3.3. Fundamental Rheological Parameters of Yellow Split Pea Fortified Wheat-Based Doughs
As expected, frequency sweep tests confirmed the elastic-like behavior of the tested wheat-based dough formulations, since the storage modulus (G′) was higher than the loss modulus (G″), while both moduli slightly increased and complex viscosity (η*) sharply decreased with frequency (
Figure 1a). As shown in
Table 3, for a selected frequency of 5.37 Hz, storage and loss moduli and complex viscosity shifted towards higher values, with an increasing level of roasted legume flour incorporated into the composite dough. Even though the values of the examined rheological parameters among the fortified systems did not show any significant difference compared to CON, the RYSP20 was characterized by significantly higher values compared to YSP10, which indicates the formation of a more elastic and firm dough. A structural modification of the YSP proteins due to roasting as well as moisture redistribution among the constituents of the composite doughs might enhance the gel network structure of the system. According to previous studies, the observed differences in the fortified doughs depend on the type and pre-processing of the flour used, as well as the fortification level. Specifically, the addition of 20% raw or germinated yellow pea and faba bean flours resulted in doughs with significantly higher G′ and G″ at the frequency of 10 Hz [
21]. Ahmed et al. [
22] also reported a concentration dependent effect, since the G′ and G′′ values increased as the concentration of lupine fiber in the composite doughs increased from 5 to 15%. In another study, addition of 30% flours from peas that had undergone different pre-treatments showed that doughs with germinated and toasted flours had significantly higher G′, G′′, and η*, whereas the dough with the raw pea did not differ from the control [
50]; those findings are consistent with these of the present work (
Table 3).
Regarding the creep-recovery test, the obtained rheological responses were typical of viscoelastic behavior, as expected for wheat-based doughs, showing the creep compliance data to slightly decrease with increasing roasted legume flour level (
Figure 1b). All rheological parameters calculated by fitting the creep-recovery test data to the Burgers model are summarized in
Table 3. The RYSP10 showed similar rheological properties with the CON sample, in both creep and recovery phases, whereas the YSP10, RYSP15, and RYSP20 exhibited significantly (
p < 0.05) lower maximum creep compliance (J
max) and viscoelastic compliance (J
m) in the creep phase. The YSP10 and RYSP20 were also characterized by significantly lower maximum creep strain compared to control. The RYSP20 dough showed the highest zero shear viscosity (η
o), which along with the lowest J
max, describes a dough with a greater resistance to flow and deformation, in agreement with the highest η* and G’ values of this sample, respectively. In previous studies, shifting of the maximum creep strain to lower values and η
o to higher values have been related to stronger doughs [
23]. It is worth noting here that the more elastic and viscous dough enhances the dough stiffness and is expected to be resistant to expansion upon proofing resulting in breads with low loaf volume. In the recovery phase, no differences were recorded among the samples for most of the measured parameters, apart from the RYSP20 sample, which exhibited significantly lower viscoelastic compliance (J
m) compared to control. The rheological properties of dough inevitably affect the textural characteristics of the final product, influencing bread quality, since the dough must be sufficiently elastic to allow the formation of the three-dimensional deformable network and strong enough to maintain the bubble structures formed during proofing and baking. Lupin fiber inclusion in a wheat dough has been found to exhibit a similar pattern, since at increased concentration (15% on flour basis), there was a significant decrease in maximum deformation, compared to lower levels of substitution [
22]. Overall, substitutions at 5 to 10% result in composite doughs with rheological properties similar to those of wheat flour doughs [
16,
17], while at higher substitution levels, with addition of vital wheat gluten, there is restoration of the rheological properties, as observed in the case of lentil-wheat composite doughs [
17].
