Raman Fingerprints of Rice Nutritional Quality: A Comparison between Japanese Koshihikari and Internationally Renowned Cultivars

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Samples
2.2. Raman Spectroscopic Assessments
2.3. Statistical Analyses
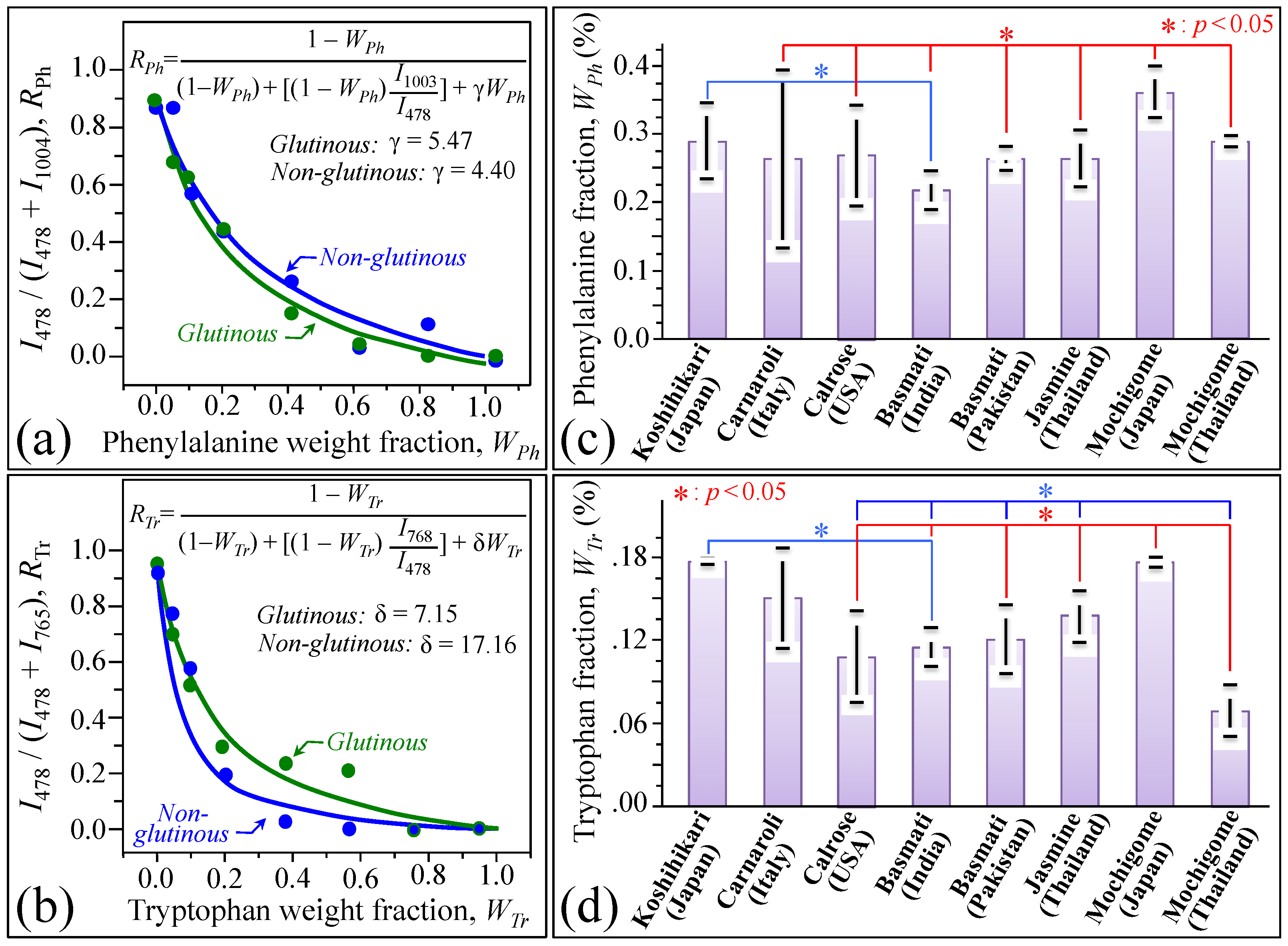
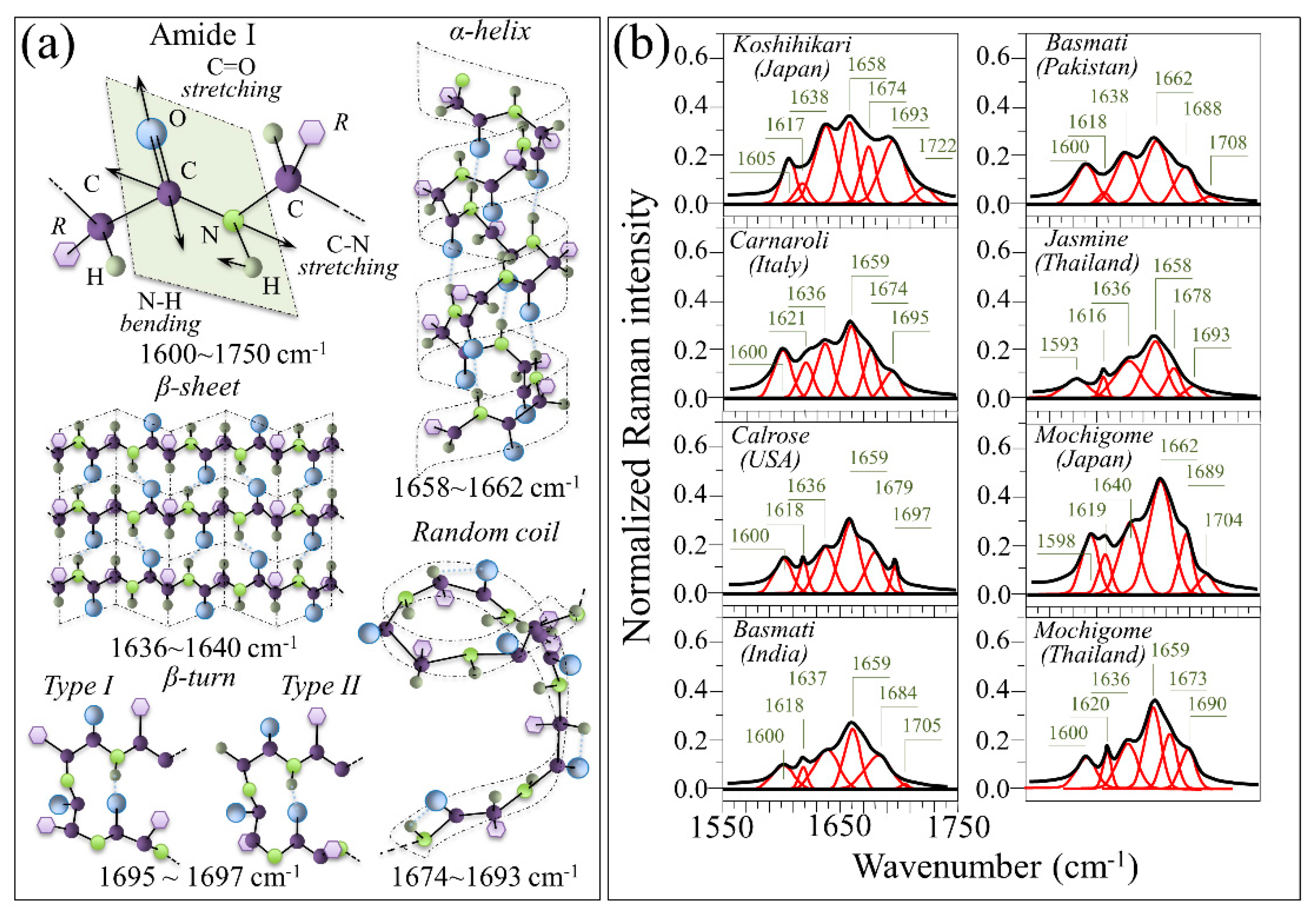
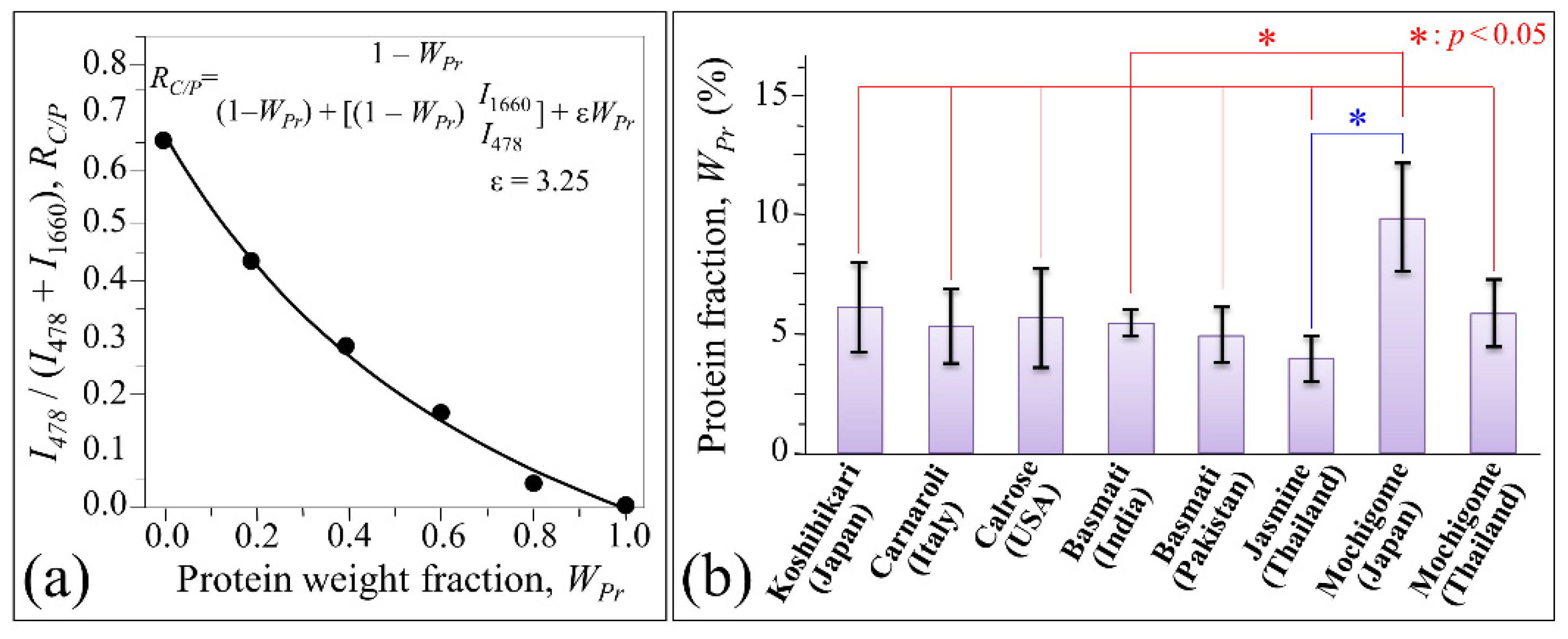
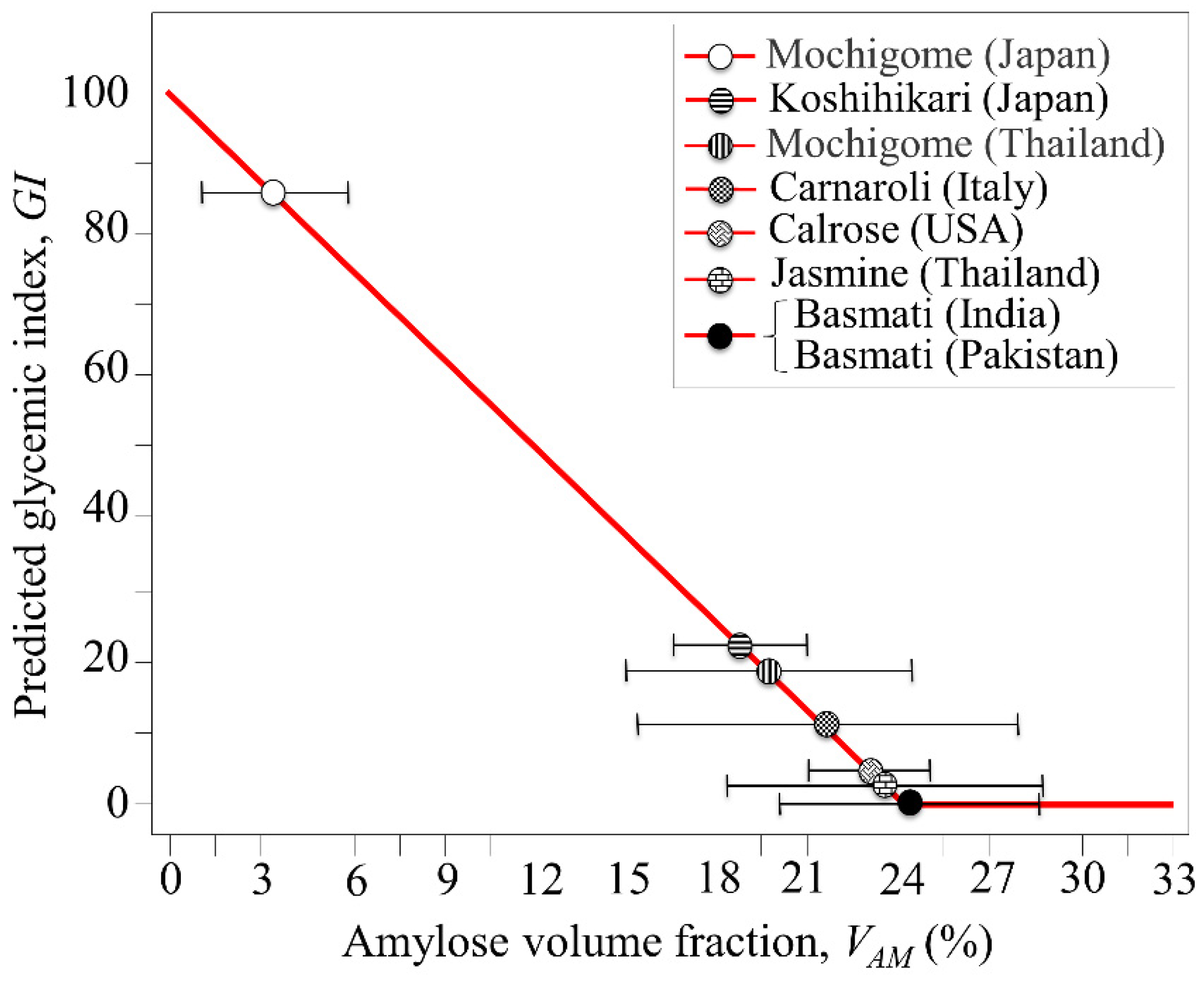
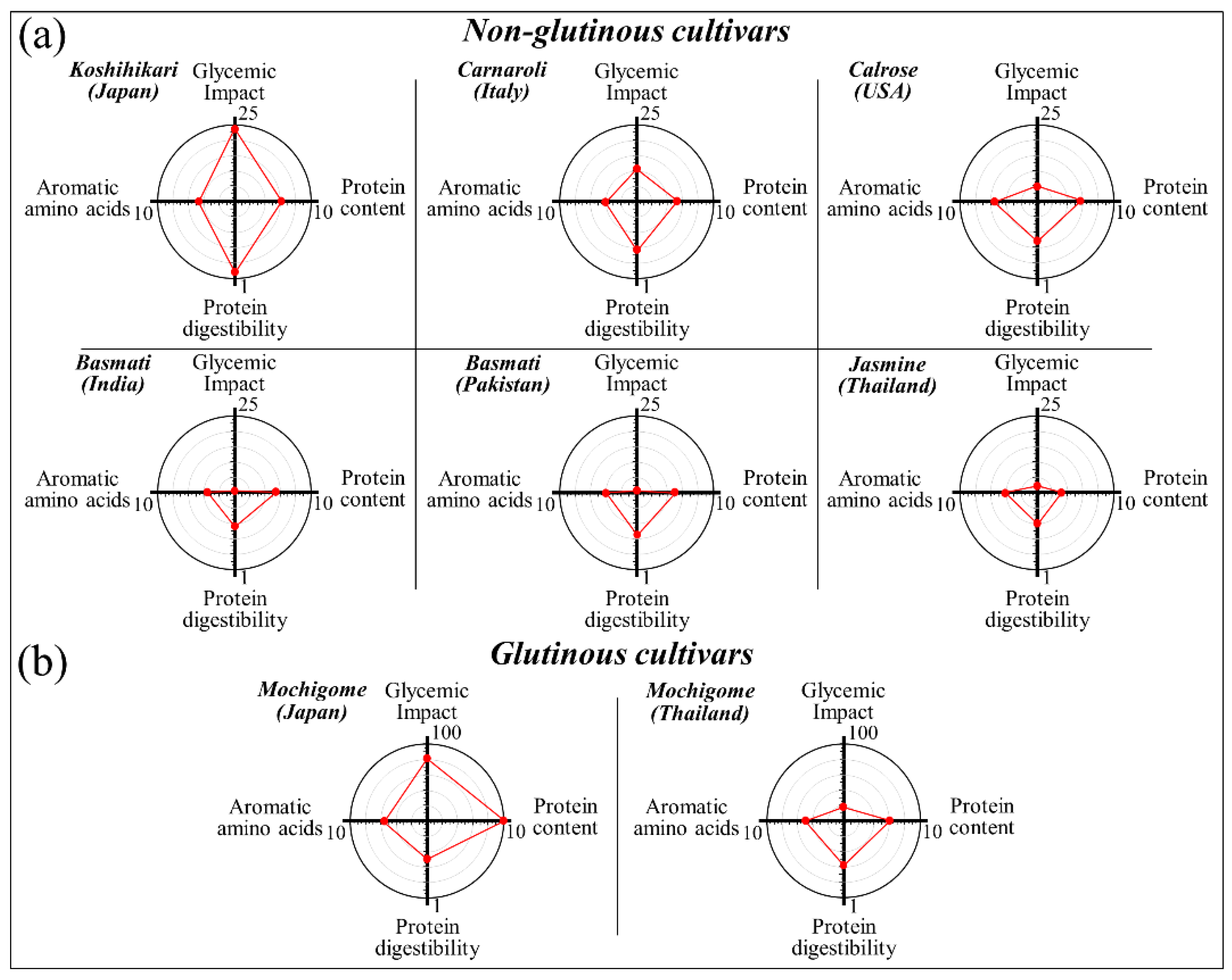
3. Results
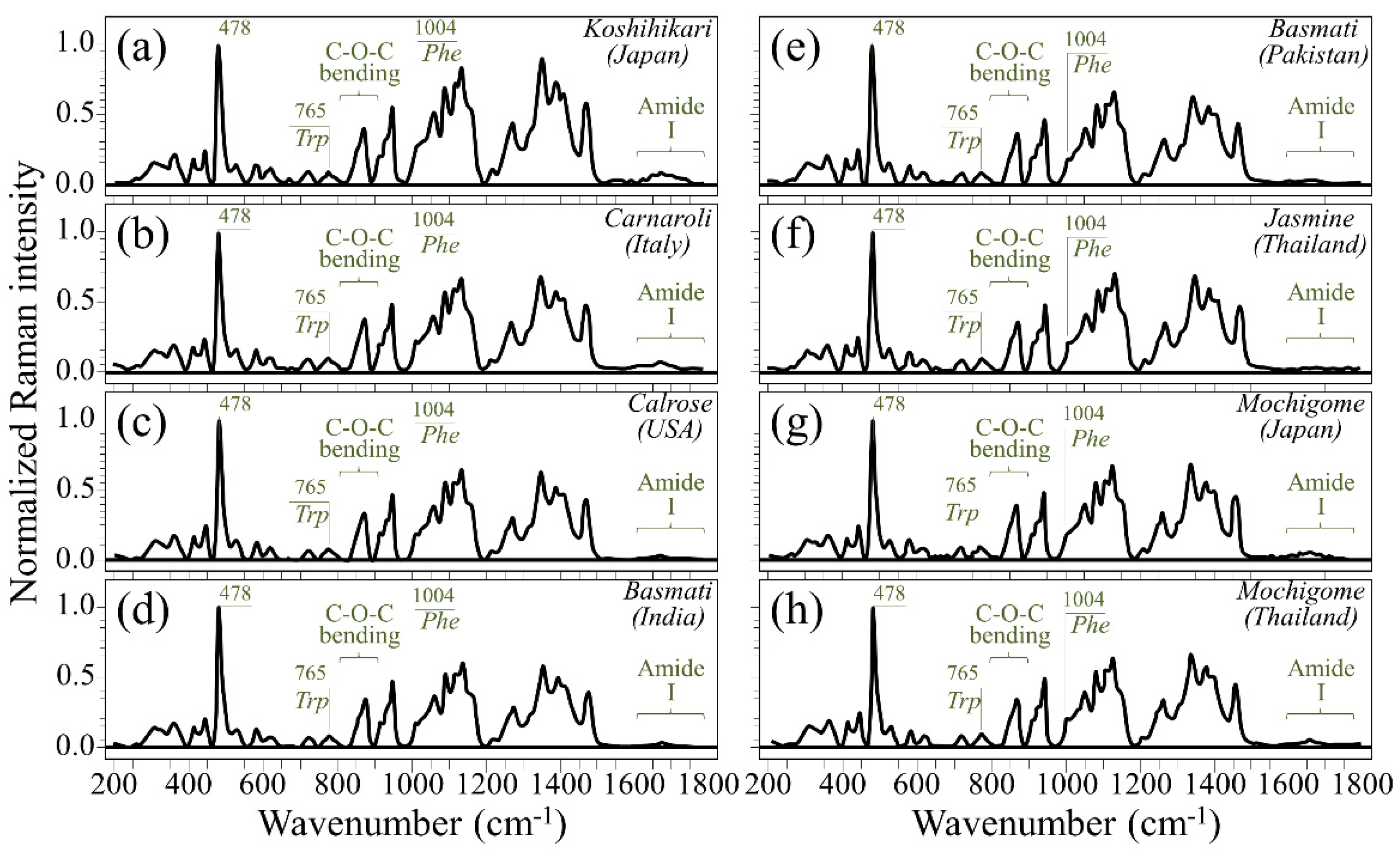
3.1. Raman Spectra of Koshihikari and Other Renowned Cultivars
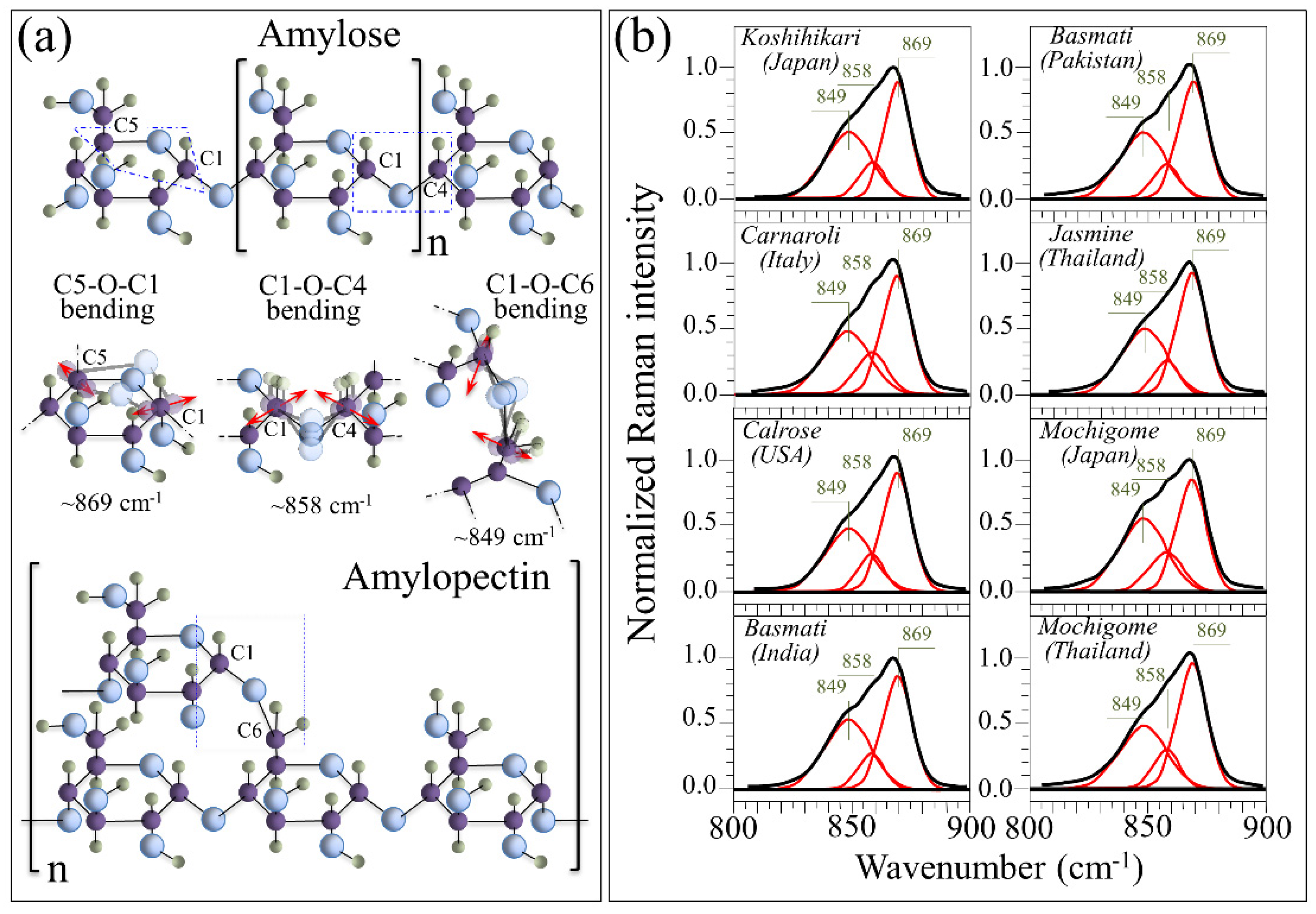
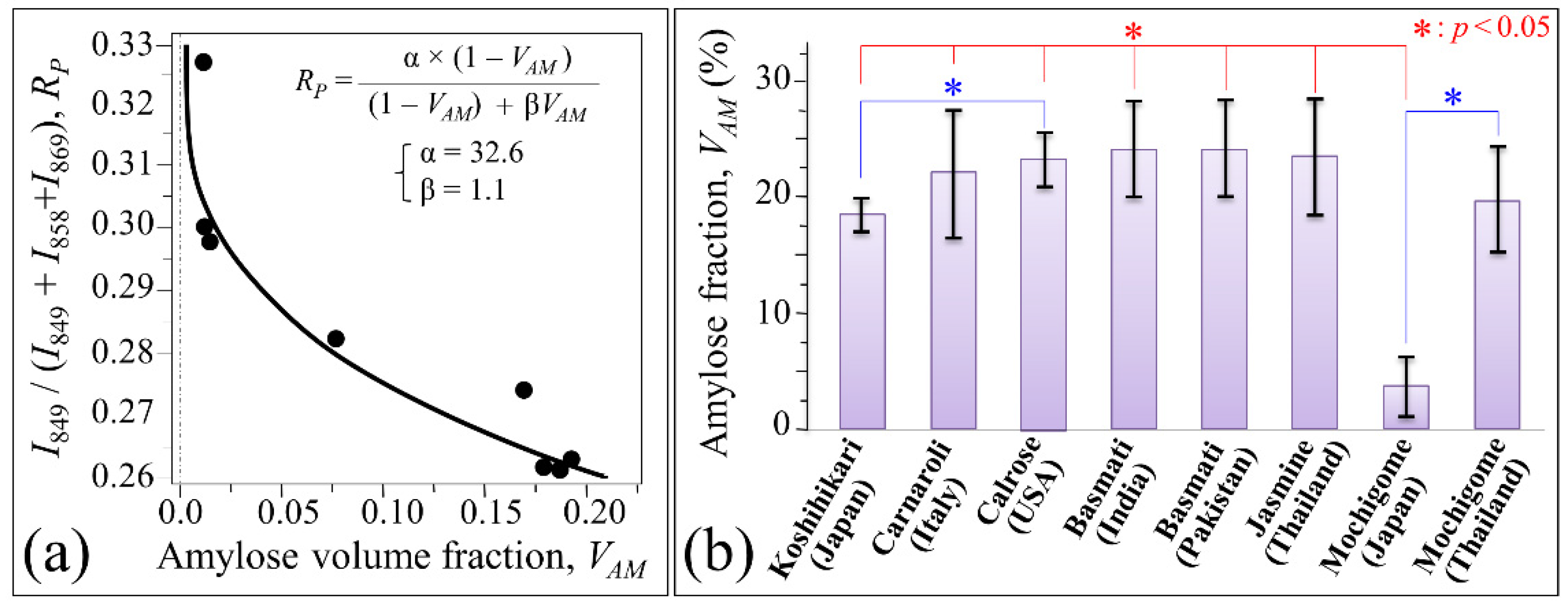
3.2. Quantitative Evaluation of Nutritional Quality of Rice Cultivars
4. Discussion
4.1. Glycemic Impact of Glutinous and Non-Glutinous Rice Cultivars
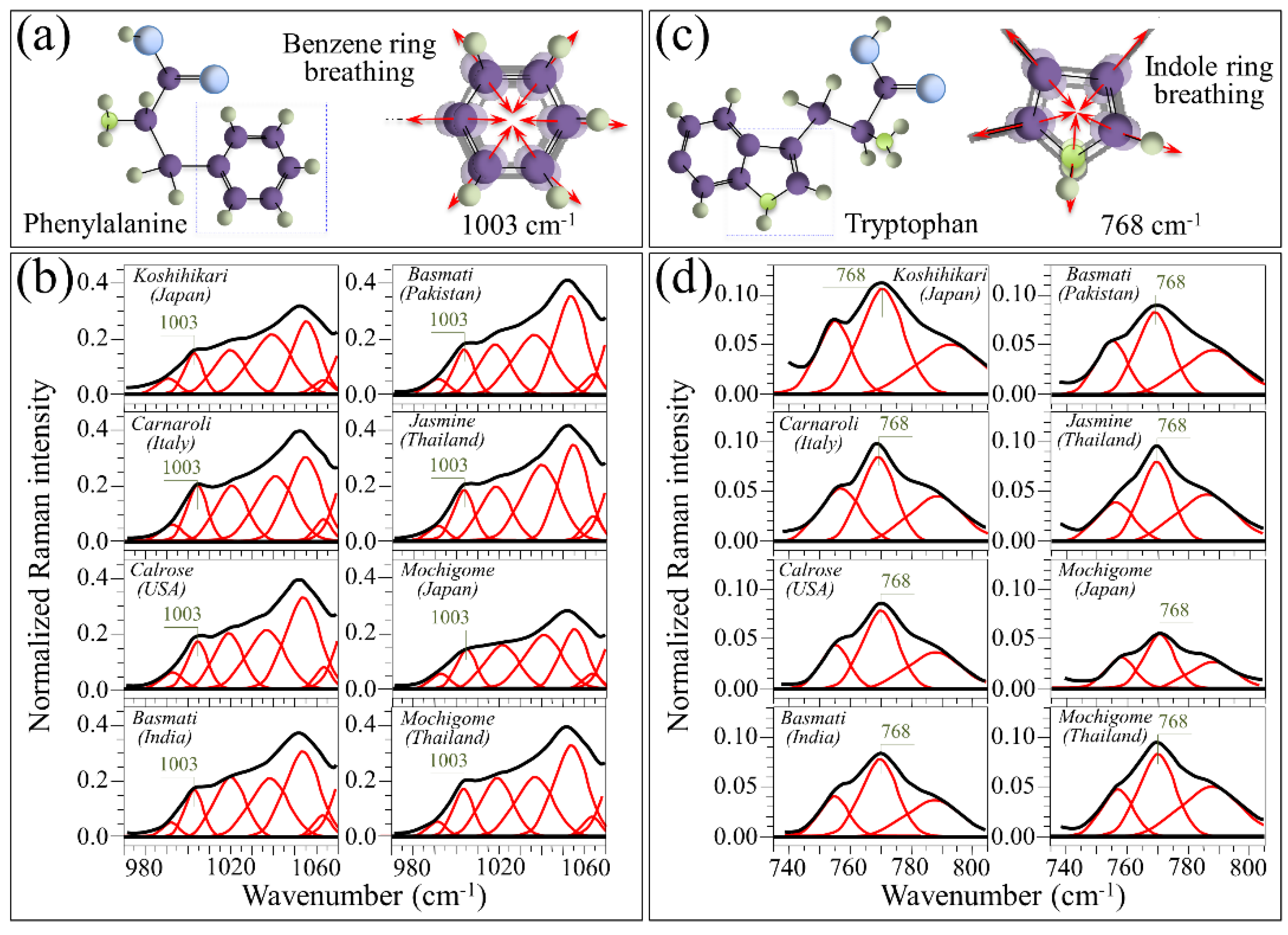
4.2. Aromatic Amino Acids Contents and Protein Content/Structure
4.3. Comparison of Nutritional Facts Retrieved from Raman Spectroscopy
