
Performance of Thermoplastic Extrusion, Germination, Fermentation, and Hydrolysis Techniques on Phenolic Compounds in Cereals and Pseudocereals

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
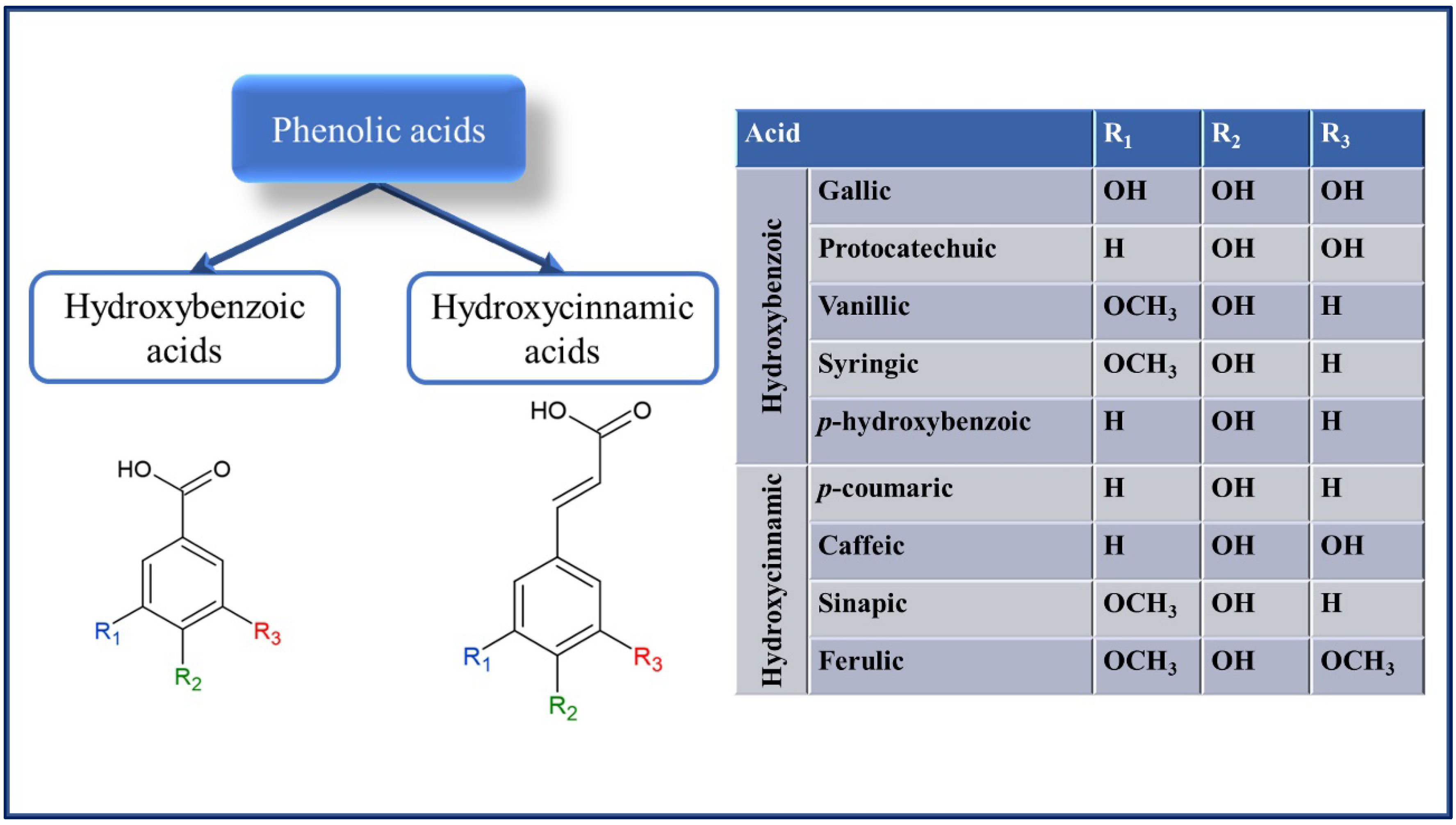
2. Phenolic Compounds Found in Cereals and Pseudocereals
3. Thermoplastic Extrusion
4. Germination
5. Fermentation and Enzymatic Hydrolysis
6. Conclusions and New Trends
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giuberti, G.; Rocchetti, G.; Lucini, L. Interactions between phenolic compounds, amylolytic enzymes and starch: An updated overview. Curr. Opin. Food Sci. 2020, 31, 102–113. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Barrow, C. Bioactive Compounds from Plant Origin: Extraction, Applications, and Potential Health Benefits, 1st ed.; Apple Academic Press: Palm Bay, FL, USA, 2020; 332p. [Google Scholar]
- Kasote, D.; Tiozon, R.N., Jr.; Sartagoda, K.J.D.; Itagi, H.; Roy, P.; Kohli, A.; Regina, A.; Sreenivasulu, N. Food processing technologies to develop functional foods with enriched bioactive phenolic compounds in cereals. Front. Plant Sci. 2021, 12, 771276. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.O.; Guido, L.F. A review on the fate of phenolic compounds during malting and brewing: Technological strategies and beer styles. Food Chem. 2022, 372, 131093. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.G.; Sonawane, S.K.; Arya, S.S. Cereal based functional beverages: A review. J. Microbiol. Biotechnol. Food Sci. 2018, 8, 914–919. [Google Scholar] [CrossRef]
- Ahmed, A.K.; Albalawi, Y.; Shora, H.Á.; Abdelseed, H.K.; Al-Kattan, N.A. Effects of quadruple therapy: Zinc, quercetin, bromelain and vitamin C on the clinical outcomes of patients infected with COVID-19. Int. J. Endocrinol. 2020, 1, 18–21. [Google Scholar]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Quercetin: Antiviral significance and possible COVID-19 integrative considerations. Nat. Prod. Commun. 2020, 15, 1934578X20976293. [Google Scholar] [CrossRef]
- Baroiu, L.; Dumitru, C.; Iancu, A.; Les, A.-C.; Drăgănescu, M.; Baroiu, N.; Anghel, L. COVID-19 impact on the liver. World J. Clin. Cases 2021, 9, 3814–3825. [Google Scholar] [CrossRef]
- George, A.S.; Brandl, M.T. Plant bioactive compounds as an intrinsic and sustainable tool to enhance the microbial safety of crops. Microorganisms 2021, 9, 2485. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Xiang, B.; Zhou, X.; Qin, D.; Li, C.; Xi, J. Infrared assisted extraction of bioactive compounds from plant materials: Current research and future prospect. Food Chem. 2022, 371, 131192. [Google Scholar] [CrossRef]
- Zhang, J.; Wen, C.; Zhang, H.; Duan, Y.; Ma, H. Recent advances in the extraction of bioactive compounds with subcritical water: A review. Trends Food Sci. Technol. 2020, 95, 183–195. [Google Scholar] [CrossRef]
