
The Effect of pH and Storage Temperature on the Stability of Emulsions Stabilized by Rapeseed Proteins




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
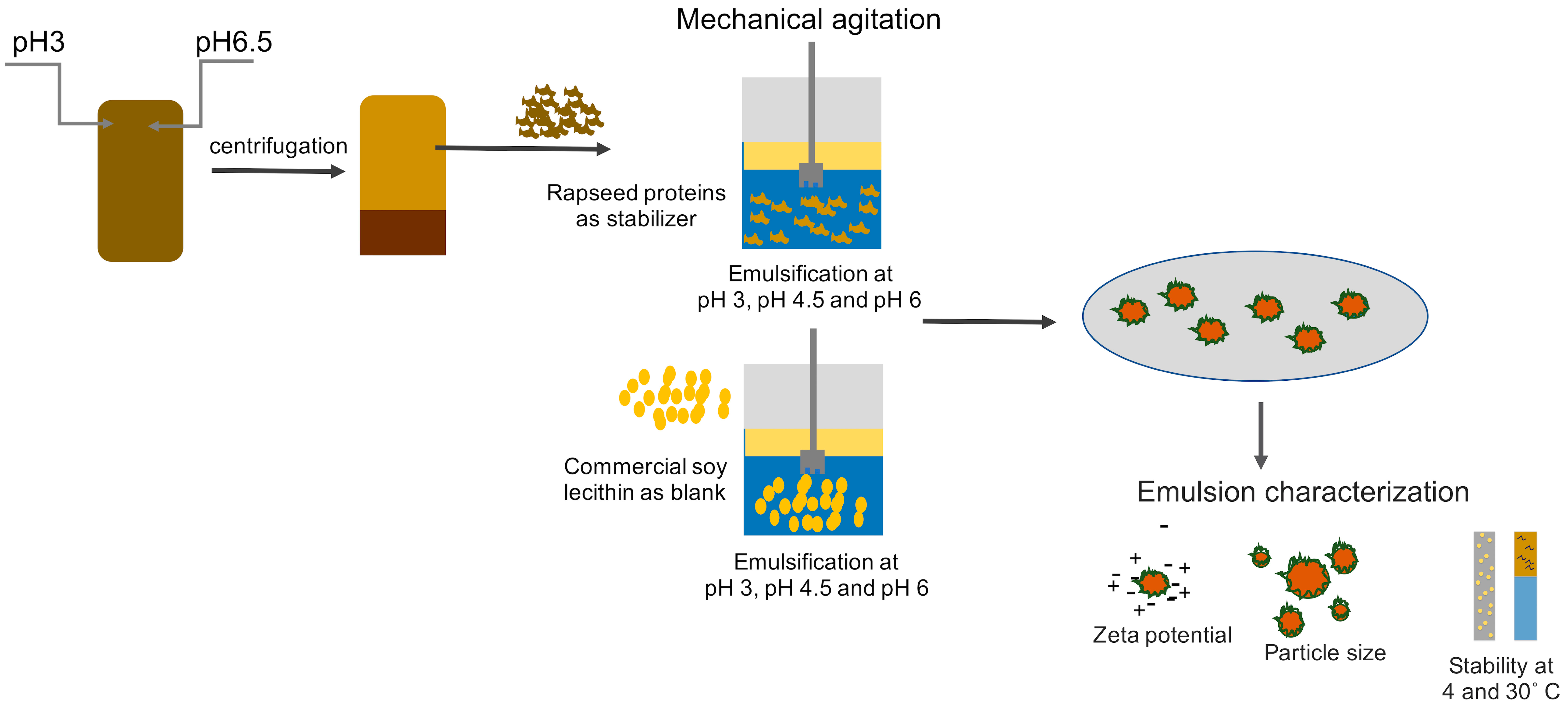
2. Materials and Methods
2.1. Sources of Materials and Chemicals
2.2. Production of Rapeseed Protein Concentrates
2.3. Emulsion Formulation
2.4. Particle Size Analysis
2.5. Zeta Potential
2.6. Surface Tension
2.7. Emulsion Stability over Time in Different Storage Temperatures
2.8. Statistical Analysis
3. Results and Discussion
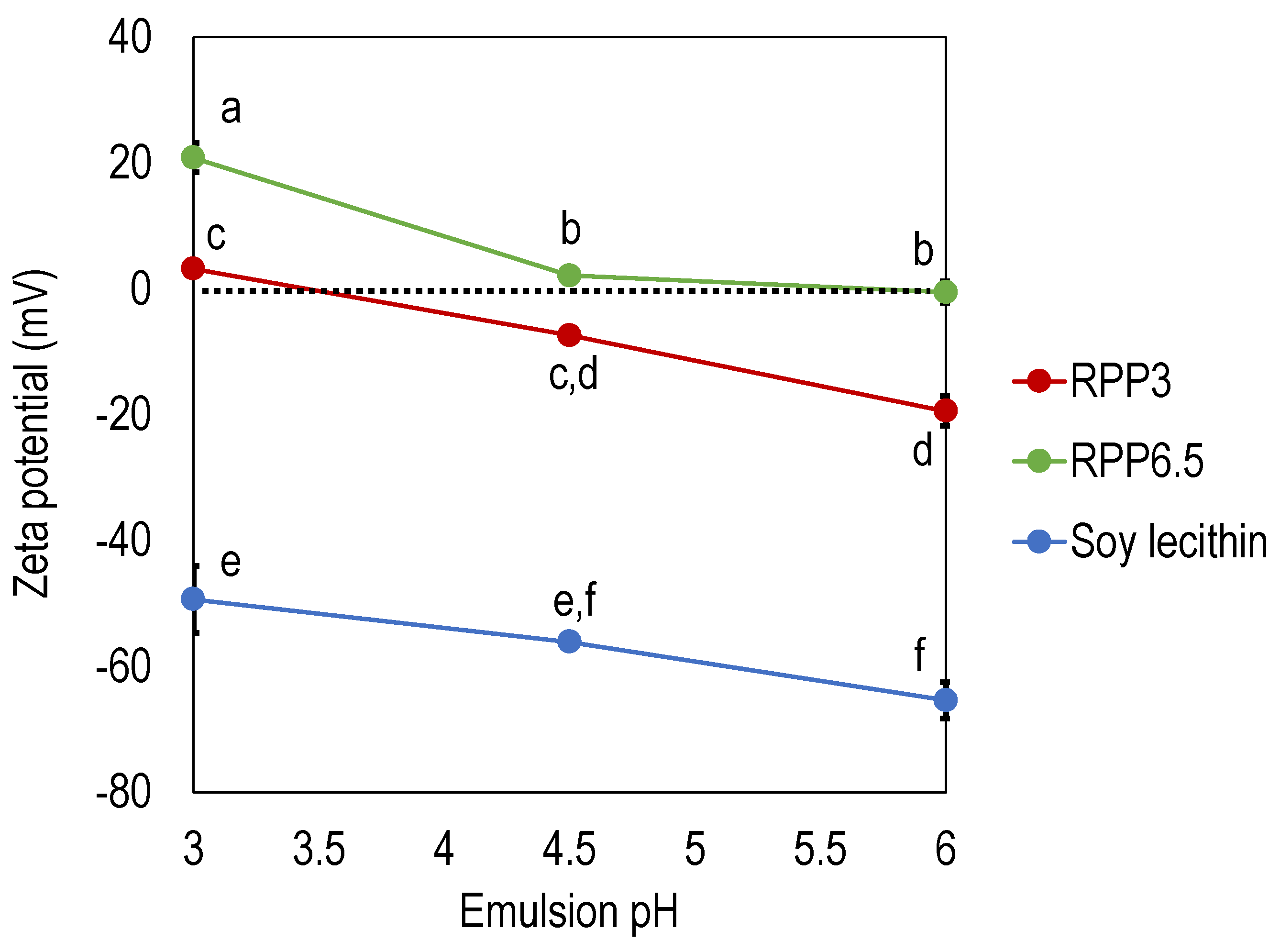
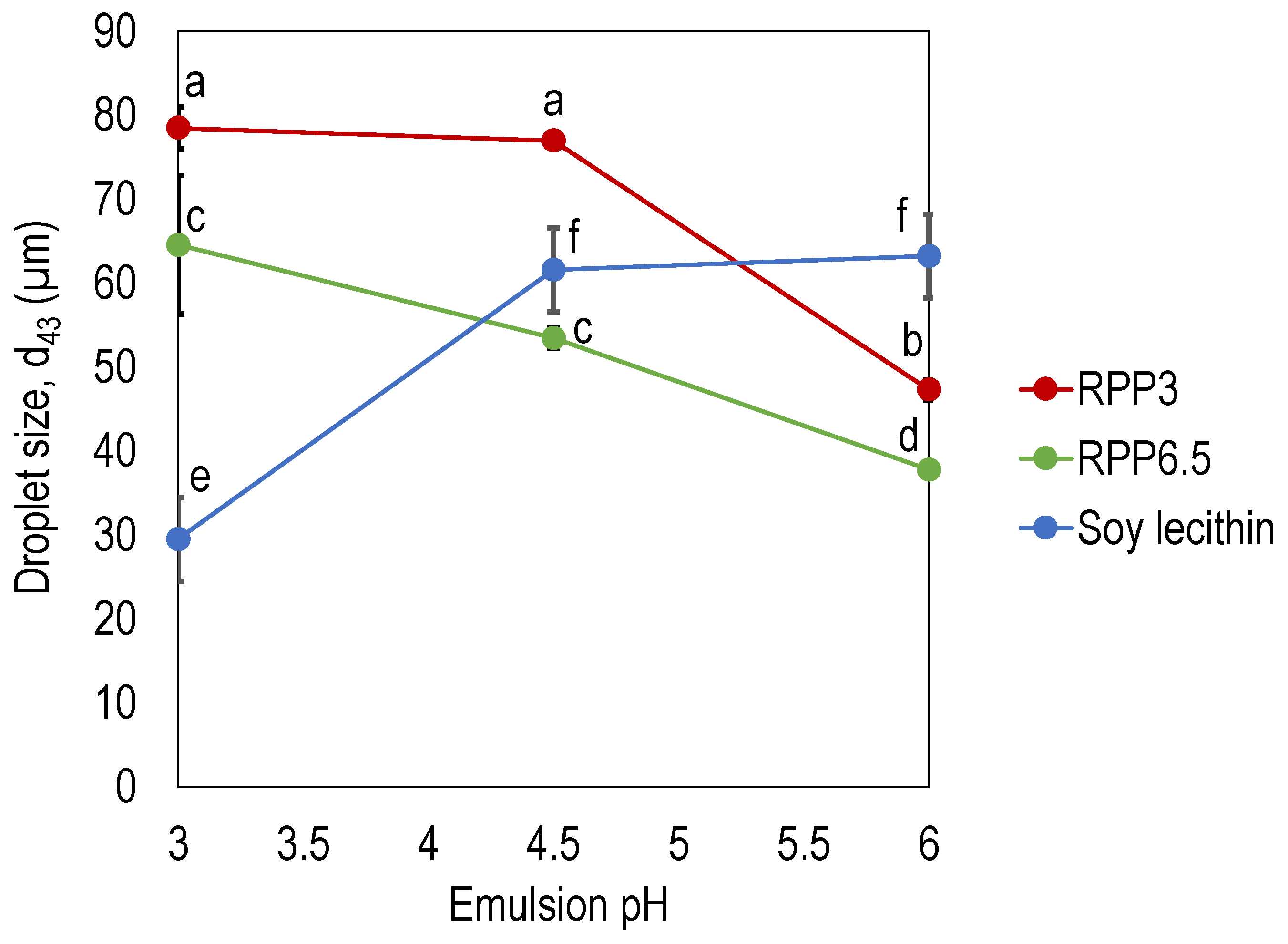
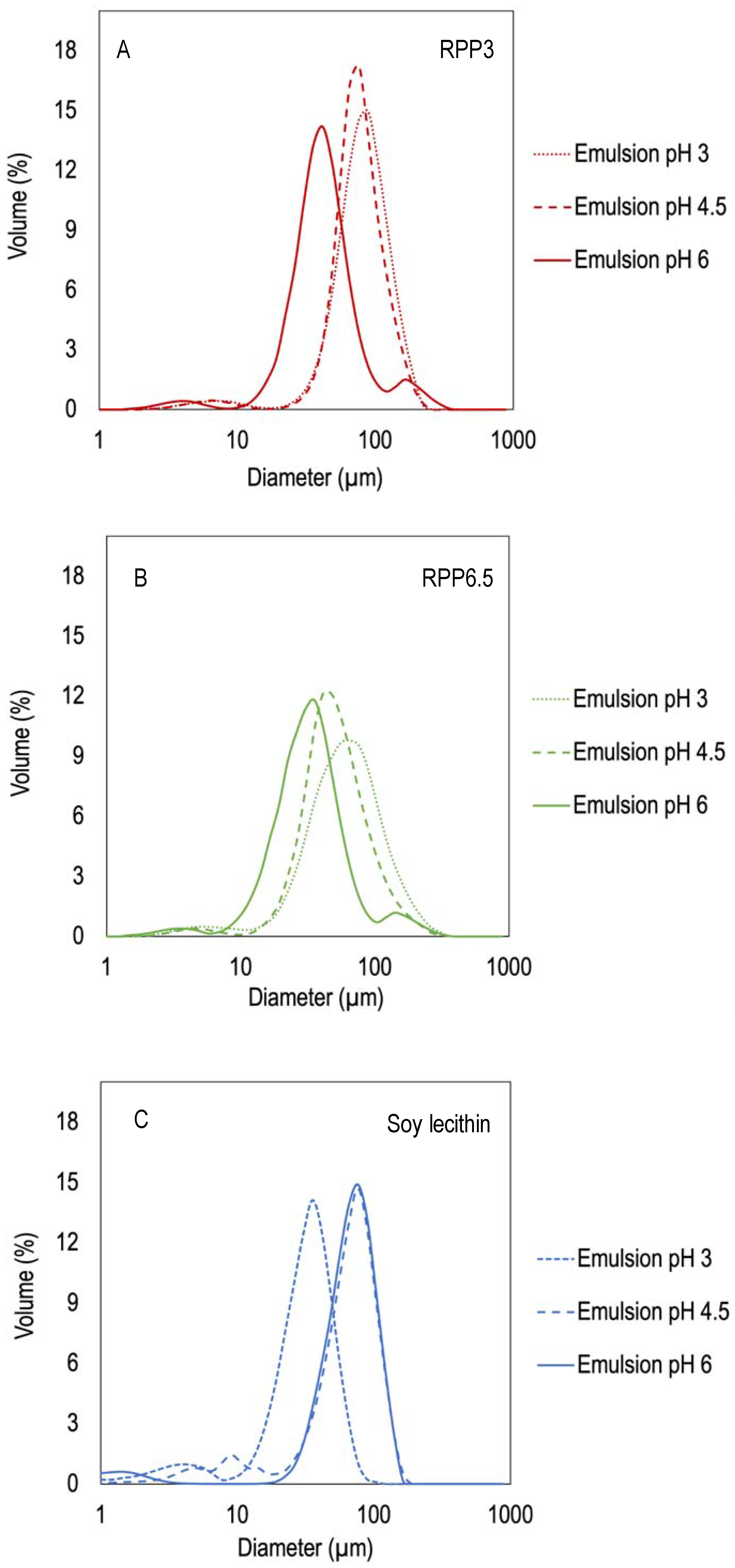
3.1. Effect of Precipitation pH on Emulsifying Properties
3.1.1. Zeta Potential
3.1.2. Emulsion Droplet Size
3.1.3. Droplet Size Distribution
3.1.4. Stability Mechanisms
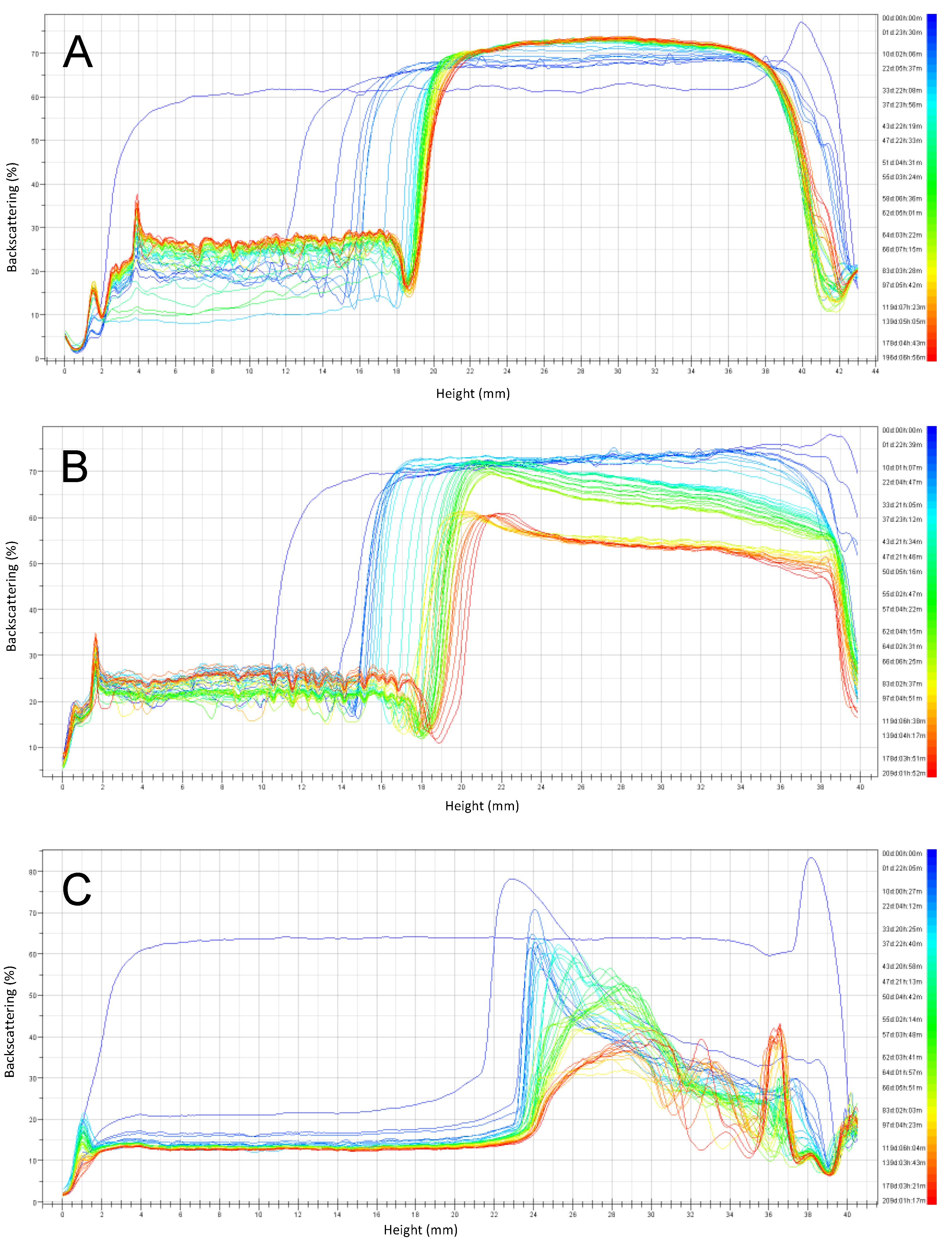
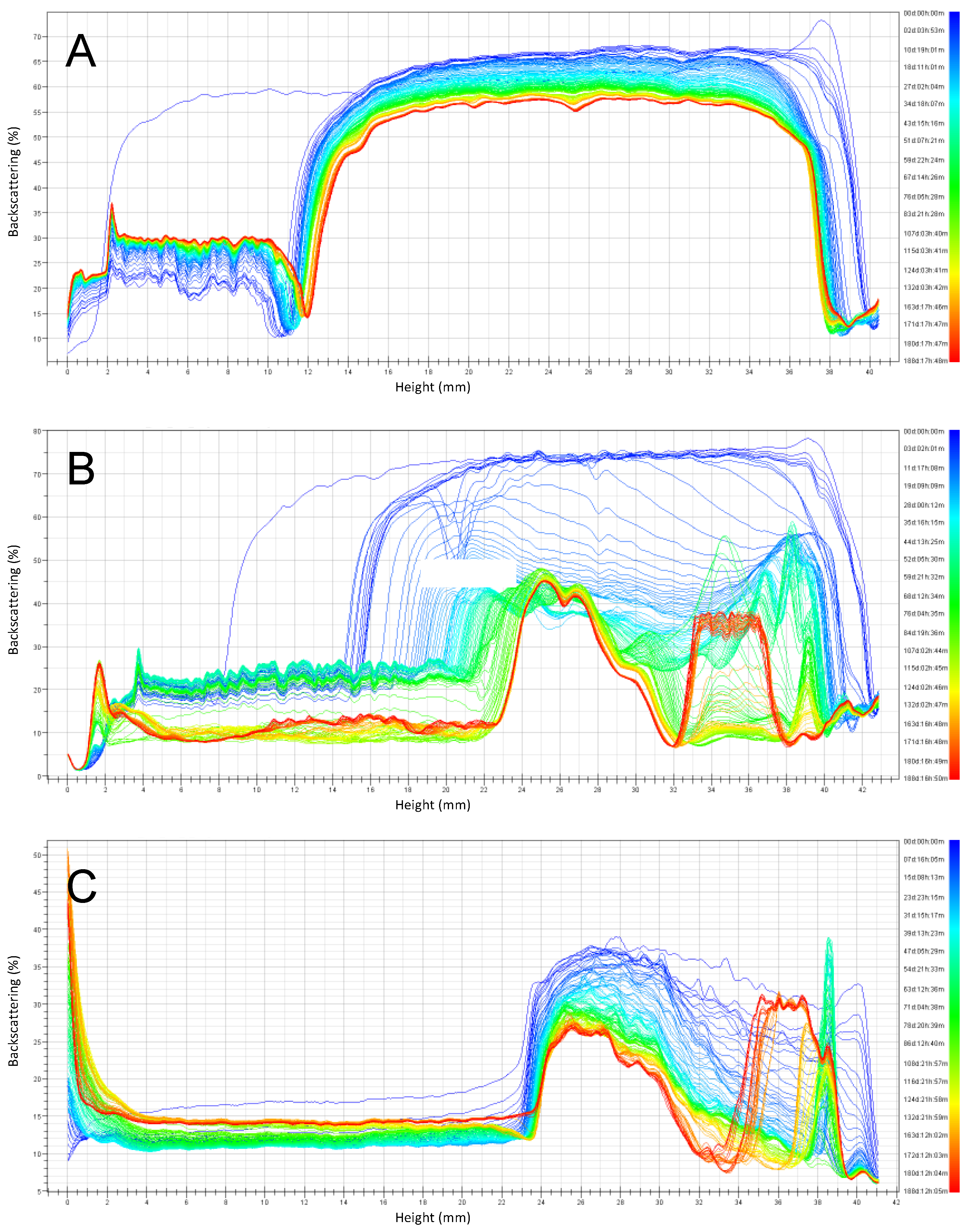
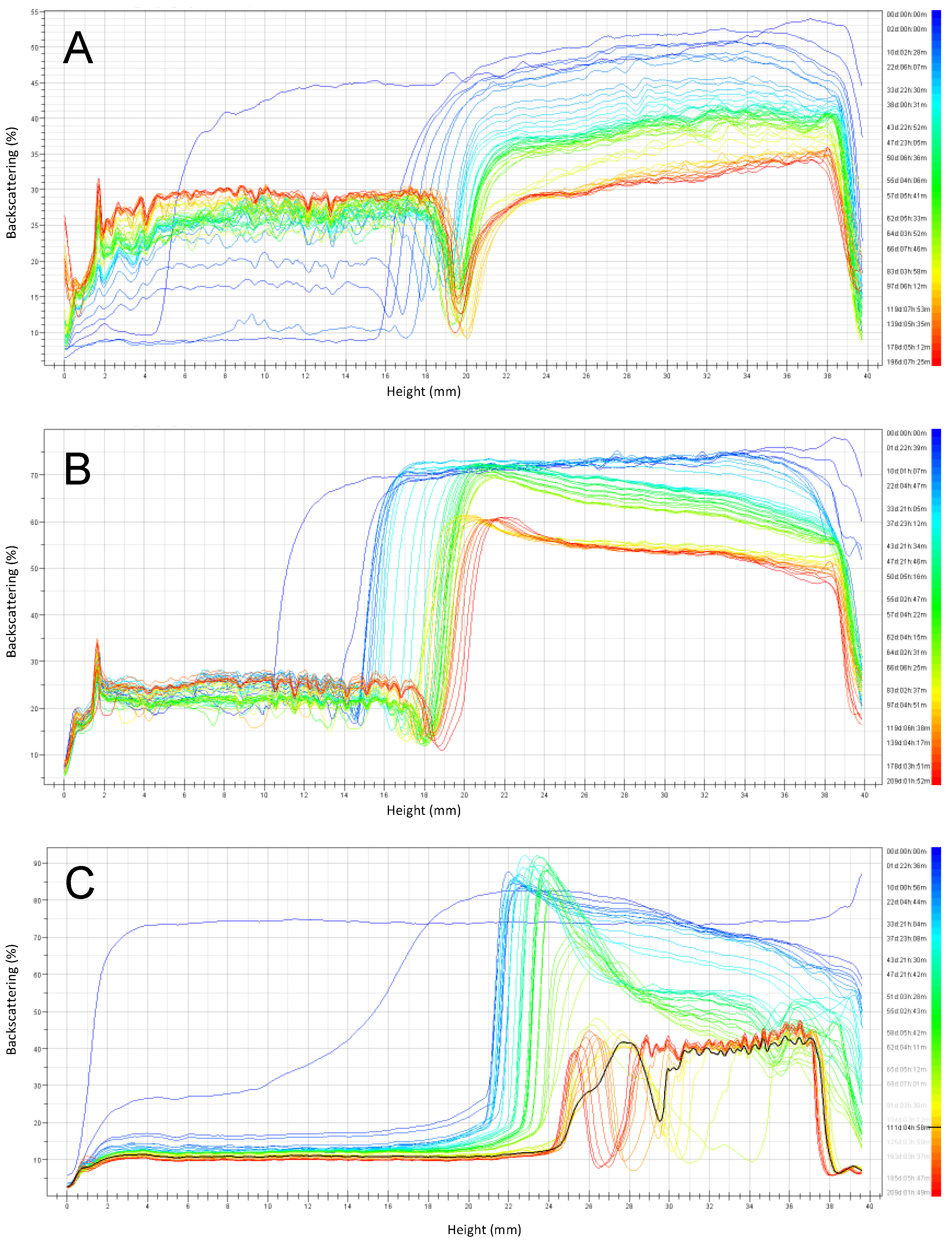
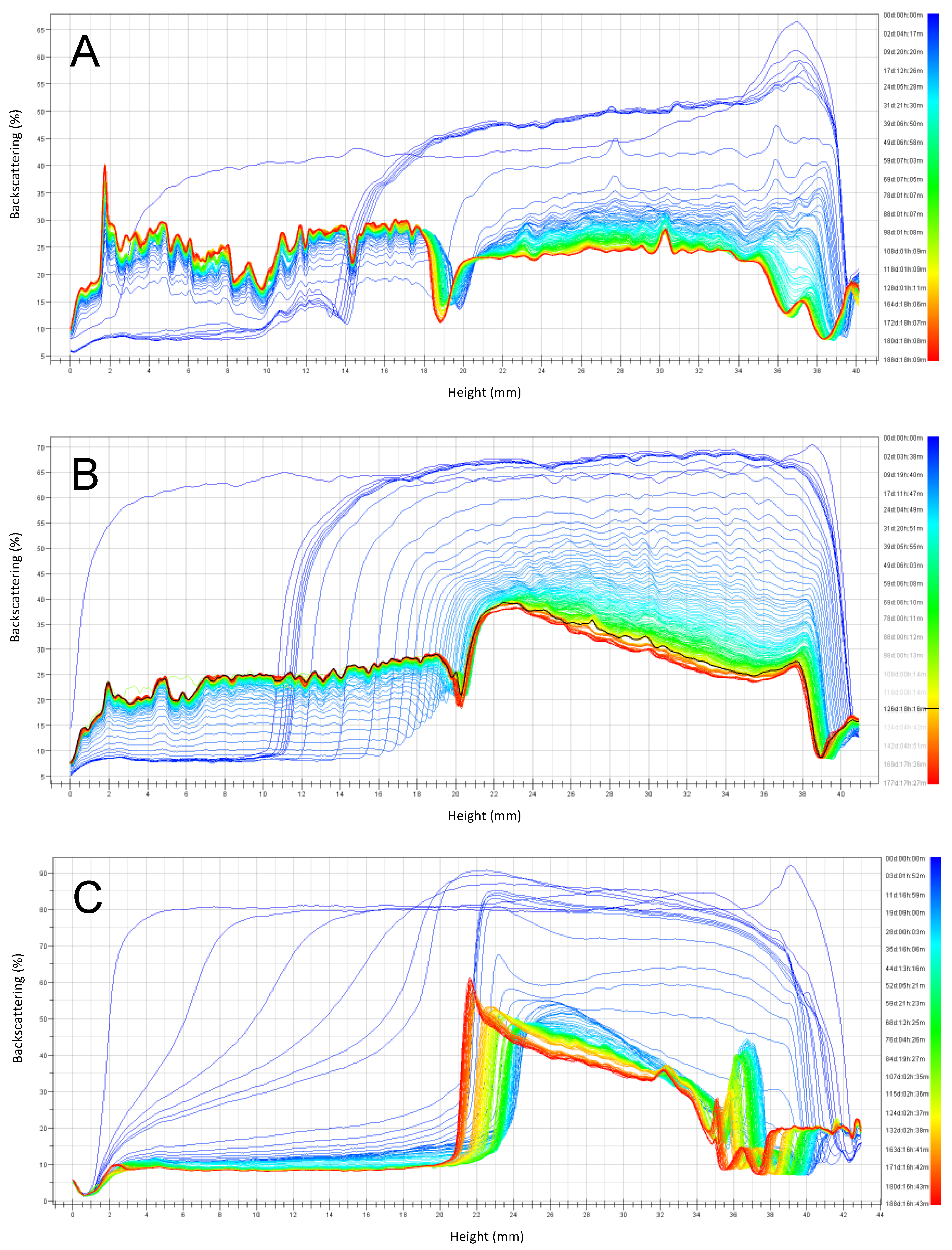
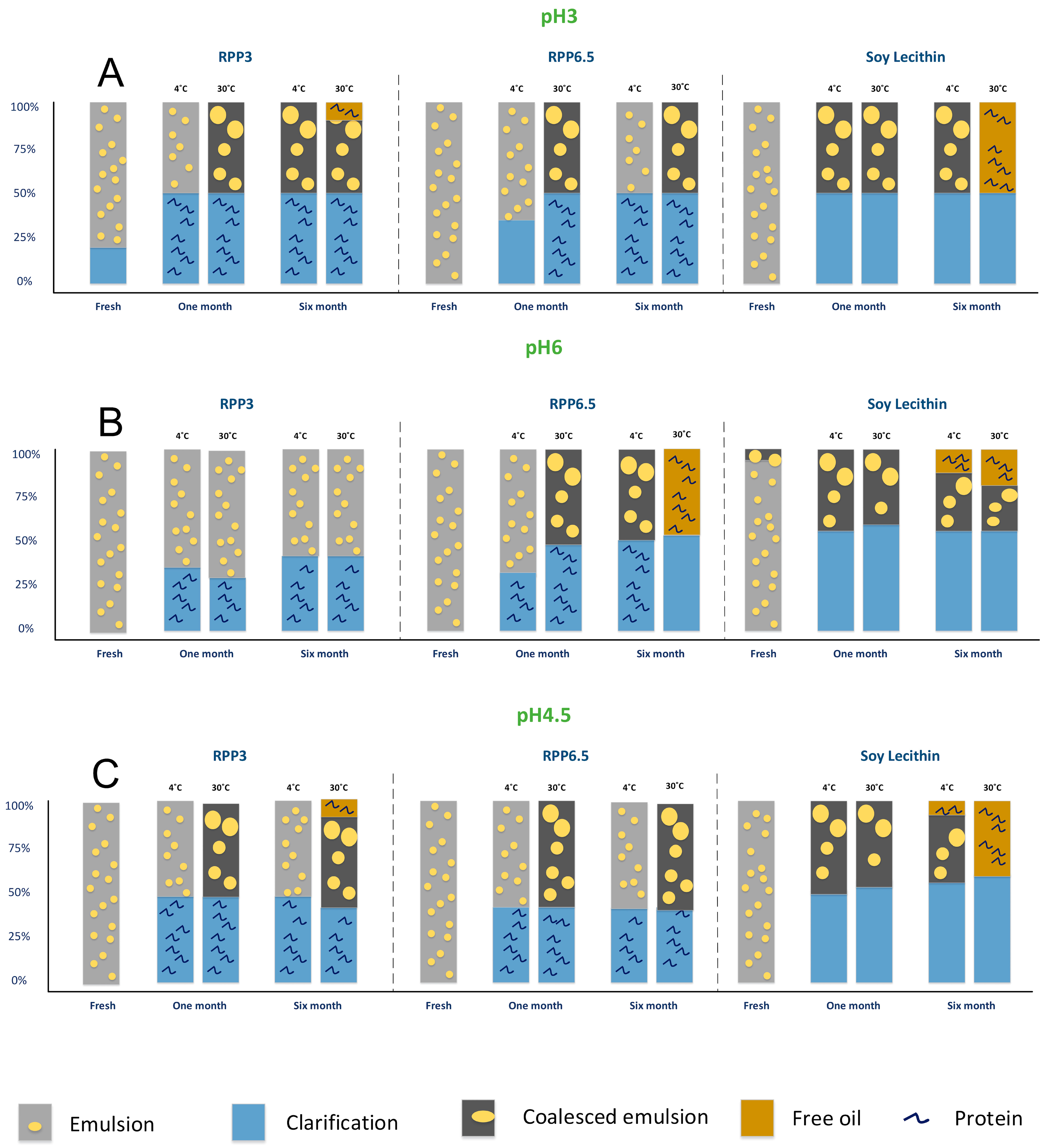
3.2. Emulsion Stability during Six Months of Storage
