
Metabolic Profiling of Bulgarian Potato Cultivars



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction, Derivatization of Polar Metabolites and Gas Chromatography Coupled with Mass Spectrometry Analysis
2.3. Statistical Analyses
2.4. Extraction and Spectrophotometric Analysis of Phenolic Compounds
2.5. Extraction and Spectrophotometric Analysis of Total Flavonoid Compounds
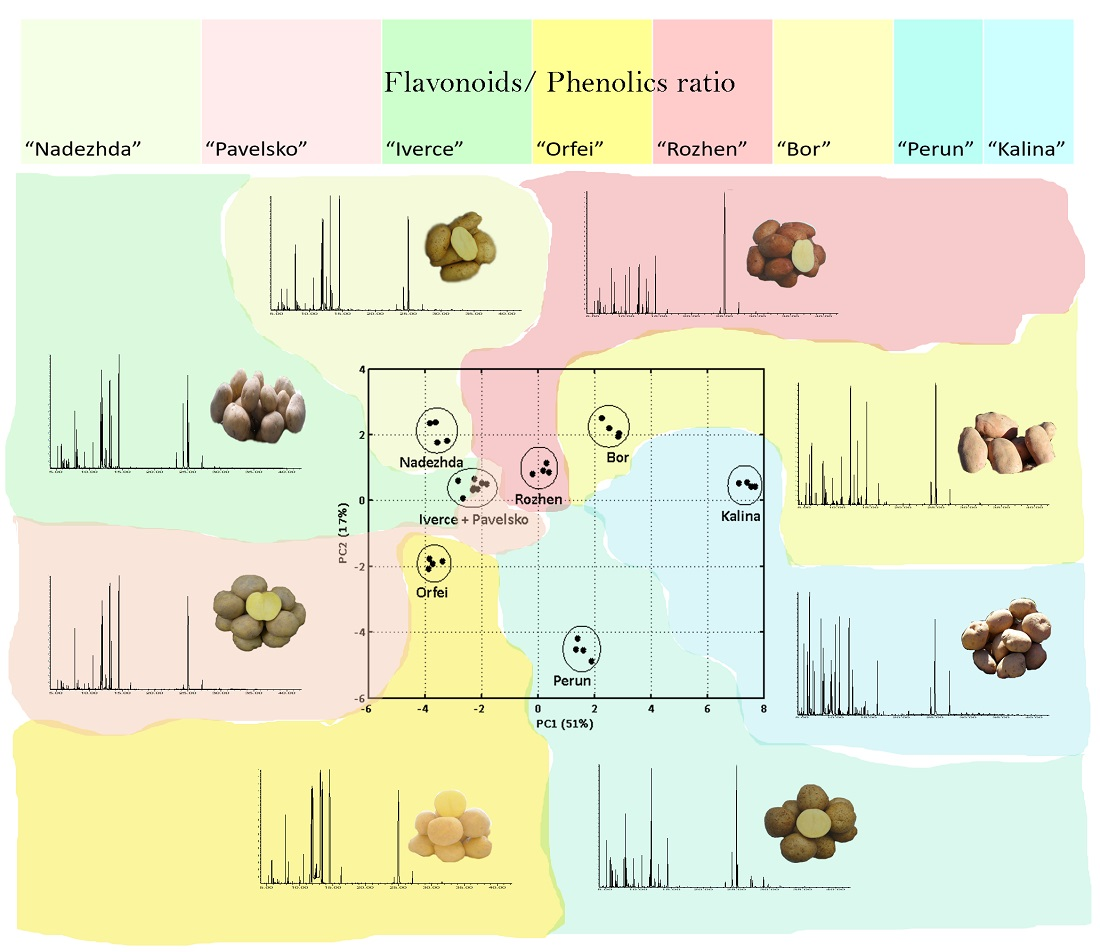
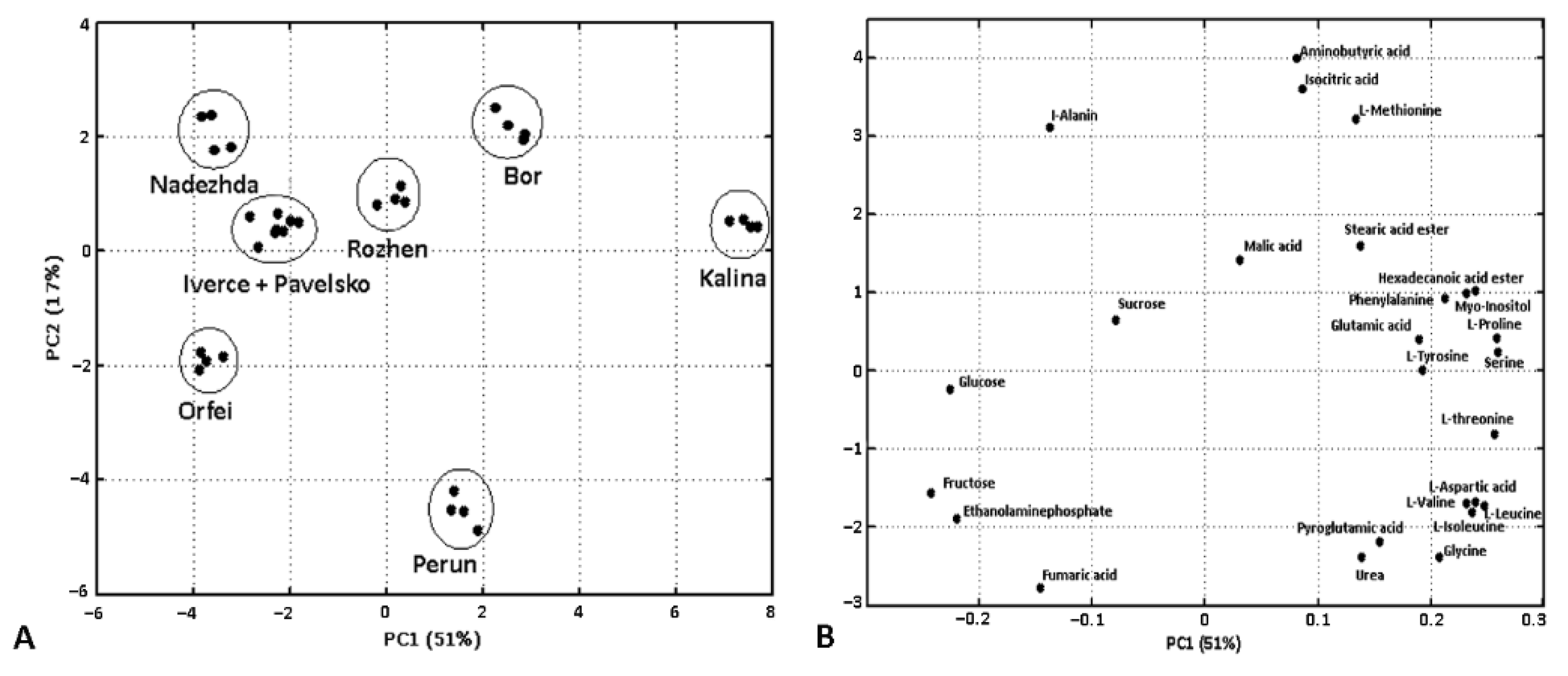
