
Sprouting of Sorghum (Sorghum bicolor [L.] Moench): Effect of Drying Treatment on Protein and Starch Features

 ,
,  ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Proximate Composition
2.3. Protein Characterization and Functionality
2.3.1. Protein Extraction and Fractionation
2.3.2. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.4. Amino Acid Analysis
2.5. Water-Holding Capacity (WHC) and Oil-Holding Capacity (OHC)
2.6. Starch Characterization and Functionality
2.6.1. Total Starch (TS), Resistant Starch (RS), Digestible Starch (DS), Amylose Content
2.6.2. α-Amylase Activity, Pasting and Thermal Properties and Swelling Power (SP)
2.7. Microstructure
2.8. Statistical Analysis
3. Results and Discussions
3.1. Proximate Composition
3.2. Protein Characterization and Functionality
3.2.1. Protein Extraction and Fractionation
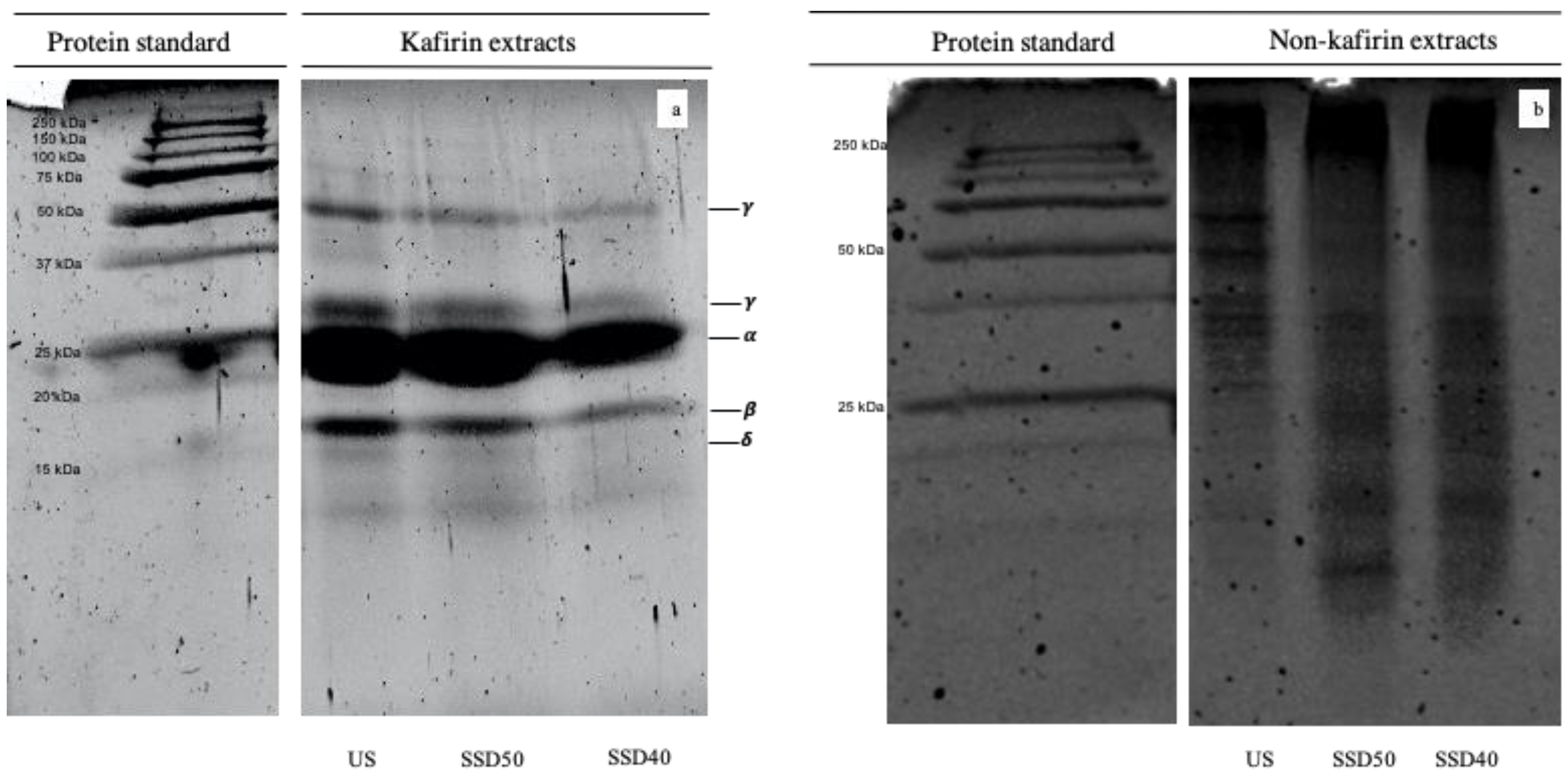
3.2.2. SDS-PAGE
3.3. Amino Acid Analysis
3.4. Water-Holding Capacity (WHC) and Oil-Holding Capacity (OHC)
3.5. Starch Characterization and Functionality
3.5.1. Total Starch (TS), Resistant Starch (RS), Digestible Starch (DS), Amylose Content
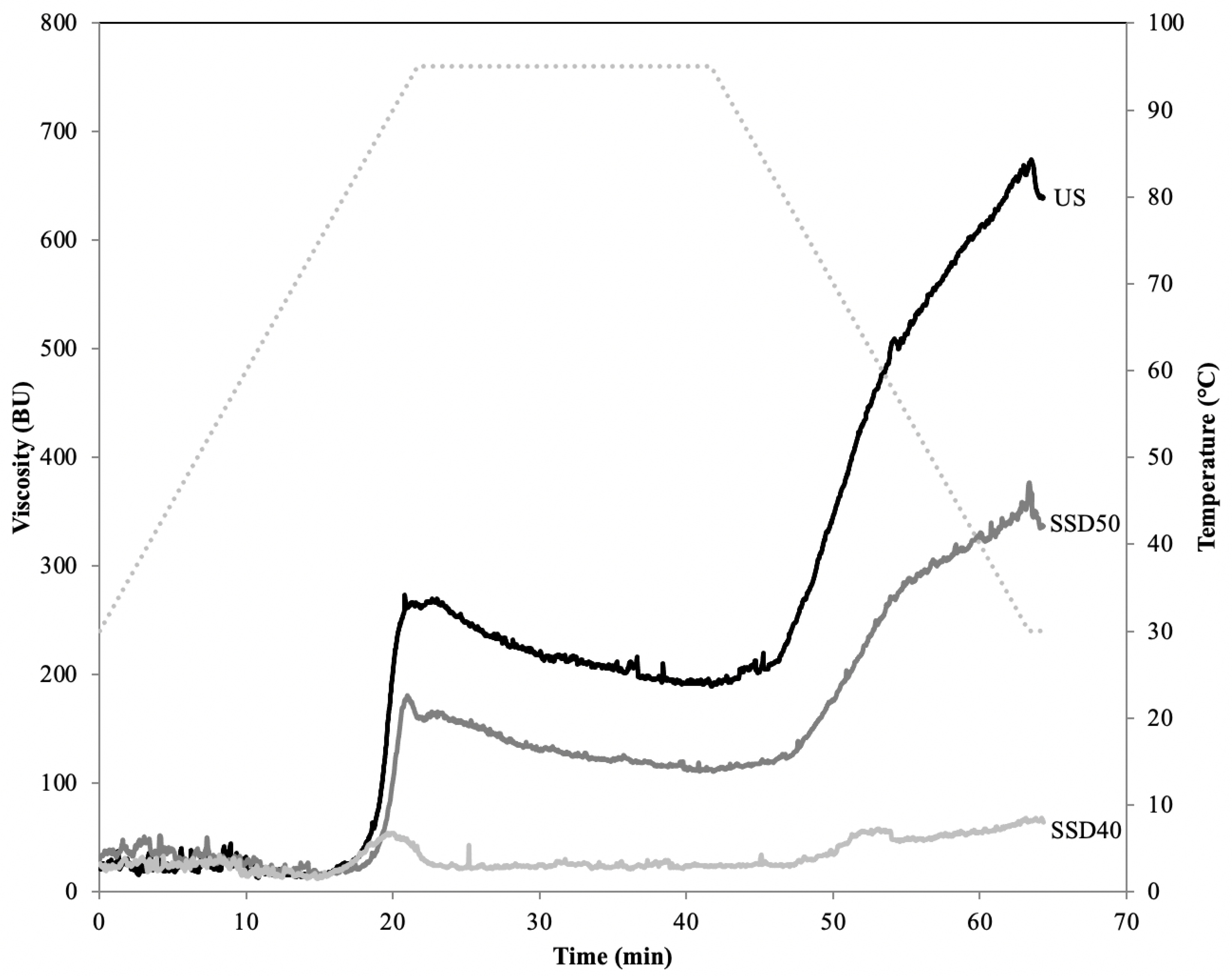
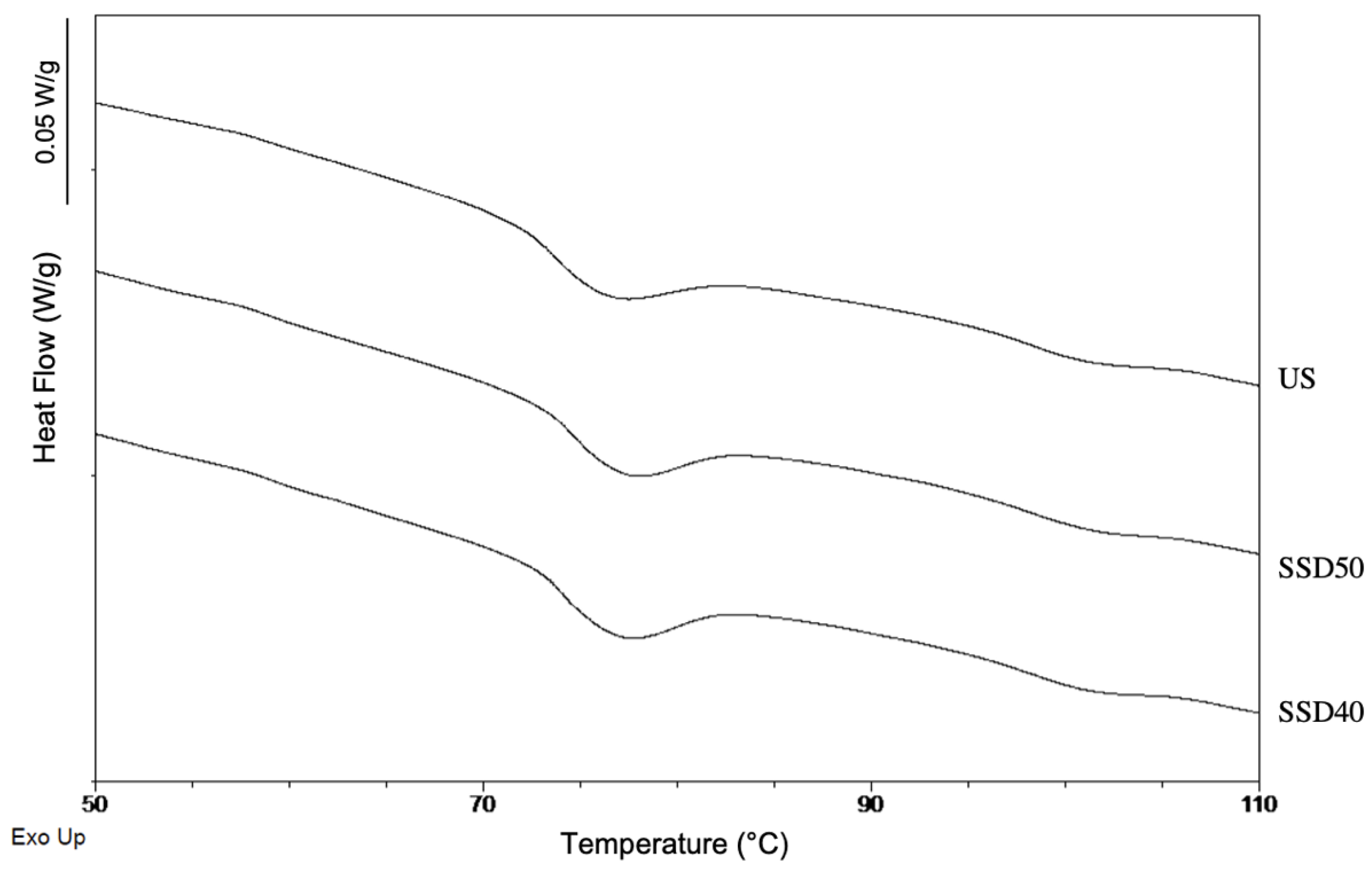
3.5.2. α-Amylase Activity, Pasting and Thermal Properties, Swelling Power (SP)
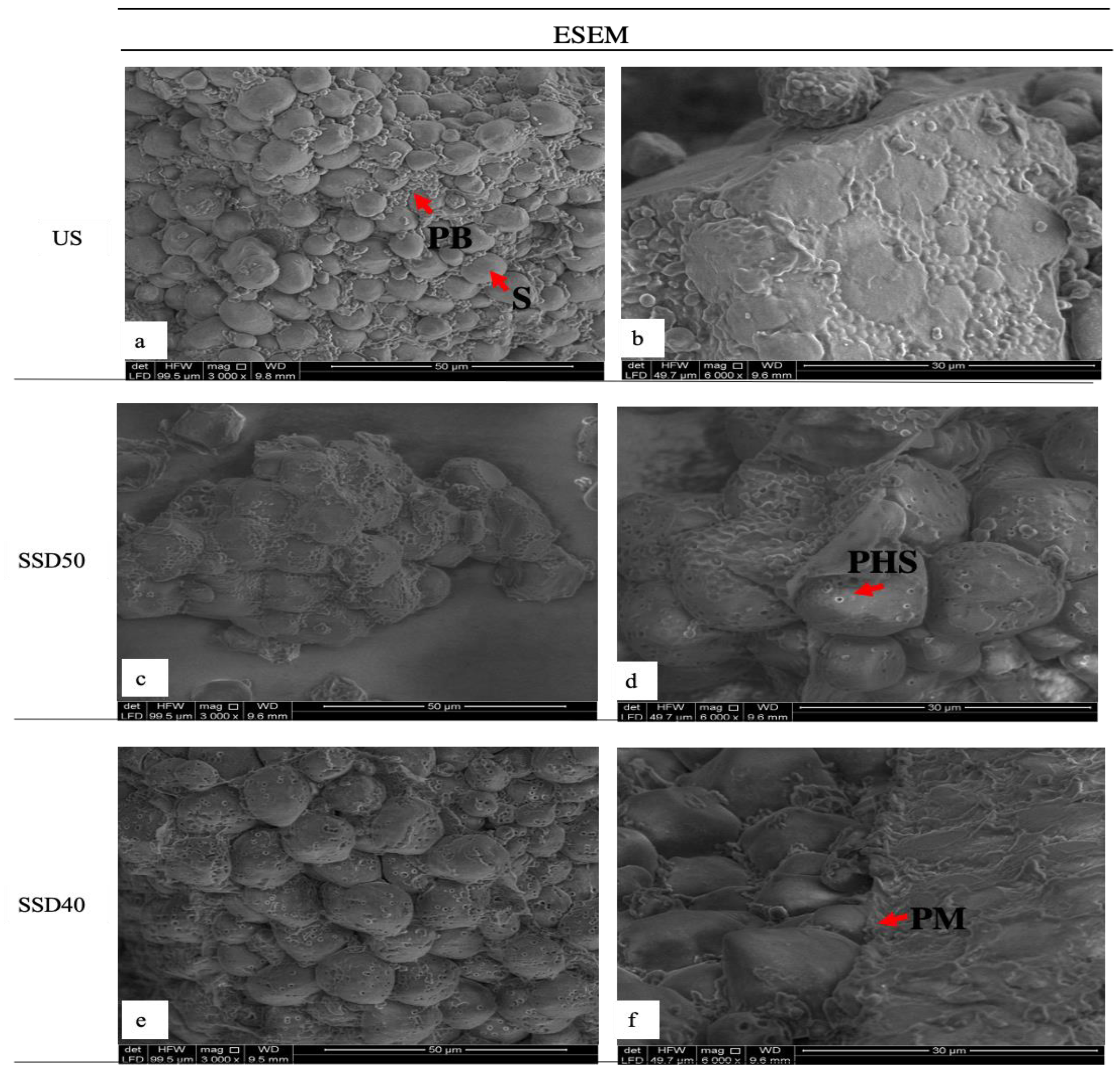
3.6. Microstructure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| US | Unsprouted sorghum flour |
| SSD50 | Flour from sprouted sorghum dried at 50 °C for 6 h |
| SSD40 | Flour from sprouted sorghum dried at 40 °C for 12 h |
| RH | Relative humidity |
| TDF | Total Dietary Fiber |
| IDF | Insoluble Dietary Fiber |
| SDF | Soluble Dietary Fiber |
| RS | Resistant starch |
| DS | Digestible Starch |
| TS | Total Starch |
| BSA | Bovine serum albumin |
| ΔH | Enthalpy |
| Ton | Onset temperature |
| Tp | Peak temperature |
| Toff | Offset temperature |
| Sp | Swelling power |
| R/T | Room temperature |
| SDS | Sodium dodecyl sulfate |
| β-ME | β-mercaptoethanol |
| DH | Degree of hydrolysis |