3.4. Appearance of Yellow Split Pea Fortified Wheat-Based Breads
The appearance of the fortified wheat breads with yellow split pea flours was evaluated by loaf-specific volume measurements and crust and crumb color parameters. Loaf-specific volume was negatively affected by the higher level of legume flour addition, as the RYSP15 and RYSP20 exhibited significantly lower specific volume values than the CON sample (
Table 4 and
Figure 2). Additionally, the reduced loaf volume of the fortified breads with high levels of YSP flour was associated with a more compact macrostructure of the crumb (
Figure 2). Probably a stronger solid-like character and increased resistance to flow and deformation of the fortified doughs (
Table 3) led to increased stiffness, resistance to expansion, and ability to regain their initial shape and thereby caused reduced loaf volumes. The dilution of gluten network by incorporation of a non-wheat flour at high levels could also contribute to the decreased loaf volumes of the fortified breads. Similarly, in previous studies, the addition of legume flours from chickpeas, lentil, faba bean, carob bean, etc., at levels higher than 10% (on wheat flour basis), in most cases significantly reduced the loaf volume, regardless the type of the legume employed [
16,
18,
24,
25,
26]. However, at higher levels of legume inclusion, the addition of vital wheat gluten can improve the specific volume of the produced loaves, by restoring the rheological properties of the dough, as it has been shown with mixtures containing 20% lentil flour [
17], or 30% flour from chickpeas, lentils, peas, and soybeans [
25].
The crust and crumb color parameters of the examined breads are summarized in
Table 4. Inclusion of YSP flours in the formulations gradually reduced bread crust lightness (L*), with RYSP15 and RYSP20 being significantly less bright compared to CON. Moreover, an increase in redness (a*) was recorded in all composite breads, and at the same time, YSP10, RYSP15, and RYSPP20 exhibited significantly lower b* values (yellowness). In this context, there was a reduction in hue angle and Chroma values in crust with increasing level of yellow split pea flour substitution in the bread formulations; the former implies a shift to a redder and less yellow hue and the latter a change from a more saturated color (brighter) to a more achromatic (closer to grey) color. Regarding crumb color parameters, differences were noted only in the case of RYSP20, which showed significantly less bright (lower L* value), redder (higher a*), and yellow (higher b*) values than CON. More distinct differences