- Yusoff, I.M.; Taher, Z.M.; Rahmat, Z.; Chua, L.S. A review of ultrasound-assisted extraction for plant bioactive compounds: Phenolics, flavonoids, thymols, saponins and proteins. Food Res. Int. 2022, 157, 111268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, L.; Fu, Y.; Jiang, J.-C. Bioactive constituents, nutritional benefits and woody food applications of Castanea mollissima: A comprehensive review. Food Chem. 2022, 393, 133380. [Google Scholar] [CrossRef]
- Dilrukshi, H.N.N.; Torrico, D.D.; Brennan, M.A.; Brennan, C.S. Effects of extrusion processing on the bioactive constituents, in vitro digestibility, amino acid composition, and antioxidant potential of novel gluten-free extruded snacks fortified with cowpea and whey protein concentrate. Food Chem. 2022, 389, 133107. [Google Scholar] [CrossRef] [PubMed]
- De La Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Phenolic Compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables, 1st ed.; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 253–271. [Google Scholar]
- Zhao, G.; Hu, M.; Lu, X.; Zhang, R. Soaking, heating and high hydrostatic pressure treatment degrade the flavonoids in rice bran. LWT Food Sci. Technol. 2022, 154, 112732. [Google Scholar] [CrossRef]
- Islam, M.Z.; Shim, M.-J.; Jeong, S.-Y.; Lee, Y.T. Effects of soaking and sprouting on bioactive compounds of black and red pigmented rice cultivars. Int. Food Sci. Technol. 2022, 57, 201–209. [Google Scholar] [CrossRef]
- Mir, N.A.; Riar, C.S.; Singh, S. Nutritional constituents of pseudo cereals and their potential use in food systems: A review. Trends Food Sci. Technol. 2018, 75, 170–180. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Rodriguez, J.M.L.; Barba, F.J.; Giuberti, G. Gluten-free flours from cereals, pseudocereals and legumes: Phenolic fingerprints and in vitro antioxidant properties. Food Chem. 2019, 271, 157–164. [Google Scholar] [CrossRef]
- Adebo, O.A.; Medina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, Y.; Li, G.; Savolainen, O.; Chen, Y.; Nielsen, J. De novo biosynthesis of bioactive isoflavonoides by engineered yeast cell factories. Nat. Commun. 2021, 12, 6085. [Google Scholar] [CrossRef]
- Wang, Q.; Li, L.; Wang, T.; Zheng, X. A review of extrusion-modified underutilized cereal flour: Chemical composition, functionality, and its modulation on starchy food quality. Food Chem. 2022, 370, 131361. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Deng, J.; Yang, K.; Zhu, Y.; Xia, C.; Chen, J.; Liu, T. Effect of processing on the release of phenolic compounds and antioxidant activity during in vitro digestion of hulless barley. Arab. J. Chem. 2021, 14, 103447. [Google Scholar] [CrossRef]
- Meza, S.L.R.; Sinnecker, P.; Schmiele, M.; Massaretto, I.L.; Chang, Y.K.; Marquez, U.M.L. Production of innovative gluten-free breakfast cereals based on red and black rice by extrusion processing technology. J. Food Sci. Technol. 2019, 56, 4855–4866. [Google Scholar] [CrossRef] [PubMed]
- Chávez, D.W.H.; Ascheri, J.L.R.; Carvalho, C.W.P.; Bernardo, C.O.; Deliza, R.; Freitas-Sá, D.G.C.; Teles, A.S.C.; Queiroz, V.A.V. Physicochemical properties, characteristics, and consumer acceptance of whole grain sorghum expanded extrudates. J. Food Process. Preserv. 2021, 45, e15837. [Google Scholar] [CrossRef]