| UPLC | Ultra-performance liquid chromatography |
| ESI-MS | Electrospray ionization mass spectroscopy |
| SIR | Single Ion Recording |
| WHC | Water-holding capacity |
| OHC | Oil-holding capacity |
| BU | Brabender Units |
| LOQ | Limit of quantification |
| ESEM | Environmental scanning electron microscopy |
| DSC | Differential scanning calorimetry |
| SDS-PAGE | Sodium dodecyl sulphate-polyacrylamide gel electrophoresis |
| HMW | High molecular weight |
| Mr | Molecular weight |
| ANOVA | Analysis of variance |
| AA | Amino acids |
| EA | Essential amino acids |
| NEA | Non-essential amino acids |
| TA | Total amino acids |
References
- Hariprasanna, K.; Rakshit, S. Economic Importance of Sorghum. In The Sorghum Genome. Compendium of Plant Genomes; Rakshit, S., Wang, Y.H., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–25. [Google Scholar] [CrossRef]
- De Mesa-Stonestreet, N.J.; Alavi, S.; Bean, S.R. Sorghum proteins: The concentration, isolation, modification, and food applications of kafirins. J. Food Sci. 2010, 75, 5. [Google Scholar] [CrossRef] [Green Version]
- Kulamarva, A.G.; Sosle, V.R.; Raghavan, G.V. Nutritional and rheological properties of sorghum. Int. J. Food Prop. 2009, 12, 55–69. [Google Scholar] [CrossRef]
- Zhu, F. Structure, physicochemical properties, modifications, and uses of sorghum starch. Compr. Rev. Food Sci. Food Saf. 2014, 13, 597–610. [Google Scholar] [CrossRef]
- Zhang, G.; Hamaker, B.R. Sorghum (Sorghum bicolor L. Moench) flour pasting properties influenced by free fatty acids and protein. Cereal Chem. 2005, 82, 534–540. [Google Scholar] [CrossRef]
- Anyango, J.O.; Taylor, J.R. Sorghum Flour and Flour Products: Production, Nutritional Quality, and Fortification. In Flour and Breads and Their Fortification in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 137–151. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, S.; Singh, B. Effect of germination time and temperature on the functionality and protein solubility of sorghum flour. J. Cereal Sci. 2017, 76, 131–139. [Google Scholar] [CrossRef]