in color parameters were noted in the bread crust samples, since they are attributed not only to the different color of legume flours added, but also to Maillard and caramelization reaction products, since the surface temperature during baking reaches ~180 °C, whereas the crumb temperature usually does not exceed 96 °C; furthermore, at the end of baking, the crust water activity reaches an intermediate value, ~0.75, which favors the development of Maillard reaction products compared to the high a
w values in crumb, ~0.97 [
40]. Similar results have been obtained for breads fortified with 20% flour from whole yellow pea that had been roasted at several temperatures, since, in all cases, crumb L* values were lower, whereas a* and b* values were higher than those of control sample [
28]. Consumers are not negatively affected by red and especially yellow color shades of breads, probably because they are related with familiar bakery ingredients/flours, such as red wheat, corn, etc.; i.e., despite the significant instrumentally observed color difference towards yellow, for pan breads fortified with thermally treated yellow split pea, the acceptability by the consumers did not appear to be affected [
53].
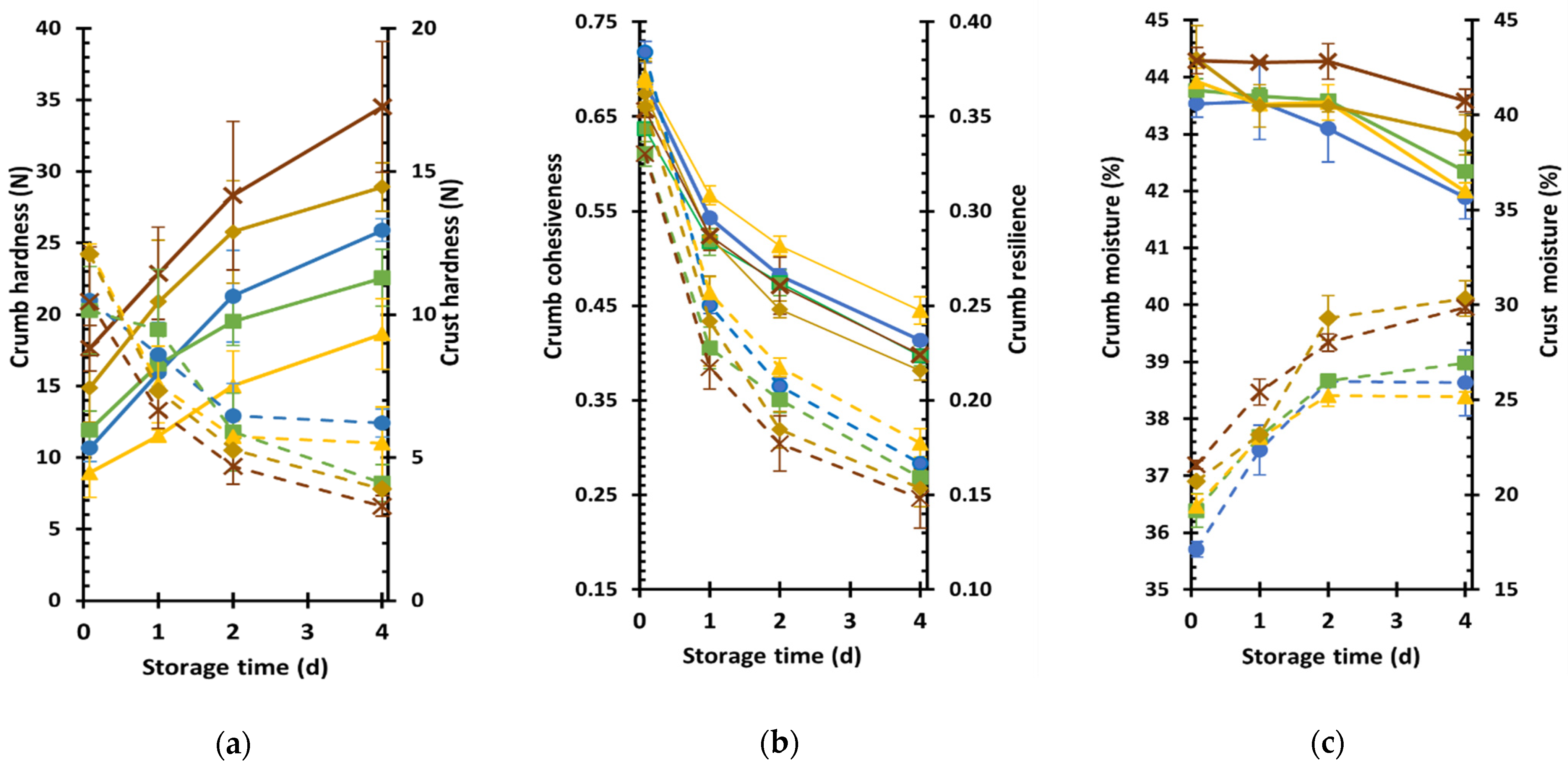
3.5. Textural Characteristics and Staling Kinetics of Yellow Split Pea Fortified Wheat-Based Breads
The textural characteristics of bread samples and their staling kinetics, as assessed by changes in crumb and crust textural attributes during product storage at 25 °C, are presented in
Table 5 and
Figure 3. The RYSP10 product exhibited the softest crumb throughout storage among the tested samples as indicated by the significantly lower hardness value, compared to those of RYSP15 and RYSP20, which had the higher hardness values among all samples (
Table 5,
Figure 3a). Additionally, the RYSP20 exhibited a significantly higher (
p < 0.05) hardening rate compared to YSP10 and RYSP10, indicating a quicker staling process. The increased crumb firmness of breads with the highest yellow split pea level is consistent with its reduced loaf volumes and rather compact crumb structure (
Table 4,
Figure 2); significant negative correlations between loaf volume of breads and crumb hardness evaluated by compression testing have been previously found [
54,
55]. In a relevant study, lentil–wheat composite breads exhibited harder crumb texture than the control sample, even at 5% inclusion level, although it has been proposed that a simultaneous use of gluten in the mixture might allow lentil substitution levels up to 15%, without negatively affecting crumb softness [
17].