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fitzgerald, M.; McCouch, S.; Hall, R. Not just a grain of rice: The quest for quality. Trends Plant Sci. 2019, 14, 133–139. [Google Scholar] [CrossRef]
- Mahender, A.; Anandan, A.; Pradhan, S.K.; Pandit, E. Rice grain nutritional traits and their enhancement using relevant genes and QTLs through advanced approaches. Springerplus 2016, 5, 2086. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Jiang, L.; Zheng, J.; Wang, T.; Wang, H.; Huang, Y.; Hong, Z. Genetic bases of rice grain shape: So many genes, so little known. Trends Plant Sci. 2013, 18, 218–226. [Google Scholar] [CrossRef]
- Li, X.; Wu, L.; Wang, J.; Sun, J.; Xia, X.; Geng, X.; Wang, X.; Xu, Z.; Xu, Q. Genome sequencing of rice subspecies and genetic analysis of recombinant lines reveals regional yield- and quality-associated loci. BMC Biol. 2018, 16, 102. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Sun, J.; Wu, G.; Wang, Y.; Zhang, H.; Wang, L.; Qian, H.; Qi, X. Identification of rice varieties and determination of their geographical origin in China using Raman spectroscopy. J. Cereal Sci. 2018, 82, 175–182. [Google Scholar] [CrossRef]
- Sha, M.; Zhang, D.; Zhang, Z.; Wei, J.; Chen, Y.; Wang, M.; Liu, J. Improving Raman spectroscopic identification of rice varieties by feature extraction. J. Raman Spectrosc. 2020, 51, 702–710. [Google Scholar] [CrossRef]
- Pezzotti, G.; Zhu, W.; Chikaguchi, H.; Marin, E.; Masumura, T.; Sato, Y.-i.; Nakazaki, T. Raman spectroscopic analysis of polysaccharides in popular Japanese rice cultivars. Food Chem. 2021, 354, 129434. [Google Scholar] [CrossRef]
- Pezzotti, G.; Zhu, W.; Chikaguchi, H.; Marin, E.; Boschetto, F.; Masumura, T.; Sato, Y.-I.; Nakazaki, T. Raman molecular fingerprints of rice nutritional quality and the concept of Raman barcode. Front. Nutr. 2021, 8, 663569. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Uehara, K.; Shim, R.; Shim, J.; Nagai, K.; Ashikari, M.; Takashi, T. Development of chromosome segment substitution lines harboring Oryza nivara genomic segments in Koshihikari and evaluation of yield- related traits. Breed. Sci. 2016, 66, 845–850. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef]
- Kobayashi, A.; Hori, K.; Yamamoto, T.; Yano, M. Koshihikari: A premium short-grain rice cultivar—Its expansion and breeding in Japan. Rice 2018, 11, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonelli, C.; Galassi, L.; Cormegna, M.; Bianchi, P. Chemical, physical, textural and sensory evaluation on Italian rice varieties. Univ. J. Agric. Res. 2017, 5, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.H. Registration of rice varieties. Agron. J. 1958, 50, 694–700. [Google Scholar] [CrossRef]
- Kishor, D.S.; Seo, J.; Chin, J.H.; Koh, H.-J. Evaluation of whole-genome sequence, genetic diversity, and agronomic traits of Basmati rice (Oryza sativa L.). Front. Genet. 2020, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Wongpornchai, S.; Sriseadka, T.; Choonvisase, S. Identification and quantitation of the rice aroma compound, 2-acetyl-1-pyrroline, in bread flowers (Vallaris glabra Ktze). J. Agric. Food Chem. 2003, 51, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Venn, B.J.; Kataoka, M.; Mann, J. The use of different reference foods in determining the glycemic index of starchy and non-starchy test foods. Nutr. J. 2014, 13, 50. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Abe, T.; Ando, H.; Sasahara, T. Classification of glutinous rice (Oryza sativa L.) starches based on X-ray diffraction pattern. Ann. Bot. 1993, 72, 551–557. [Google Scholar] [CrossRef]
- Pezzotti, G. Raman spectroscopy of piezoelectrics. J. Appl. Phys. 2013, 113, 211301. [Google Scholar] [CrossRef]
- Wiercigroch, E.; Szafraniec, E.; Czamara, K.; Pacia, M.Z.; Majzner, K.; Kochan, K.; Kaczor, A.; Baranska, M.; Malek, K. Raman and infrared spectroscopy of carbohydrates: A review. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 185, 317–335. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, Y.; Ren, H.; Zhao, Y.; Wang, Z.; Wu, F.; Xiao, Z. Structural changes in rice bran protein upon different extrusion temperatures: A Raman spectroscopic study. J. Chem. 2016, 2016, 6898715. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ma, C.-Y.; Yuen, S.N.; Phillips, D.L. Study of succinylated food proteins by Raman spectroscopy. J. Agric. Food Chem. 2004, 52, 1815–1823. [Google Scholar] [CrossRef]
- Alix, A.J.P.; Pedanou, G.; Berjot, M. Fast determination of the quantitative secondary structure of proteins by using some parameters of the Raman Amide I band. J. Mol. Struct. 1988, 174, 159–164. [Google Scholar] [CrossRef]
- Suwannaporn, P.; Linnemann, A. Rice-eating quality among consumers in different rice grain preference countries. J. Sens. Stud. 2008, 23, 1–13. [Google Scholar] [CrossRef]
- International-Organisation-for-Standardisation. ISO 6647:1987E. Rice: Determination of Amylose Content. 1987. Available online: http://iso.org/standard/13073.html (accessed on 29 September 2021).