- Lemmens, E.; Moroni, A.V.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.A.; Van den Broeck, H.C.; Brouns, F.J.; De Brier, N.; et al. Impact of cereal seed sprouting on its nutritional and technological properties: A critical review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 305–328. [Google Scholar] [CrossRef] [Green Version]
- Correia, I.; Nunes, A.; Barros, A.S.; Delgadillo, I. Protein profile and malt activity during sorghum germination. J. Sci. Food Agric. 2008, 88, 2598–2605. [Google Scholar] [CrossRef]
- Afify, A.E.M.M.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Protein solubility, digestibility and fractionation after germination of sorghum varieties. PLoS ONE 2012, 7, e31154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkhalifa, A.E.O.; Bernhardt, R. Influence of grain germination on functional properties of sorghum flour. Food Chem. 2010, 121, 387–392. [Google Scholar] [CrossRef]
- Singh, A.K.; Rehal, J.; Kaur, A.; Jyot, G. Enhancement of attributes of cereals by germination and fermentation: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1575–1589. [Google Scholar] [CrossRef]
- Marengo, M.; Bonomi, F.; Marti, A.; Pagani, M.A.; Abd Elmoneim, O.E.; Iametti, S. Molecular features of fermented and sprouted sorghum flours relate to their suitability as components of enriched gluten-free pasta. LWT 2015, 63, 511–518. [Google Scholar] [CrossRef]
- Muralikrishna, G.; Nirmala, M. Cereal α-amylases—An overview. Carbohydr. Polym. 2005, 60, 163–173. [Google Scholar] [CrossRef]
- Phiarais, B.P.N.; Wijngaard, H.H.; Arendt, E.K. The impact of kilning on enzymatic activity of buckwheat malt. J. Inst. Brew. 2005, 111, 290–298. [Google Scholar] [CrossRef]
- Nkama, I.; Gbenyi, D.I.; Hamaker, B.R. Effects of malting and roasting of millet and sorghum on protein digestibility, mineral availability, soluble sugar composition and consumer acceptability of Dakuwa. Indian J. Nutr. 2015, 2, 1–6. [Google Scholar]