3.2.1. Emulsions with pH 6 Stored at 4 °C and 30 °C
3.2.2. Emulsions with pH 3 Stored at 4 °C and 30 °C
3.2.3. Summary of Stability as a Function of Emulsion pH and Storage Temperature
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- McClements, D.J. Critical review of techniques and methodologies for characterization of emulsion stability. Crit. Rev. Food Sci. Nutr. 2007, 47, 611–649. [Google Scholar] [CrossRef]
- Walstra, P. Physical Chemistry of Foods; Marcel Dekker: New York, NY, USA, 2003. [Google Scholar]
- McClements, D.J. Protein-stabilized emulsions. Curr. Opin. Colloid Interface Sci. 2004, 9, 305–313. [Google Scholar] [CrossRef]
- Karaca, A.C.; Low, N.; Nickerson, M. Emulsifying properties of canola and flaxseed protein isolates produced by isoelectric precipitation and salt extraction. Food Res. Int. 2011, 44, 2991–2998. [Google Scholar] [CrossRef]
- Wanasundara, J.P.; McIntosh, T.C.; Perera, S.P.; Withana-Gamage, T.S.; Mitra, P. Canola/rapeseed protein-functionality and nutrition. OCl 2016, 23, D407. [Google Scholar] [CrossRef] [Green Version]
