahani, R.M.; Hunter, N. Regulation of reactionary dentin formation by odontoblasts in response to polymicrobial invasion of dentin matrix. Bone 2012, 50, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Toledano, M.; Nieto-Aguilar, R.; Alaminos, M. Differential expression of matrix metalloproteinase-2 in human coronal and radicular sound and carious dentine. J. Dent. 2010, 38, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Boukpessi, T.; Menashi, S.; Chaussain-Miller, C. The effect of stromelysin-1 (MMP-3) on non-collagenous extracellular matrix proteins of demineralized dentin and the adhesive properties of restorative resins. Biomaterials 2008, 29, 4367–4373. [Google Scholar] [CrossRef]
- Nascimento, F.D.; Minciotti, C.L.; Tersariol, I.L.S. Cysteine cathepsins in human carious dentin. J. Dent. Res. 2011, 90, 506–511. [Google Scholar] [CrossRef]
- Gursoy, U.K.; Könönen, E.; Sorsa, T. Salivary MMP-8, TIMP-1, and ICTP as markers of advanced periodontitis. J. Clin. Periodontol. 2010, 37, 487–493. [Google Scholar] [CrossRef]
- Mei, M.L.; Ito, L.; Lo, E.C. The inhibitory effects of silver diamine fluorides on cysteine cathepsins. J. Dent. 2014, 42, 329–335. [Google Scholar] [CrossRef]
- Liew, P.X.; Kubes, P. The neutrophil’s role during health and disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Cassatella, M.A.; Östberg, N.K.; Soehnlein, O. Biological roles of neutrophil-derived granule proteins and cytokines. Trends Immunol. 2019, 40, 648–664. [Google Scholar] [CrossRef] [PubMed]
- Phillipson, M.; Kubes, P. The healing power of neutrophils. Trends Immunol. 2019, 40, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Kovtun, A.; Messerer, D.A.C.; Ignatius, A. Neutrophils in tissue trauma of the skin, bone, and lung: Two sides of the same coin. J. Immunol. Res. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Papapanou, P.N.; Sanz, M.; Tonetti, M.S. Periodontitis: Consensus report of workgroup 2 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Periodontol. 2018, 89, S173–S182. [Google Scholar] [CrossRef]
- Nicu, E.A.; Loos, B.G. Polymorphonuclear neutrophils in periodontitis and their possible modulation as a therapeutic approach. Periodontology 2000 2016, 71, 140–163. [Google Scholar] [CrossRef]
- Vitkov, L.; Muñoz, L.E.; Hannig, M. Neutrophils orchestrate the periodontal pocket. Front. Immunol. 2021, 12, 788766. [Google Scholar] [CrossRef]
- Jiang, Q.; Zhao, Y.; Li, M. Interactions between neutrophils and periodontal pathogens in late-onset periodontitis. Front. Cell. Infect. Microbiol. 2021, 11, 627328. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef]
- Maekawa, T.; Krauss, J.L.; Hajishengallis, G. Porphyromonas gingivalis manipulates complement and TLR signaling to uncouple bacterial clearance from inflammation and promote dysbiosis. Cell Host Microbe 2014, 15, 768–778. [Google Scholar] [CrossRef]
- Beklen, A.; Tüter, G.; Konttinen, Y.T. Gingival tissue and crevicular fluid co-operation in adult periodontitis. J. Dent. Res. 2006, 85, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.; Gamonal, J.; Sorsa, T. Associations between matrix metalloproteinase-8 and-14 and myeloperoxidase in gingival crevicular fluid from subjects with progressive chronic periodontitis: A longitudinal study. J. Periodontol. 2010, 81, 1644–1652. [Google Scholar] [CrossRef]
- Hirschfeld, J. Dynamic interactions of neutrophils and biofilms. J. Oral Microbiol. 2014, 6, 26102. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.J.; Nannuru, K.C.; Singh, R.K. Cathepsin G–Mediated Activation of Pro–Matrix Metalloproteinase 9 at the Tumor-Bone Interface Promotes Transforming Growth Factor-β Signaling and Bone Destruction. Mol. Cancer Res. 2009, 7, 1224–1233. [Google Scholar] [CrossRef]
- Candido, S.; Abrams, S.L.; McCubrey, J.A. Roles of NGAL and MMP-9 in the tumor microenvironment and sensitivity to targeted therapy. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 438–448. [Google Scholar] [CrossRef]