- Phattanakulkaewmorie, N.; Paseephol, T.; Moongngarm, A. Chemical compositions and physico-chemical properties of malted sorghum flour and characteristics of gluten free bread. World Acad. Sci. Eng. Technol. 2011, 5, 532–538. [Google Scholar] [CrossRef]
- Afify, A.E.M.M.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Bioavailability of iron, zinc, phytate and phytase activity during soaking and germination of white sorghum varieties. PLoS ONE 2011, 6, e25512. [Google Scholar] [CrossRef] [Green Version]
- Marti, A.; Cardone, G.; Nicolodi, A.; Quaglia, L.; Pagani, M.A. Sprouted wheat as an alternative to conventional flour improvers in bread-making. LWT 2017, 80, 230–236. [Google Scholar] [CrossRef]
- AACC. Approved Methods of Analysis, 11th ed.; Cereals & Grains Association: St. Paul, MN, USA, 2001. [Google Scholar]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Analytical: Washington, DC, USA, 2003; Volume I. [Google Scholar]
- Hamaker, B.R.; Mohamed, A.A.; Habben, J.E.; Huang, C.P.; Larkins, B.A. Efficient procedure for extracting maize and sorghum kernel proteins reveal higher prolamin contents than the conventional method. Cereal Chem. 1995, 72, 583–588. [Google Scholar]
- Park, S.H.; Bean, S.R. Investigation and Optimization of the Factors Influencing Sorghum Protein Extraction. J. Agric. Food Chem. 2003, 51, 7050–7054. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Anzani, C.; Prandi, B.; Tedeschi, T.; Baldinelli, C.; Sorlini, G.; Wierenga, P.A.; Dossena, A.; Sforza, S. Degradation of Collagen Increases Nitrogen Solubilisation During Enzymatic Hydrolysis of Fleshing Meat. Waste Biomass Valorization 2018, 9, 1113–1119. [Google Scholar] [CrossRef]
- Buhler, S.; Solari, F.; Gasparini, A.; Montanari, R.; Sforza, S.; Tedeschi, T. UV irradiation as a comparable method to thermal treatment for producing high quality stabilized milk whey. LWT 2019, 105, 127–134. [Google Scholar] [CrossRef]