- Aider, M.; Barbana, C. Canola proteins: Composition, extraction, functional properties, bioactivity, applications as a food ingredient and allergenicity–a practical and critical review. Trends Food Sci. Technol. 2011, 22, 21–39. [Google Scholar] [CrossRef]
- Huang, A. Oleosins and oil bodies in seeds and other organs. Plant. Physiol. 1996, 110, 1055. [Google Scholar] [CrossRef] [Green Version]
- Rayner, M. Current status on novel ways for stabilizing food dispersions by oleosins, particles and microgels. Curr. Opin. Food Sci. 2015, 3, 94–109. [Google Scholar] [CrossRef]
- Tzen, J.; Lie, G.; Huang, A. Characterization of the charged components and their topology on the surface of plant seed oil bodies. J. Biol. Chem. 1992, 267, 15626–15634. [Google Scholar] [CrossRef]
- Deleu, M.; Vaca-Medina, G.; Fabre, J.-F.; Roïz, J.; Valentin, R.; Mouloungui, Z. Interfacial properties of oleosins and phospholipids from rapeseed for the stability of oil bodies in aqueous medium. Colloids Surf. B 2010, 80, 125–132. [Google Scholar] [CrossRef]
- Tzen, J.T.; Cao, Y.; Laurent, P.; Ratnayake, C.; Huang, A.H. Lipids, proteins, and structure of seed oil bodies from diverse species. Plant. Physiol. 1993, 101, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Purkrtova, Z.; Jolivet, P.; Miquel, M.; Chardot, T. Structure and function of seed lipid body-associated proteins. Comptes Rendus Biol. 2008, 331, 746–754. [Google Scholar] [CrossRef]
- Wu, J.; Muir, A. Comparative structural, emulsifying, and biological properties of 2 major canola proteins, cruciferin and napin. J. Food Sci. 2008, 73, C210–C216. [Google Scholar] [CrossRef]
- Tan, S.H.; Mailer, R.J.; Blanchard, C.L.; Agboola, S.O. Emulsifying properties of proteins extracted from Australian canola meal. LWT 2014, 57, 376–382. [Google Scholar] [CrossRef]
- Ntone, E.; van Wesel, T.; Sagis, L.M.; Meinders, M.; Bitter, J.H.; Nikiforidis, C.V. Adsorption of rapeseed proteins at oil/water interfaces. Janus-like napins dominate the interface. J. Colloid Interface Sci. 2021, 583, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Krause, J.-P.; Schwenke, K. Behaviour of a protein isolate from rapeseed (Brassica napus) and its main protein components—Globulin and albumin—At air/solution and solid interfaces, and in emulsions. Colloids Surf. B 2001, 21, 29–36. [Google Scholar] [CrossRef]