- Beazoglou, T.; Eklund, S.; Bailit, H. The economic impact of regulating the use of amalgam restorations. Public Heal. Rep. 2007, 122, 657–663. [Google Scholar] [CrossRef]
- Cadenaro, M.; Josic, U.; Mazzoni, A. Progress in dental adhesive materials. J. Dent. Res. 2023, 102, 254–262. [Google Scholar] [CrossRef]
- Ferracane, J.L. Resin composite—state of the art. Dent. Mater. 2011, 27, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Ilie, N.; Hickel, R. Resin composite restorative materials. Aust. Dent. J. 2011, 56, 59–66. [Google Scholar] [CrossRef]
- Breschi, L.; Maravic, T.; Mazzoni, A. Dentin bonding systems: From dentin collagen structure to bond preservation and clinical applications. Dent. Mater. 2018, 34, 78–96. [Google Scholar] [CrossRef]
- Breschi, L.; Mazzoni, A.; Dorigo, E.D.S. Dental adhesion review: Aging and stability of the bonded interface. Dent. Mater. 2008, 24, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Perdigão, J. Dentin bonding—Variables related to the clinical situation and the substrate treatment. Dent. Mater. 2010, 26, e24–e37. [Google Scholar] [CrossRef] [PubMed]
- Paken, G.; Çömlekoğlu, M.E.; Sonugelen, M. Detection of the hybrid layer biodegradation initiation factor with a scanning electron microscope. Microsc. Res. Tech. 2021, 84, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Stape, T.H.S.; Uctasli, M.; Tezvergil-Mutluay, A. Dry bonding to dentin: Broadening the moisture spectrum and increasing wettability of etch-and-rinse adhesives. Dent. Mater. 2021, 37, 1676–1687. [Google Scholar] [CrossRef] [PubMed]
- De Munck, J.; Van Landuyt, K.; Van Meerbeek, B. A critical review of the durability of adhesion to tooth tissue: Methods and results. J. Dent. Res. 2005, 84, 118–132. [Google Scholar] [CrossRef]
- Borges, M.A.P.; Matos, I.C.; Miranda, M.S. Degradation of polymeric restorative materials subjected to a high caries challenge. Dent. Mater. 2011, 27, 244–252. [Google Scholar] [CrossRef]
- Huang, B.; Siqueira, W.L.; Cvitkovitch, D.G.; Finer, Y. Esterase from a cariogenic bacterium hydrolyzes dental resins. Acta Biomater. 2018, 71, 330–338. [Google Scholar] [CrossRef]
- Huang, B.; Sadeghinejad, L.; Finer, Y. Gene expression and protein synthesis of esterase from Streptococcus mutans are affected by biodegradation by-product from methacrylate resin composites and adhesives. Acta Biomater. 2018, 81, 158–168. [Google Scholar] [CrossRef]
- Marashdeh, M.Q.; Gitalis, R.; Finer, Y. Enterococcus faecalis hydrolyzes dental resin composites and adhesives. J. Endod. 2018, 44, 609–613. [Google Scholar] [CrossRef]
- Huang, B.; Cvitkovitch, D.G.; Santerre, J.P.; Finer, Y. Biodegradation of resin–dentin interfaces is dependent on the restorative material, mode of adhesion, esterase or MMP inhibition. Dent. Mater. 2018, 34, 1253–1262. [Google Scholar] [CrossRef]
- Love, R.M.; Jenkinson, H.F. Invasion of dentinal tubules by oral bacteria. Crit. Rev. Oral. Biol. Med. 2002, 13, 171–183. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peled, Y.; Stewart, C.A.; Glogauer, M.; Finer, Y. The Role of Bacterial, Dentinal, Salivary, and Neutrophil Degradative Activity in Caries Pathogenesis. Dent. J. 2023, 11, 217. https://doi.org/10.3390/dj11090217
Peled Y, Stewart CA, Glogauer M, Finer Y. The Role of Bacterial, Dentinal, Salivary, and Neutrophil Degradative Activity in Caries Pathogenesis. Dentistry Journal. 2023; 11(9):217. https://doi.org/10.3390/dj11090217
Chicago/Turabian StylePeled, Yuval, Cameron A. Stewart, Michael Glogauer, and Yoav Finer. 2023. "The Role of Bacterial, Dentinal, Salivary, and Neutrophil Degradative Activity in Caries Pathogenesis" Dentistry Journal 11, no. 9: 217. https://doi.org/10.3390/dj11090217
APA StylePeled, Y., Stewart, C. A., Glogauer, M., & Finer, Y. (2023). The Role of Bacterial, Dentinal, Salivary, and Neutrophil Degradative Activity in Caries Pathogenesis. Dentistry Journal, 11(9), 217. https://doi.org/10.3390/dj11090217