- Marchini, M.; Carini, E.; Cataldi, N.; Boukid, F.; Blandino, M.; Ganino, T.; Vittadini, E.; Pellegrini, N. The use of red lentil flour in bakery products: How do particle size and substitution level affect rheological properties of wheat bread dough? LWT 2020, 110299. [Google Scholar] [CrossRef]
- Ekstrand, B.; Gangby, I.; Akesson, G. Lipase activity in oats–distribution, pH dependence and heat inactivation. Cereal Chem. 1992, 69, 379–381. [Google Scholar]
- El Nour, I.N.A.; Peruffo, A.D.; Curioni, A. Characterization of sorghum kafirins in relation to their cross-linking behaviour. J. Cereal Sci. 1998, 28, 197–207. [Google Scholar] [CrossRef]
- Belton, P.S.; Delgadillo, I.; Halford, N.G.; Shewry, P.R. Kafirin structure and functionality. J. Cereal Sci. 2006, 44, 272–286. [Google Scholar] [CrossRef] [Green Version]
- Georget, D.M.; Abd Elmoneim, O.E.; Peter, S.B. Structural changes in kafirin extracted from a white type II tannin sorghum during germination. J. Cereal Sci. 2012, 55, 106–111. [Google Scholar] [CrossRef]
- Mokhawa, G.; Kerapeletswe-Kruger, C.K.; Ezeogu, L.I. Electrophoretic analysis of malting degradability of major sorghum reserve proteins. J. Cereal Sci. 2013, 58, 191–199. [Google Scholar] [CrossRef]
- Mokrane, H.; Amoura, H.; Belhaneche-Bensemra, N.; Courtin, C.M.; Delcour, J.A.; Nadjemi, B. Assessment of Algerian sorghum protein quality [Sorghum bicolor (L.) Moench] using amino acid analysis and in vitro pepsin digestibility. Food Chem. 2010, 121, 719–723. [Google Scholar] [CrossRef]
- Gan, R.Y.; Lui, W.Y.; Wu, K.; Chan, C.L.; Dai, S.H.; Sui, Z.Q.; Corke, H. Bioactive compounds and bioactivities of germinated edible seeds and sprouts: An updated review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Mahajan, H.; Gupta, M. Nutritional, functional and rheological properties of processed sorghum and ragi grains. Cogent Food Agric. 2015, 1, 1109495. [Google Scholar] [CrossRef]