Cohesiveness and springiness values were similar among all fresh samples, while the RYSP20 crumb was less resilient compared to CON (
Table 5). Resilience decreased during storage, as shown in
Figure 3b, indicating the loss of elasticity due to the staling process. Fresh RYSP20 loaves also exhibited the highest chewiness among all tested bread formulations, although during staling, its chewiness increase rate was the lowest among samples (
Table 5). The high level of legume flour inclusion in RYSP20 formulation, which can lead to a substantial decrease in gluten concentration and rather large interference in the development of a well-structured wheat protein network by the proteins and dietary fibers of yellow split pea, might have contributed to the formation of a harder and less elastic crumb.
Overall, the staling process during storage negatively affects bread texture resulting in harder, less elastic, and crumblier crumb (
Figure 3a,b); the latter is evident by the decrease in crumb cohesiveness values with increasing storage time. Staling is a complex phenomenon attributed to several mechanisms that occur in bakery items; the most important ones being gluten dehydration, leading to the transition of gluten from a rubbery to a glassy state [
56], amylopectin retrogradation, and water transfer from crumb to crust [
57]. Finally, all fresh samples exhibited similar crust hardness levels (
Table 5), indicating that the applied substitution levels did not have any substantial effect on this parameter. Nevertheless, crust softening upon bread storage (
Figure 3a) due to staling occurred at a higher rate for bread containing raw YSP than the control, while the inclusion of roasted YPS into the bread formulation also appeared to increase the crust softening rate, but not in a concentration-dependent manner (
Table 5). It is worthy to note that findings of the present study are referred to the samples prepared under the conditions described in this work and for the specific commercial yellow split pea used. There are several factors that could affect the quality characteristics of bakery products supplemented with legume seeds, such as variety, growth location, storage time [
58], after harvest, and after milling of the seeds as well as flour particle size [