- Zhu, G.; Zhu, X.; Fan, Q.; Wan, X. Raman spectra of amino acids and their aqueous solutions. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2011, 78, 1187–1195. [Google Scholar] [CrossRef]
- Takeuchi, H. UV Raman markers for structural analysis of aromatic side chains in proteins. Anal. Sci. 2011, 27, 1077–1086. [Google Scholar] [CrossRef] [Green Version]
- Phillips, D.L.; Xing, J.; Chong, C.K.; Liu, H.; Corke, H. Determination of the degree of succinylation in diverse modified starches by Raman spectroscopy. J. Agric. Food Chem. 2000, 48, 5105–5108. [Google Scholar] [CrossRef]
- Wolever, T.M.S.; Katzman-Relle, L.; Jenkins, A.L.; Vuksan, V.; Josse, R.G.; Jenkins, D.J.A. Glycaemic index of 102 complex carbohydrate foods in patients with diabetes. Nutr. Res. 1994, 4, 651–669. [Google Scholar] [CrossRef]
- Dona, A.C.; Guilhem, P.; Robert, G.G.; Philip, W.K. Digestion of starch: In vivo and in vitro kinetic models used to characterize oligosaccharide or glucose release. Carbohydr. Polym. 2010, 80, 599–617. [Google Scholar] [CrossRef]
- Ao, Z.; Simsek, S.; Zhang, G.; Venkatachalam, M.; Reuhs, B.L.; Hamaker, B.R. Starch with a slow digestion property produced by altering its chain length, branch density, and crystalline structure. J. Agric. Food Chem. 2007, 55, 4540–4547. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Y.; Hamaker, B.R. Slowly digestible starch: Concept, mechanism, and proposed extended glycemic index. Crit. Rev. Food Sci. 2009, 49, 852–867. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.-J.; Liu, Q.; Lee, L.; Wei, D. Relationship between the structure, physicochemical properties and in vitro digestibility of rice starches with different amylose contents. Food Hydrocoll. 2011, 25, 968–975. [Google Scholar] [CrossRef]
- Jeevetha, S.; Barakatun-Nisak, M.Y.; Ngan, H.-B.; Ismail, A.; Azlan, A. Relationship between amylose content and glycemic index of commonly consumed white rice. IOSR J. Agric. Vet. Sci. 2014, 7, 12–18. [Google Scholar] [CrossRef]
- Brand-Miller, J.; Pang, E.; Bramall, L. Rice: A high or low glycemic index food. Am. J. Clin. Nutr. 1992, 56, 1034–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NikShanita, S.; Hasnah, H.; Khoo, C.W. Amylose and amylopectin in selected Malaysian foods and its relationship to glycemic index. Sains Malays. 2011, 40, 865–870. [Google Scholar]
- Wolever, T.M.S.; Giddens, J.L.; Sievenpiper, J.L. Effect of ethnicity on glycaemic index: A systematic review and meta-analysis. Nutr. Diabetes 2015, 5, e170. [Google Scholar] [CrossRef]
- Hu, E.A.; Pan, A.; Malik, V.; Sun, Q. White rice consumption and risk of type 2 diabetes: Meta-analysis and systematic review. Br. Med. J. 2012, 344, e1454. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.B. Globalization of diabetes. Diabetes Care 2011, 34, 1249–1257. [Google Scholar] [CrossRef] [Green Version]
- Wakasa, K.; Ishihara, A. Metabolic engineering of the tryptophan and phenylalanine biosynthetic pathways in rice. Plant Biotechnol. 2009, 26, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Matsuda, F.; Kasai, K.; Fukuoka, S.; Kitamura, K.; Tozawa, Y.; Miyagawa, H.; Wakasa, K. Mutation of a rice gene encoding a phenylalanine biosynthetic enzyme results in accumulation of phenylalanine and tryptophan. Plant Cell 2008, 20, 1316–1329. [Google Scholar] [CrossRef] [Green Version]
- Tome, D. Criteria and markers for protein quality assessment—A review. Br. J. Nutr. 2012, 108, S222–S229. [Google Scholar] [CrossRef] [Green Version]
- Teng, B.; Zeng, R.Z.; Wang, Y.C.; Liu, Z.Q.; Zhang, Z.M.; Zhu, H.T.; Ding, X.H.; Li, W.T.; Zhang, G.Q. Detection of allelic variation at the Wx locus with single segment substitution lines in rice (Oryza sativa L.). Mol. Breed. 2012, 30, 583–595. [Google Scholar] [CrossRef]
- Nakamura, S.; Satoh, H.; Ohtsubo, K. Development of formulae for estimating amylose content, amylopectin chain length distribution, and resistant starch content based on the iodine absorption curve of rice starch. Biosci. Biotechnol. Biochem. 2015, 79, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Prakash, S.; Nicholson, T.M. Instrumental measurement of cooked rice texture by dynamic rheological testing and its relation to the fine structure of rice starch. Carbohydr. Polym. 2016, 146, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.L.; Sugiyama, J.; Shibata, M.; Kokawa, M.; Fujita, K.; Tsuta, M.; Nabetani, H.; Araki, T. Changes in the texture and viscoelastic properties of bread containing rice porridge during storage. Biosci. Biotechnol. Biochem. 2012, 76, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.U.; Suzuki, K.; Okadome, H.; Ikezaki, H.; Homma, S.; Ohtsubo, K. Detection of changes in taste of japonica and indica brown and milled rice (Oryza sativa L.) during storage using physicochemical analyses and a taste sensing system. J. Agric. Food Chem. 2005, 53, 1108–1118. [Google Scholar] [CrossRef]
- Ma, Z.-H.; Wang, Y.-B.; Cheng, H.-T.; Zhang, G.-C.; Lyu, W.-Y. Biochemical composition distribution in different grain layers is associated with the edible quality of rice cultivars. Food Chem. 2020, 311, 125896. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Anthony, L.D.; Tesfamichael, H.K.; Lloyd, W.S. Biomarker metabolites capturing the metabolite variance present in a rice plant developmental period. BMC Plant Biol. 2005, 5, 8. [Google Scholar]
- Koshihikari Brands Dominate Japanese Rice Ranking. Available online: https://www.nippon.com/en/japan-data/h00436/koshihikari-brands-dominate-japanese-rice-ranking.html (accessed on 23 September 2021).