- De Carvalho Teixeira, N.; Queiroz, V.A.; Rocha, M.C.; Amorim, A.C.; Soares, T.O.; Monteiro, M.A.; de Menezes, C.B.; Schaffert, R.E.; Garcia, M.A.; Junqueira, R.G. Resistant starch content among several sorghum (Sorghum bicolor) genotypes and the effect of heat treatment on resistant starch retention in two genotypes. Food Chem. 2016, 197, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Gamel, T.H.; Linssen, J.P.; Mesallem, A.S.; Damir, A.A.; Shekib, L.A. Effect of seed treatments on the chemical composition and properties of two amaranth species: Starch and protein. J. Sci. Food Agric. 2005, 85, 319–327. [Google Scholar] [CrossRef]
- Mariotti, M.; Zardi, M.; Lucisano, M.; Pagani, M.A. Influence of the heating rate on the pasting properties of various flours. Starch-Stärke 2005, 57, 564–572. [Google Scholar] [CrossRef]
- Sun, Q.; Han, Z.; Wang, L.; Xiong, L. Physicochemical differences between sorghum starch and sorghum flour modified by heat-moisture treatment. Food Chem. 2014, 145, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Yeh, A.I. Relationships between thermal, rheological characteristics and swelling power for various starches. J. Food Eng. 2001, 50, 141–148. [Google Scholar] [CrossRef]
- Palmer, G.H. Enzymic degradation of the endosperm cell walls of germinated sorghum. World J. Microbiol. Biotechnol. 1991, 7, 17–21. [Google Scholar] [CrossRef]
- Wu, H.C.; Bulgakov, V.P.; Jinn, T.L. Pectin methylesterases: Cell wall remodeling proteins are required for plant response to heat stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| US | SSD50 | SSD40 | |
|---|---|---|---|
| Protein (g/100 g d.b.) | 11.27 ± 0.31b | 10.98 ± 0.48b | 11.97 ± 0.00a |
| Fat (g/100 g d.b.) | 3.71 ± 0.03a | 3.25 ± 0.00b | 3.07 ± 0.02c |
| Moisture (g/100 g w.b.) | 11.31 ± 0.15b | 12.69 ± 0.04a | 12.67 ± 0.03a |
| Ash (g/100 g d.b.) | 1.35 ± 0.02a | 1.30 ± 0.00c | 1.33 ± 0.00b |
| Carbohydrates (g/100 g d.b.) | 85.25 ± 0.12a | 84.48 ± 0.53b | 83.63 ± 0.07c |
| Total Dietary Fiber (TDF, g/100 g d.b.) | 7.4 ± 0.67 | 7.1 ± 0.5 | 7.4 ± 0.7 |
| Insoluble (IDF, g/100 g d.b.) | 7.3 ± 0.8 (98.65) | 7.1 ± 0.5 (98.59) | 7.1 ± 0.85 (95.95) |
| Soluble (SDF, g/100 g d.b.) | <LOQ | <LOQ | <LOQ |