- Abbas, Y.; Ahmad, A. Impact of processing on nutritional and antinutritional factors of legumes: A review. Ann. Food Sci. Technol. 2021, 20, 199–215. [Google Scholar]
- Meza, S.L.R.; Massaretto, I.L.; Sinnecker, P.; Schmiele, M.; Chang, Y.K.; Noldin, J.Á.; Marquez, U.M.L. Impact of thermoplastic extrusion process on chemical, nutritional, technological and sensory properties of gluten-free breakfast cereals from pigmented rice. Int. J. Food Sci. Technol. 2021, 56, 3218–3226. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, Y.; Dong, L.; Jia, X.; Liu, L.; Huang, F.; Chi, J.; Xiao, J.; Zhang, M.; Zhang, R. Extrusion and fungal fermentation change the profile and antioxidant activity of free and bound phenolics in rice bran together with the phenolic bioaccessibility. LWT Food Sci. Technol. 2019, 115, 108461. [Google Scholar] [CrossRef]
- D’Almeida, C.T.S.; Mameri, H.; Menezes, N.S.; Carvalho, C.W.P.; Queiroz, V.A.V.; Cameron, L.C.; Morel, M.-H.; Takeiti, C.Y.; Ferreira, M.S.L. Effect of extrusion and turmeric addition on phenolic compounds and kafirin properties in tannin and tannin-free sorghum. Food Res. Int. 2021, 149, 110663. [Google Scholar] [CrossRef]
- Ma, Y.J.; Guo, X.D.; Liu, H.; Xu, B.N.; Wang, M. Cooking, textural, sensorial, and antioxidant properties of common and tartary buckwheat noodles. Food Sci. Biotechnol. 2013, 22, 153–159. [Google Scholar] [CrossRef]
- Yalcin, S. Quality characteristics, mineral contents and phenolic compounds of gluten free buckwheat noodles. J. Food Sci. Technol. 2021, 58, 2661–2669. [Google Scholar] [CrossRef]
- Shao, Y.; Hua, Z.; Yu, Y.; Moua, R.; Zhu, Z.; Beta, T. Phenolic acids, anthocyanins, proanthocyanidins, antioxidant activity, minerals and their correlations in non-pigmented, red, and black rice. Food Chem. 2018, 239, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.C.; Lima, D.C.N.; Bailoni, M.A.; Risso, E.M.; Schmiele, M.; Steel, C.J.; Chang, Y.K. Physical characteristics, nutritional quality, and antioxidant potential of extrudates produced with polished rice and whole red bean flours. Cereal Chem. 2017, 94, 74–81. [Google Scholar] [CrossRef]
- Blandino, M.; Bresciani, A.; Loscalzo, M.; Vanara, F.; Marti, A. Extruded snacks from pigmented rice: Phenolic profile and physical properties. J. Cereal Sci. 2022, 103, 103347. [Google Scholar] [CrossRef]
- Melini, V.; Panfili, G.; Fratianni, A.; Acquistucci, R. Bioactive compounds in rice on Italian market: Pigmented varieties as a source of carotenoids, total phenolic compounds and anthocyanins, before and after cooking. Food Chem. 2019, 277, 119–127. [Google Scholar] [CrossRef]
- Amer, S.A.; Rizk, A.E. Production and evaluation of novel functional extruded corn snacks fortified with ginger, bay leaves and turmeric powder. Food Prod. Process. Nutr. 2022, 4, 4. [Google Scholar] [CrossRef]