- Chang, C.; Tu, S.; Ghosh, S.; Nickerson, M. Effect of pH on the inter-relationships between the physicochemical, interfacial and emulsifying properties for pea, soy, lentil and canola protein isolates. Food Res. Int. 2015, 77, 360–367. [Google Scholar] [CrossRef]
- Pirestani, S.; Nasirpour, A.; Keramat, J.; Desobry, S.; Jasniewski, J. Effect of glycosylation with gum Arabic by Maillard reaction in a liquid system on the emulsifying properties of canola protein isolate. Carbohydr. Polym. 2017, 157, 1620–1627. [Google Scholar] [CrossRef]
- Alashi, A.M.; Blanchard, C.L.; Mailer, R.J.; Agboola, S.O.; Mawson, J.A.; Aluko, R.E. Influence of enzymatic hydrolysis, pH and storage temperature on the emulsifying properties of canola protein isolate and hydrolysates. Int. J. Food Sci. Tech. 2018, 53, 2316–2324. [Google Scholar] [CrossRef]
- Wijesundera, C.; Boiteau, T.; Xu, X.; Shen, Z.; Watkins, P.; Logan, A. Stabilization of Fish Oil-In-Water Emulsions with Oleosin Extracted from Canola Meal. J. Food Sci. 2013, 78, C1340–C1347. [Google Scholar] [CrossRef] [PubMed]