| WHC (g/g) | 1.51 ± 0.05b | 1.52 ± 0.08b | 1.68 ± 0.05a |
| OHC (g/g) | 0.95 ± 0.01b | 1.00 ± 0.07ab | 1.05 ± 0.00a |
| Total Starch (TS, g/100 g d.b.) | 73.7 ± 0.0a | 69.9 ± 0.1b | 69.1 ± 0.2c |
| Digestible (DS, g/100 g d.b.) | 66.9 ± 0.0a (90.8) | 65.3 ± 0.0b (93.5) | 64.8 ± 0.3c (93.7) |
| Resistant (RS, g/100 g d.b.) | 6.8 ± 0.0a (9.2) | 4.6 ± 0.1b (6.6) | 4.3 ± 0.2c (6.3) |
| Amylose (%) | 29 ± 3b | 34 ± 4b | 47 ± 0a |
| Amylose/Amylopectin ratio | 0.41 | 0.51 | 0.88 |
| α-amylase activity (CU/g) | 0.06 ± 0.01b | 22.60 ± 0.85a | 20.64 ± 0.88a |
| Pasting properties (MVAG Test) | |||
| Pasting temperature (°C) | 78.7 ± 0.3b | 80.9 ± 1.1a | 76.5 ± 0.1c |
| Peak viscosity (BU) | 269.5 ± 3.5a | 187.0 ± 6.0b | 92.5 ± 5.5c |
| Peak temperature (°C) | 93.7 ± 1.3a | 92.2 ± 0.8ab | 89.9 ± 1.9b |
| Final viscosity (BU) | 643.5 ± 4.5a | 335.5 ± 1.5b | 105.0 ± 11.0c |
| Breakdown (BU) | 80.0 ± 2.0a | 74.5 ± 6.5b | 47.5 ± 6.5c |
| Setback (BU) | 482.5 ± 5.5a | 247.50± 15.5b | 59.0 ± 17.0c |
| SP (g/g) | |||
| 60 °C | 5.33 ± 0.33aB | 5.39 ± 0.24aC | 4.08 ± 0.43bC |
| 70 °C | 5.32 ± 0.32B | 5.35 ± 0.23C | 5.01 ± 0.33B |
| 80 °C | 8.36 ± 0.22aA | 6.86 ± 0.24bB | 5.57 ± 0.07cA |
| 90 °C | 8.23 ± 0.15aA | 7.63 ± 0.13bA | 6.08 ± 0.14cA |
| Amino Acid (g/100 g Flour) | US | SSD50 | SSD40 |
|---|---|---|---|
| Essential amino acids (EA) | |||
| Histidine | 0.193 ± 0.011 | 0.210 ± 0.005 | 0.204 ± 0.005 |
| Isoleucine | 0.372 ± 0.005 b | 0.386 ± 0.007 ab (+3.76) | 0.391 ± 0.010 a (+5.11) |
| Leucine | 1.256 ± 0.021 | 1.232 ± 0.027 | 1.276 ± 0.029 |
| Lysine | 0.147 ± 0.002 c | 0.153 ± 0 b (+4.08) | 0.160 ± 0.001 a (+8.84) |
| Methionine | 0.169 ± 0.011 b | 0.174 ± 0.003 b (+2.96) | 0.189 ± 0 a (+11.83) |
| Phenylalanine | 0.550 ± 0.007 | 0.544 ± 0.005 | 0.561 ± 0.036 |
| Threonine | 0.315 ± 0.008 | 0.327 ± 0.002 | 0.328 ± 0.007 |
| Valine | 0.500 ± 0.011 | 0.476 ± 0.032 | 0.518 ± 0.010 |
| Cysteine | 0.188 ± 0.002 b | 0.190 ± 0.005 b (+1.06) | 0.198 ± 0.002 a (+5.32) |