- Dueñas, M.; Sánchez-Acevedo, T.; Alcalde-Eon, C.; Escribano-Bailón, M.T. Effects of different industrial processes on the phenolic composition of white and brown teff (Eragrostis tef (Zucc.) Trotter). Food Chem. 2021, 335, 127331. [Google Scholar] [CrossRef]
- Ortiz-Cruz, R.A.; Ramírez-Wong, B.; Ledesma-Osuna, A.I.; Torres-Chávez, P.I.; Sánchez-Machado, D.I.; Montaño-Leyva, B.; López-Cervantes, J.; Gutiérrez-Dorado, R. Effect of extrusion processing conditions on the phenolic compound content and antioxidant capacity of sorghum (Sorghum bicolor (L.) Moench) bran. Plant Foods Hum. Nutr. 2020, 75, 252–257. [Google Scholar] [CrossRef]
- Gong, K.; Chen, L.; Li, X.; Sun, L.; Liu, K. Effects of germination combined with extrusion on the nutritional composition, functional properties and polyphenol profile and related in vitro hypoglycemic effect of whole grain corn. J. Cereal Sci. 2018, 83, 1–8. [Google Scholar] [CrossRef]
- Bangar, S.P.; Sandhu, K.S.; Trif, M.; Rusu, A.; Pop, I.D.; Kumar, M. Enrichment in different health components of barley flour using twin-screw extrusion technology to support nutritionally balanced diets. Front. Nutr. 2022, 8, 823148. [Google Scholar] [CrossRef]
- Chávez, D.W.H.; Ascheri, J.L.R.; Carvalho, C.W.P.; Godoy, R.L.O.; Pacheco, S. Sorghum and roasted coffee blends as a novel extruded product: Bioactive compounds and antioxidant capacity. J. Funct. Foods 2017, 29, 93–103. [Google Scholar] [CrossRef]
- Song, J.; Shao, Y.; Chen, X.; Li, X. Release of characteristic phenolics of quinoa based on extrusion technique. Food Chem. 2022, 374, 128780. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.O.K.; Jeong, D.I.; Adnan, M.; Salitxay, T.; Heo, J.W.; Naznin, M.T.; Lim, J.D.; Cho, D.H.; Park, B.J.; Park, C.H. Effect of different processing methods on the accumulation of the phenolic compounds and antioxidant profile of broomcorn millet (Panicum miliaceum L.) flour. Foods 2019, 8, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.Z.; Yu, D.S.; Lee, Y.-T. The effect of heat processing on chemical composition and antioxidative activity of tea made from barley sprouts and wheat sprouts. J. Food Sci. 2019, 84, 1340–1345. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.Z.; Park, B.J.; Kang, H.-M.; Lee, Y.-T. Influence of selenium biofortification on the bioactive compounds and antioxidant activity of wheat microgreen extract. Food Chem. 2020, 309, 125763. [Google Scholar] [CrossRef]
- Müller, C.P.; Hoffmann, J.F.; Ferreira, C.D.; Diehl, G.W.; Rossi, R.C.; Ziegler, V. Effect of germination on nutritional and bioactive properties of red rice grains and its application in cupcake production. Int. J. Gastron. Food Sci. 2021, 25, 100379. [Google Scholar] [CrossRef]