- Rygula, A.; Majzner, K.; Marzec, K.M.; Kaczor, A.; Pilarczyka, M.; Baranska, M. Raman spectroscopy of proteins: A review. J. Raman Spectrosc. 2013, 44, 1061–1076. [Google Scholar] [CrossRef]
- Pelton, J.T.; McLean, L.R. Spectroscopic methods for analysis of protein secondary structure. Anal. Biochem. 2000, 277, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Tran, U.T.; Okadome, H.; Murata, M.; Homma, S.; Ohtsubo, K. Comparison of Vietnamese and Japanese rice cultivars in terms of physicochemical properties. Food Sci. Technol. Res. 2001, 7, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Mawal, Y.R.; Mawal, M.R.; Sainani, M.N.; Ranjekar, P.K. Rice seed storage proteins: A structural insight. Plant Sci. 1990, 70, 73–80. [Google Scholar] [CrossRef]
- Ogawa, M.; Kumamaru, T.; Satoh, H.; Iwata, N.; Omura, T.; Kasai, Z.; Tanaka, K. Purification of protein body-I of rice seed and its polypeptide composition. Plant Cell Physiol. 1987, 28, 1517–1527. [Google Scholar]
- Uehara, Y.; Kobayashi, A.; Ohta, H.; Shimizu, H.; Fukui, K.; Miura, K.; Otsuki, H.; Komaki, Y.; Sasahara, H. A new rice variety “Shunyou”. Bull. Natl. Agric. Res. Cent. 2002, 1, 1–21. (In Japanese) [Google Scholar]
- Yang, L.; Kumagai, T.; Kawamura, H.; Watanabe, T.; Kubota, M.; Fujimura, S.; Watanabe, R.; Kadowaki, M. Effects of rice proteins from two cultivars, Koshihikari and Shunyo, on cholesterol and triglyceride metabolism in growing and adult rats. Biosci. Biotechnol. Biochem. 2007, 71, 694–703. [Google Scholar] [CrossRef]
- Kodama, T.; Miyazaki, T.; Kitamura, I.; Suzuki, Y.; Namba, Y.; Sakurai, J.; Torikai, Y.; Inoue, S. Effects of single and long-term administration of wheat albumin on blood glucose control: Randomized controlled clinical trials. Eur. J. Clin. Nutr. 2005, 59, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Lankisch, M.; Layer, P.; Rizza, R.A.; DiMagno, E.P. Acute postprandial gastrointestinal and metabolic effects of wheat amylase inhibitor (WAI) in normal, obese, and diabetic humans. Pancreas 1998, 17, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. Rice: Chemistry and Technology, 2nd ed.; American Association of Cereal Chemists (AACC): St. Paul, MN, USA, 1985; p. 774. [Google Scholar]
- Ina, S.; Ninomiya, K.; Mogi, T.; Hase, A.; Ando, T.; Matsukaze, N.; Ogihara, J.; Akao, M.; Kumagai, H.; Kumagai, H. Rice (Oryza sativa japonica) albumin suppresses the elevation of blood glucose and plasma insulin levels after oral glucose loading. J. Agric. Food Chem. 2016, 64, 4882–4890. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.; Nguyen, S.D.; Kim, M.R.; Sok, D.-E. Rice albumin N-terminal (Asp-His-His-Gln) prevents against copper ion-catalyzed oxidations. J. Agric. Food Chem. 2007, 55, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Ofosu, F.K.; Fafa Mensah, D.-J.; Banan-Mwine Daliri, E.; Oh, D.-H. Exploring molecular insights of cereal peptidic antioxidants in metabolic syndrome prevention. Antioxidants 2021, 10, 518. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pezzotti, G.; Zhu, W.; Hashimoto, Y.; Marin, E.; Masumura, T.; Sato, Y.-I.; Nakazaki, T. Raman Fingerprints of Rice Nutritional Quality: A Comparison between Japanese Koshihikari and Internationally Renowned Cultivars. Foods 2021, 10, 2936. https://doi.org/10.3390/foods10122936
Pezzotti G, Zhu W, Hashimoto Y, Marin E, Masumura T, Sato Y-I, Nakazaki T. Raman Fingerprints of Rice Nutritional Quality: A Comparison between Japanese Koshihikari and Internationally Renowned Cultivars. Foods. 2021; 10(12):2936. https://doi.org/10.3390/foods10122936
Chicago/Turabian StylePezzotti, Giuseppe, Wenliang Zhu, Yuuki Hashimoto, Elia Marin, Takehiro Masumura, Yo-Ichiro Sato, and Tetsuya Nakazaki. 2021. "Raman Fingerprints of Rice Nutritional Quality: A Comparison between Japanese Koshihikari and Internationally Renowned Cultivars" Foods 10, no. 12: 2936. https://doi.org/10.3390/foods10122936
APA StylePezzotti, G., Zhu, W., Hashimoto, Y., Marin, E., Masumura, T., Sato, Y. -I., & Nakazaki, T. (2021). Raman Fingerprints of Rice Nutritional Quality: A Comparison between Japanese Koshihikari and Internationally Renowned Cultivars. Foods, 10(12), 2936. https://doi.org/10.3390/foods10122936