40,
47].
Crust softening during staling was also evidenced (
Figure 3c) and is generally ascribed to water redistribution between crust and crumb. In fresh breads, crumb moisture content did not differ among the samples, while an increase in crust moisture with an increasing level of YSP flour into the bread formulation was noted probably due to the increasing content of proteins and dietary fibers that enhance water retention (
Table 5); at the end of storage, the fortified breads with RYSP15 and RYSP20 had considerably higher crust moisture compared to control sample (
Figure 3c). Instead, fortification of breads with YSP at any level did not seem to have an impact on moisture gain rate of crust, whereas the moisture loss rate of crumb decreased significantly compared to control bread, only in case of the highest fortification level (RYSP20) (
Table 5), resulting in a product with the highest moisture content at the end of storage (
Figure 3c).
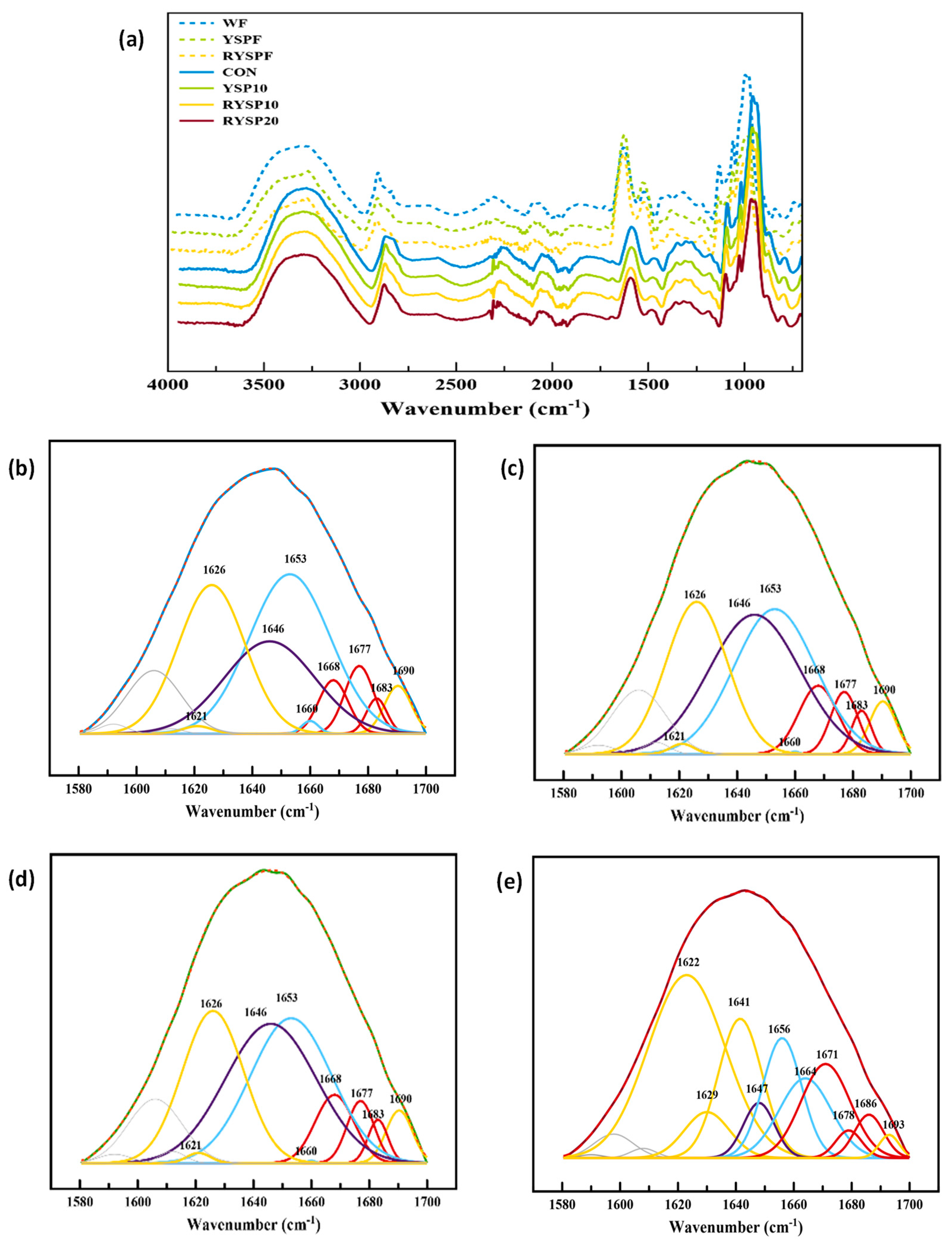
3.7. FTIR Spectroscopy Analysis of Yellow Split Pea Fortified Wheat-Based Flours and Breads
Figure 5a presents the FTIR normalized full spectra of wheat and raw or roasted yellow split pea flours and crumb samples of fresh breads. There are three distinct regions in all spectra. The bands around 2800–3000 cm
−1 are attributed to the C-H stretching modes, and the broad band at ~3300 cm
−1 corresponds to intermolecular H-bonding (O-H stretching modes) [
44]. As was expected, the broad peak in the region of -OH vibrations was less intense in the case of roasted YSP flour, probably, as a response to the loss of water due to the roasting process of the YSP flour (
Figure 5a); RYSPF flour had 8.3% moisture content versus 10.3% of the YSPF. The region at 1700–1500 cm
−1 is characteristic of the presence of protein molecules and is attributed to the Amide I (80% C = O stretch, 10% C-N stretch) and Amide II (60%N-H bend, 30%C-N stretch, and 10% C-C stretch) bond vibrations [
44,
62]. The higher protein content of YSPF (27.0%) and RYSPF (27.5%) flours compared to wheat flour alone (11.1%) was reflected in the higher intensity of the bands in this region observed for the legume flour samples (
Figure 5a).