- Chu, C. Dynamics of antioxidant activities, metabolites, phenolic acids, flavonoids, and phenolic biosynthetic genes in germinating Chinese wild rice (Zizania latifolia). Food Chem. 2020, 318, e126483. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Peñas, E.; Dueñas, M.; Frias, J.; Martínez-Villaluenga, C. Optimizing germination conditions to enhance the accumulation of bioactive compounds and the antioxidant activity of kiwicha (Amaranthus caudatus) using Response Surface Methodology. LWT Food Sci. Technol. 2017, 76, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Jan, R.; Riar, C.S.; Bansal, V. Analyzing the effect of germination on the pasting, rheological, morphological and in- vitro antioxidant characteristics of kodo millet flour and extracts. Food Chem. 2021, 361, 130073. [Google Scholar] [CrossRef]
- Dumitru, C.; Dinica, R.M.; Bahrim, G.-E.; Vizireanu, C.; Baroiu, L.; Iancu, A.V.; Draganescu, M. New insights into the antioxidant compounds of achenes and sprouted buckwheat cultivated in the Republic of Moldova. Appl. Sci. 2021, 11, 10230. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J.; Peñas, E. Optimization of germination time and temperature to maximize the content of bioactive compounds and the antioxidant activity of purple corn (Zea mays L.) by Response Surface Methodology. LWT Food Sci. Technol. 2017, 76, 236–244. [Google Scholar] [CrossRef]
- Gómez-Velázquez, H.D.J.; Aparicio-Fernández, X.; Reynoso-Camacho, R. Chia sprouts elicitation with salicylic acid and hydrogen peroxide to improve their phenolic content, antioxidant capacities in vitro and the antioxidant status in obese rats. Plant Foods Hum. Nutr. 2021, 76, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Tsafrakidou, P.; Michaelidou, A.-M.; Biliaderis, C.G. Fermented cereal-based products: Nutritional aspects, possible impact on gut microbiota and health implications. Foods 2020, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Melini, F.; Melini, V. Impact of fermentation on phenolic compounds and antioxidant capacity of quinoa. Fermentation 2021, 7, 20. [Google Scholar] [CrossRef]
- Roasa, J.; Villa, R.; Mine, Y.; Tsao, R. Phenolics of cereal, pulse and oilseed processing by-products and potential effects of solid-state fermentation on their bioaccessibility, bioavailability and health benefits: A review. Trends Food Sci. Technol. 2021, 116, 954–974. [Google Scholar] [CrossRef]
- Verni, M.; Verardo, V.; Rizzello, C.G. How fermentation affects the antioxidant properties of cereals and legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Zhang, Y.; Li, H.; Deng, Z.; Tsao, R. A review on insoluble-bound phenolics in plant-based food matrix and their contribution to human health with future perspectives. Trends Food Sci. Technol. 2020, 105, 347–362. [Google Scholar] [CrossRef]
- Ripari, V.; Bai, Y.; Gänzle, M.G. Metabolism of phenolic acids in whole wheat and rye malt sourdoughs. Food Microbiol. 2019, 77, 43–51. [Google Scholar] [CrossRef]
- Khan, S.A.; Zhang, M.; Liu, L.; Dong, L.; Ma, Y.; Wei, Z.; Chi, J.; Zhang, R. Co-culture submerged fermentation by Lactobacillus and yeast more effectively improved the profiles and bioaccessibility of phenolics in extruded brown rice than single-culture fermentation. Food Chem. 2020, 326, 126985. [Google Scholar] [CrossRef]