| Tyrosine | 0.243 ± 0.001 | 0.255 ± 0.002 | 0.260 ± 0.022 |
| E/T (%) | 43.5 | 43.2 | 42.7 |
| Non-essential amino acids (NEA) | |||
| Alanine | 0.801 ± 0.020 ab | 0.785 ± 0.010 b (− 2.00) | 0.830 ± 0.018 a (+3.62) |
| Arginine | 0.283 ± 0.014 | 0.289 ± 0.011 | 0.282 ± 0.007 |
| Aspartic acid | 0.586 ± 0.023 c | 0.656 ± 0.008 b (+11.95) | 0.707 ± 0.037 a (+20.65) |
| Glutamic acid | 1.898 ± 0.103 | 1.861 ± 0.015 | 1.962 ± 0.066 |
| Glycine | 0.317 ± 0.004 | 0.303 ± 0.002 | 0.323 ± 0.014 |
| Hydroxyproline | 0.004 ± 0 | 0.005 ± 0.001 | 0.005 ± 0 |
| Proline | 0.782 ± 0 c | 0.852 ± 0.023 b (+8.95) | 0.908 ± 0.021 a (+16.11) |
| Serine | 0.437 ± 0.002 b | 0.447 ± 0.011 ab (+2.29) | 0.464 ± 0.012 a (+6.18) |
| Total | 9.040 ± 0.190 b | 9.142 ± 0.142 b (+1.13) | 9.564 ± 0.056 a (+5.80) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchini, M.; Marti, A.; Folli, C.; Prandi, B.; Ganino, T.; Conte, P.; Fadda, C.; Mattarozzi, M.; Carini, E. Sprouting of Sorghum (Sorghum bicolor [L.] Moench): Effect of Drying Treatment on Protein and Starch Features. Foods 2021, 10, 407. https://doi.org/10.3390/foods10020407
Marchini M, Marti A, Folli C, Prandi B, Ganino T, Conte P, Fadda C, Mattarozzi M, Carini E. Sprouting of Sorghum (Sorghum bicolor [L.] Moench): Effect of Drying Treatment on Protein and Starch Features. Foods. 2021; 10(2):407. https://doi.org/10.3390/foods10020407
Chicago/Turabian StyleMarchini, Mia, Alessandra Marti, Claudia Folli, Barbara Prandi, Tommaso Ganino, Paola Conte, Costantino Fadda, Monica Mattarozzi, and Eleonora Carini. 2021. "Sprouting of Sorghum (Sorghum bicolor [L.] Moench): Effect of Drying Treatment on Protein and Starch Features" Foods 10, no. 2: 407. https://doi.org/10.3390/foods10020407
APA StyleMarchini, M., Marti, A., Folli, C., Prandi, B., Ganino, T., Conte, P., Fadda, C., Mattarozzi, M., & Carini, E. (2021). Sprouting of Sorghum (Sorghum bicolor [L.] Moench): Effect of Drying Treatment on Protein and Starch Features. Foods, 10(2), 407. https://doi.org/10.3390/foods10020407