Deconvolution of the Amide I peak of the flour spectra yielded several peaks attributed to different secondary structures of the protein components (
Figure 5b–e). CON, YSP10, and RYSP10 Amide I band deconvoluted to peaks centered at 1621, 1626, and 1690 cm
−1, assigned to the β-sheet structure, peaks centered at 1668, 1677, and 1683 cm
−1, assigned to the β-turn structure, peaks centered at 1653 and 1660 cm
−1, assigned to α-helix structure, and a peak centered at 1646 cm
−1, assigned to random coil structures. The RYSP20 sample revealed peaks centered at 1622, 1629, and 1693 cm
−1 for β-sheet structure, peaks centered at 1671, 1678, and 1686 cm
−1 for β-turn structure, peaks centered at 1656 and 1664 cm
−1 for α-helix structure, and a peak centered at 1647 cm
−1 for random coil structure [
44,
45,
46]. The estimated protein secondary structures of the flours are given in
Table 7. The protein conformation of the β-sheet structure was the most abundant structure in wheat flour (41.5%), which is in agreement with previously published data [
63,
64]. Raw and roasted YSP flours, also, had a high content in β-sheet structures, 47.8% and 45.4% for raw and roasted flour, respectively (
Table 7). Similarly, it has been reported that pea, lentil, and common bean flour have a high content of β-sheet conformations [
63,
65]. Roasting of yellow split pea flour led to a small conformational change in the proteins, seemingly, from the β-sheet structure (a decrease from 47.8% to 45.4%) towards random structure (an increase from 13.3% to 15.4%) (
Table 7), leading to a more disordered conformation [
65,
66]. This has been also reported for thermally treated common bean flours and was attributed to protein unfolding and denaturation events [
65]. No change for the α-helix of the YSP flour was noted after roasting, which was consistent with the results reported for dry gluten heated up to 85 °C [
67] and for thermally treated common bean flour at 120 °C for 24 h [
65]. The YSP flours exhibited a much higher content of the random structure and lower α-helices of proteins compared to wheat flour alone (
Table 7); other constituents in legume flours might disrupt the hydrogen bonding in some α-helices and destabilize towards random coil structures [
68].
Figure 5b–e presents the deconvolution and peak fitting procedure applied to the spectra region of Amide I to obtain the ratios of the secondary structures of proteins in crumb of fresh breads. As it is elucidated by the areas obtained from the Amide I band constituents, control and samples containing raw or roasted YSP at the 10% level exhibited similar secondary structures with almost equally distributed structures between random coil (24.5–28.5%), α-helix (25.2–29.1%) and β-sheet (25.8–27.9%), while the β-turn conformation was the least favored in these samples (10.7–11.7%) (
Table 7). However, incorporation of roasted YPS flour at the highest level (20%) into the bread formulation largely affected the protein secondary structure by greatly increasing β-sheets (52.2%) and β-turns (26.0%), at the expense of α-helix (10.9%) and random (3.9%) conformation, i.e., the increase in β-sheets has been associated with dehydration of gluten that results in chain aggregation through intermolecular β-sheets [
44,
45,
69]. The increase in β-sheets could also be partly attributed to legume protein aggregation due to their possible denaturation following the roasting treatment. Concerning gluten aggregation, it seems feasible to occur in the case of RYSP20 bread, since the YSP flour at 20% greatly fortified this formulation with dietary fibers and proteins (
Table 8) that may cause dehydration of gluten due to competition for water with these polymers; i.e., gluten dehydration could strengthen the macrostructure of crumb resulting in low volume [
70], hardening of the crumb, and fast staling [
71], phenomena in agreement with the data presented in
Table 5 and
Figure 3. In previous studies, it has been found that fortification of dough and bread formulations with flours or flour fractions or concentrates enriched in dietary fibers resulted in water redistribution, which is accompanied by a shift of the secondary structure of gluten from β-spiral (consecutive β-turns) to β-sheet conformation [
44,
45,
69,
70]; such a conformational transition has adverse effects on bread quality. In our study, the increase in β-sheets in secondary protein structure of the RYSP20 bread sample is accompanied with an increase in β-turns as well (
Table 7). There was also a decrease in α-helices content and a large increase in β-sheets in the protein structures of RYSP20 bread compared to all other samples (
Table 7) that could at least in part be attributed to the presence of legume globulins, resulting in poor bread quality.