- Becher, P. Liquid/Liquid Interfaces in: Encyclopedia of Emulsion Technology; Marcel Dekker: New York, NY, USA, 1983. [Google Scholar]
- Matos, M.; Lobo, A.; Benito, J.M.; Coca, J.; Pazos, C. Extending the useful life of metalworking fluids in a copper wire drawing industry by monitoring their functional properties. Tribol. Trans. 2012, 55, 685–692. [Google Scholar] [CrossRef]
- Tang, Y.R.; Ghosh, S. Stability and rheology of canola protein isolate-stabilized concentrated oil-in-water emulsions. Food Hydrocoll. 2021, 113, 106399. [Google Scholar] [CrossRef]
- Östbring, K.; Nilsson, K.; Ahlström, C.; Fridolfsson, A.; Rayner, M. Emulsifying and anti-oxidative properties of proteins extracted from industrially cold-pressed rapeseed press-cake. Foods 2020, 9, 678. [Google Scholar] [CrossRef]
- Züge, L.C.B.; Haminiuk, C.W.I.; Maciel, G.M.; Silveira, J.L.M.; de Paula Scheer, A. Catastrophic inversion and rheological behavior in soy lecithin and Tween 80 based food emulsions. J. Food Eng. 2013, 116, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Xu, Q.; Liu, X.; Xu, Y.; Yang, L.; Wang, S.; Li, J.; He, Y.; Liu, H. Soy glycinin-soyasaponin mixtures at oil–water interface: Interfacial behavior and O/W emulsion stability. Food Chem. 2020, 327, 127062. [Google Scholar] [CrossRef]
- Burgos-Díaz, C.; Wandersleben, T.; Marqués, A.M.; Rubilar, M. Multilayer emulsions stabilized by vegetable proteins and polysaccharides. Curr. Opin. Colloid Interface Sci. 2016, 25, 51–57. [Google Scholar] [CrossRef]
- Dalgleish, D.G. Adsorption of protein and the stability of emulsions. Trends Food Sci. Technol. 1997, 8, 1–6. [Google Scholar] [CrossRef]
- Liu, C.; Pei, R.; Peltonen, L.; Heinonen, M. Assembling of the interfacial layer affects the physical and oxidative stability of faba bean protein-stabilized oil-in-water emulsions with chitosan. Food Hydrocoll. 2020, 102, 105614. [Google Scholar] [CrossRef]
- Tong, X.; Cao, J.; Sun, M.; Liao, P.; Dai, S.; Cui, W.; Cheng, X.; Li, Y.; Jiang, L.; Wang, H. Physical and oxidative stability of oil-in-water (O/W) emulsions in the presence of protein (peptide): Characteristics analysis and bioinformatics prediction. LWT 2021, 149, 111782. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Östbring, K.; Matos, M.; Marefati, A.; Ahlström, C.; Gutiérrez, G. The Effect of pH and Storage Temperature on the Stability of Emulsions Stabilized by Rapeseed Proteins. Foods 2021, 10, 1657. https://doi.org/10.3390/foods10071657
Östbring K, Matos M, Marefati A, Ahlström C, Gutiérrez G. The Effect of pH and Storage Temperature on the Stability of Emulsions Stabilized by Rapeseed Proteins. Foods. 2021; 10(7):1657. https://doi.org/10.3390/foods10071657
Chicago/Turabian StyleÖstbring, Karolina, María Matos, Ali Marefati, Cecilia Ahlström, and Gemma Gutiérrez. 2021. "The Effect of pH and Storage Temperature on the Stability of Emulsions Stabilized by Rapeseed Proteins" Foods 10, no. 7: 1657. https://doi.org/10.3390/foods10071657
APA StyleÖstbring, K., Matos, M., Marefati, A., Ahlström, C., & Gutiérrez, G. (2021). The Effect of pH and Storage Temperature on the Stability of Emulsions Stabilized by Rapeseed Proteins. Foods, 10(7), 1657. https://doi.org/10.3390/foods10071657