- Zielinski, H.; Honke, J.; Topolska, J.; Baczek, N.; Piskuła, M.K.; Wiczkowski, W.; Wronkowska, M. ACE inhibitory properties and phenolics profile of fermented flours and of baked and digested biscuits from buckwheat. Foods 2020, 9, 847. [Google Scholar] [CrossRef]
- Balli, D.; Bellumori, M.; Pucci, L.; Gabriele, M.; Longo, V.; Paoli, P.; Melani, F.; Mulinacci, N.; Innocenti, M. Does fermentation really increase the phenolic content in cereals? A study on millet. Foods 2020, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- Neves, N.A.; Gomes, P.T.G.; Carmo, E.M.R.; Silva, B.S.; Amaral, T.N.; Schmiele, M. Sourdough and jaboticaba (Plinia cauliflora) for improvement on pan bread characteristics. Res. Soc. Dev. 2020, 9, e90691110552. [Google Scholar] [CrossRef]
- Neves, N.A.; Gomes, P.T.G.; Schmiele, M. An exploratory study about the preparation and evaluation of sourdough breads with araticum pulp (Annona crassiflora Mart.). Res. Soc. Dev. 2020, 9, e956998036. [Google Scholar] [CrossRef]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Vodnar, D.C.; Teleky, B.-E.; Pop, C.R.; Stan, L.; Borsai, O.; Kadar, C.B.; Urcan, A.C.; et al. Quinoa sourdough fermented with Lactobacillus plantarum ATCC 8014 designed for gluten-free muffins—A powerful tool to enhance bioactive compounds. Appl. Sci. 2020, 10, 7140. [Google Scholar] [CrossRef]
- Luo, D.; Li, X.; Zhao, J.; Chen, G. Regulation of phenolic release in corn seeds (Zea mays L.) for improving their antioxidant activity by mix-culture fermentation with Monascus anka, Saccharomyces cerevisiae and Bacillus subtilis. J. Biotechnol. 2021, 325, 334–340. [Google Scholar] [CrossRef]
- Chen, G.; Liu, Y.; Zeng, J.; Tian, X.; Bei, Q.; Wu, Z. Enhancing three phenolic fractions of oats (Avena sativa L.) and their antioxidant activities by solid-state fermentation with Monascus anka and Bacillus subtilis. J. Cereal Sci. 2020, 93, 102940. [Google Scholar] [CrossRef]
- Bei, Q.; Wu, Z.; Chen, G. Dynamic changes in the phenolic composition and antioxidant activity of oats during simultaneous hydrolysis and fermentation. Food Chem. 2020, 305, 125269. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Material | Extrusion Conditions | TPC mg GAE/100 g | FPC mg GAE/100 g | BFC mg GAE/100 g | Reference |
|---|---|---|---|---|---|
| Sorghum bran | Unextruded | 2941.65 | 638.14 | 2303.52 | [39] |
| Mildest extrusion condition (feed moisture—30% and the 4th extrusion zone temperature—140 °C) | 3429.23 | 742.27 | 2686.96 | ||
| Most severe extrusion condition (feed moisture—26.46% and the 4th extrusion zone temperature—174.14 °C) | 2712.30 | 644.93 | 2470.64 | ||
| Yellow corn grits | Unextruded | 180.37 | - | - | [37] |
| Extruded with feed moisture (13%) and extrusion die temperature (180 °C) | 21.63 | - | - | ||
| Whole corn flour | Unextruded | 40.00 | 17.50 | 23.50 | [40] |
| Extruded with feed moisture (14%) and extrusion die temperature (120 °C) | 101.70 | 23.50 | 78.20 | ||
| Barley flour (BH-885) | Unextruded | 392.20 | - | - | [41] |