The evolution of the CON, YSP10, RYSP10, and RYSP20 protein secondary structures was also monitored with FTIR spectra through storage time (
Table 7). The protein chains in the crumb of CON, YSP10, and RYSP20 samples reorganized upon storage, as it was elucidated by decrease in α-helix conformation and an increase in random conformation in the case of CON and YSP10 from 0 day (fresh breads) until the end of storage (4th day). The available data about changes in protein secondary structure in bread upon storage are very limited, focusing mostly on the gliadin and glutenin fractions extracted from steamed bread [
66]. In the latter study, a downward trend in α-helix and β-turn content in the gliadin and glutenin components was noticed along with an opposite trend regarding β-sheet and random structures, as a result of the loss of moisture upon storage. In the present study, only the RYSP10 sample showed an increase in β-sheets, while the YPS10 and RYSP20 breads surprisingly showed small, but significant (
p < 0.05) increased estimates of β-turns. As reported by other investigators, for protein structures in a complex system, such as bread, conformational stabilities are very sensitive to interactions with the water molecules and presence of polysaccharides, mainly due to gluten dehydration upon kneading and baking [
44,
45,
72]. Sivam et al. [
69], suggested that the presence of other proteins, such as albumins, globulins, prolamins, glutelins, or non-starch polysaccharides, and fibers present in wheat flour should have an impact on the gluten protein network. Since legume flours, in general, contain such proteins, their inclusion in composite cereal flours is expected to further alter the gluten network. Rearrangements of the protein structure during storage of the baked product could be attributed to water molecule redistribution among the various constituents in the composite bread matrix as well as to water loss (
Table 5,
Figure 3).
The spectral features of flour and bread samples (
Figure 5a) around 1200–800 cm
−1 are attributed to the carbohydrate region and are associated with starch. The peaks around 1010–1020, 1080, and 1150 cm
−1 have been attributed to the coupled C-O and C-C stretching vibrations of the polysaccharide molecules [
44]. The higher starch content of wheat flour was reflected by the higher intensities of this area, compared to the raw and roasted YSP flours (
Figure 5a). The key bands at 1047 and 1022 cm
−1 have been previously assigned to short-range molecularly ordered or crystalline structures and amorphous forms of starch, respectively [
73,
74], and therefore, the ratio (R
1047/1022) of the intensities at 1047 and 1022 cm
−1 has been adopted as an indicator of the relative level of starch chain ordering [
62].
Table 7 presents the estimated R
1047/1022 values for the flours used in the present study. Crystallinity calculated by this index was not significantly (
p > 0.05) different between wheat and raw YSP flours being 0.455 for both samples; this value is lower than those reported for wheat and waxy maize starches (0.63 and 0.69, respectively) [
75] due to the presence of amorphous amylose in wheat and YSP starches of flours used in the present work. RYSPF displayed significantly (
p < 0.05) lower R
1047/1022 ratio (0.432) compared to YSPF probably due to the thermal processing, which may have caused some disordering of organized chains in starch granules.
Moreover, the ratio R
1047/1022 has been proposed as a relative index for monitoring starch retrogradation during bread storage and, thus, product staling [
62,
74], as a result of formation of ordered structures involving starch chains (corresponding to 1047 cm
−1 band) and the loss of amorphous chain domains (corresponding to 1022 cm
−1). As expected, the increase in starch molecular ordering and the decrease in the amorphous starch structure, due to retrogradation events, were evidenced by the increased R
1047/1022 values calculated for the crumb during storage, although such an increase was pronounced only for the RYSP20 bread (
Table 7); the latter was consistent with the apparent melting enthalpy values of the retrograded amylopectin, ΔH
ret, and the retrogradation index, RI, values (
Figure 4b,
Table 6) of this sample, as assessed from the DSC data. Additionally, the RYSP20 bread exhibited a significantly higher rate of increase for the R
1047/1022 index during product storage compared to other samples (
Table 7). The greater the rate and extent of starch retrogradation during storage of breads fortified with roasted YSP flour at the highest level (20%), as evidenced by both FTIR and DSC analyses, are also consistent with the harder bread crumb (higher hardening rate) when compared to the other tested bread formulations (
Figure 3a,
Table 5). Similarly, other researchers have reported an increase in the R
1047/1022 value during the storage of bakery products due to starch retrogradation that also concurred with increased crumb hardness [
62,
73,
76,
77,
78].



 ,
, 

 ), YSP10 (
), YSP10 (  ), RYSP10 (
), RYSP10 (  ), RYSP15 (
), RYSP15 (  ), RYSP20 (
), RYSP20 (  ); notation of samples as in Table 1.
); notation of samples as in Table 1.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}