| Extruded with feed moisture (30%) and extrusion die temperature (150 °C) | 327.70 | - | - | ||
| Extruded with feed moisture (13%) and extrusion die temperature (180 °C) | 275.60 | - | - | ||
| Defatted rice bran | Unextruded | 206.80 | 19.06 | 187.70 | [29] |
| Extruded with feed moisture (25%) and extrusion zones temperature (70, 98 and 134 °C) | 320.02 | 47.39 | 272.64 | ||
| Sorghum flour with tannins (cultivar 9929026) * | Unextruded | 317.00 | 7.83 | 82.04 | [42] |
| Extruded with feed moisture (16%) and extrusion zones temperature (60, 120 and 140 °C) | 189.00 | 8.35 | 57.00 | ||
| Sorghum flour without tannins (cultivar 2012038) * | Unextruded | 139.00 | 6.13 | 85.59 | |
| Extruded with feed moisture (16%) and extrusion zones temperature (60, 120 and 140 °C) | 124.00 | 8.02 | 91.37 | ||
| Red quinoa flour | Unextruded | 1806.70 | 221.50 | 1585.20 | [43] |
| Extruded with feed moisture (25%) and extrusion die temperature (120 °C) | 1299.40 | 458.60 | 840.80 | ||
| Extruded with feed moisture (25%) and extrusion die temperature (140 °C) | 1182.90 | 410.40 | 772.50 | ||
| Extruded with feed moisture (25%) and extrusion die temperature (160 °C) | 1253.50 | 472.50 | 781.00 | ||
| Extruded with feed moisture (25%) and extrusion die temperature (180 °C) | 1233.40 | 431.70 | 801.70 | ||
| Broomcorn millet flour | Unextruded | 10.77 | - | - | [44] |
| Extruded with feed moisture (20%) and extrusion die temperature (110 °C) | 19.24 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paucar-Menacho, L.M.; Castillo-Martínez, W.E.; Simpalo-Lopez, W.D.; Verona-Ruiz, A.; Lavado-Cruz, A.; Martínez-Villaluenga, C.; Peñas, E.; Frias, J.; Schmiele, M. Performance of Thermoplastic Extrusion, Germination, Fermentation, and Hydrolysis Techniques on Phenolic Compounds in Cereals and Pseudocereals. Foods 2022, 11, 1957. https://doi.org/10.3390/foods11131957
Paucar-Menacho LM, Castillo-Martínez WE, Simpalo-Lopez WD, Verona-Ruiz A, Lavado-Cruz A, Martínez-Villaluenga C, Peñas E, Frias J, Schmiele M. Performance of Thermoplastic Extrusion, Germination, Fermentation, and Hydrolysis Techniques on Phenolic Compounds in Cereals and Pseudocereals. Foods. 2022; 11(13):1957. https://doi.org/10.3390/foods11131957
Chicago/Turabian StylePaucar-Menacho, Luz María, Williams Esteward Castillo-Martínez, Wilson Daniel Simpalo-Lopez, Anggie Verona-Ruiz, Alicia Lavado-Cruz, Cristina Martínez-Villaluenga, Elena Peñas, Juana Frias, and Marcio Schmiele. 2022. "Performance of Thermoplastic Extrusion, Germination, Fermentation, and Hydrolysis Techniques on Phenolic Compounds in Cereals and Pseudocereals" Foods 11, no. 13: 1957. https://doi.org/10.3390/foods11131957
APA StylePaucar-Menacho, L. M., Castillo-Martínez, W. E., Simpalo-Lopez, W. D., Verona-Ruiz, A., Lavado-Cruz, A., Martínez-Villaluenga, C., Peñas, E., Frias, J., & Schmiele, M. (2022). Performance of Thermoplastic Extrusion, Germination, Fermentation, and Hydrolysis Techniques on Phenolic Compounds in Cereals and Pseudocereals. Foods, 11(13), 1957. https://doi.org/10.3390/foods11131957