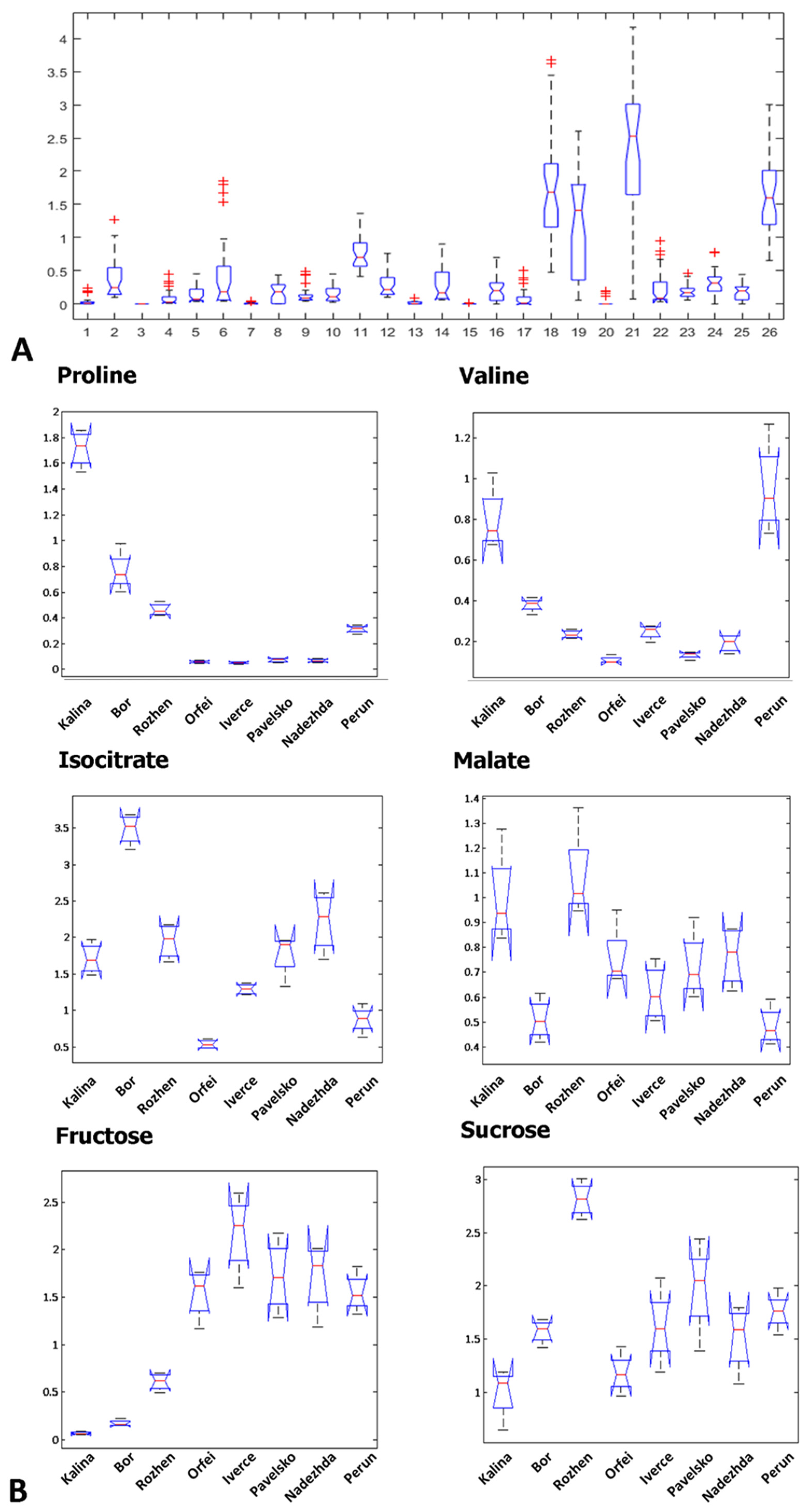
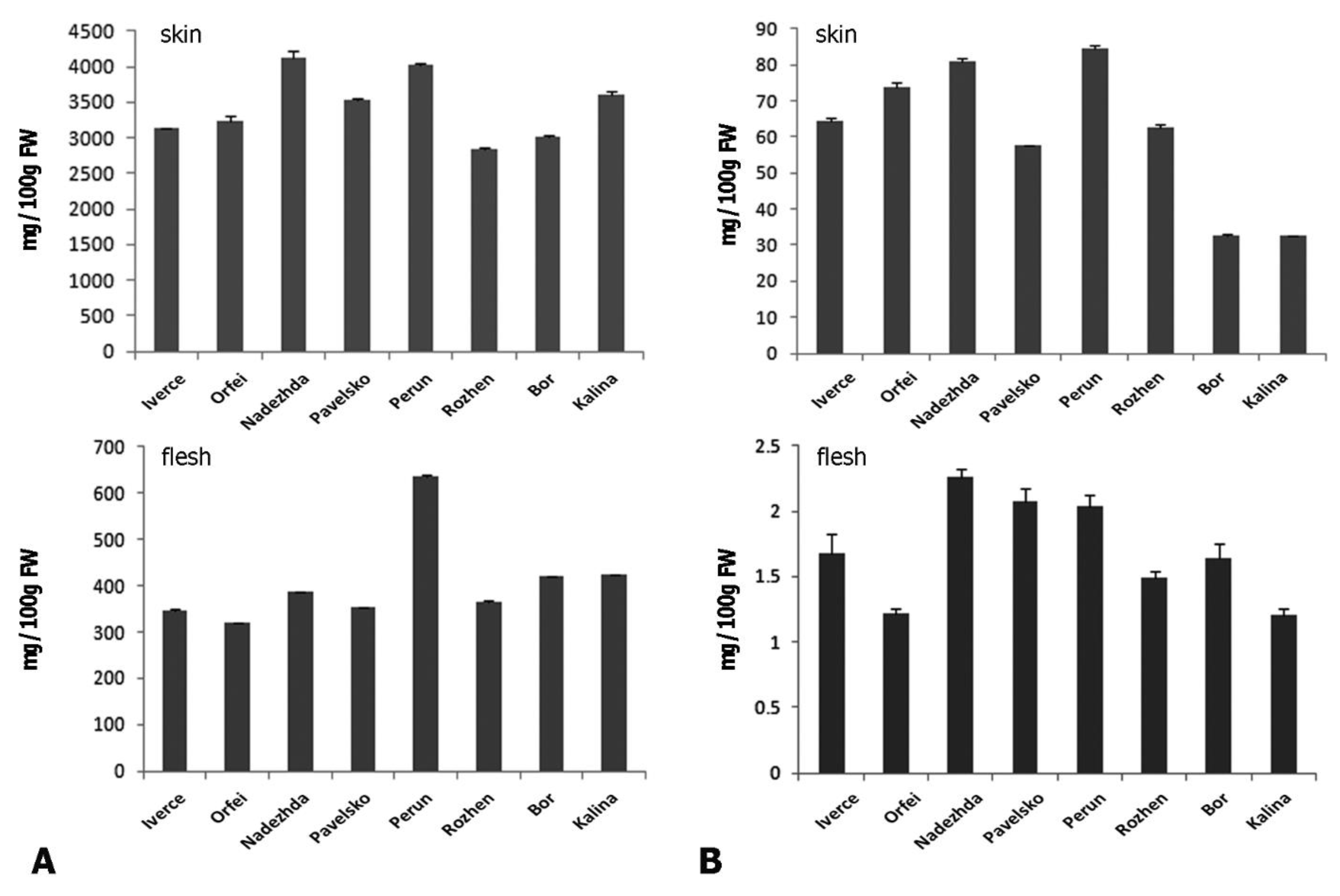
3. Results
3.1. GC-MS Metabolite Profiling
3.2. Spectrophotometric Analysis of Total Flavonoids and Phenolics Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| AV | average value |
| DW | dry weight |
| GC | gas chromatography |
| FW | fresh weight |
| MATLAB | mathematical graphics programming |
| MS | mass chromatography |
| NIST | mass spectral database |
| PC | parent component |
| PCA | principal component analysis |
| PC1, PC2 | principal component 1, 2 |
| TMS | trimethylsilyl |
| TIC | total ion current |
| VCRI | Vegetable Crops Research Institute |
References
- Mahgoub, H.A.M.; Eisa, G.S.A.; Youssef, M.A.H. Molecular, biochemical and anatomical analysis of some potato (Solanum tuberosum L.) cultivars growing in Egypt. J. Genet. Eng. Biotechnol. 2015, 13, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, F.; Wu, Y.; Hu, H.-H.; Dai, X.-F. Progress of potato staple food research and industry development in China. J. Integr. Agric. 2017, 16, 2924–2932. [Google Scholar] [CrossRef]
- FAOSTAT Statistics Division. Potatoes Production in the World. 2020. Available online: http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor (accessed on 16 March 2022).
- Sosa, P.; Guild, G.; Burgos, G.; Bonierbale, M.; Zum Felde, T. Potential and application of X-ray fluorescence spectrometry to estimate iron and zinc concentration in potato tubers. J. Food Compos. Anal. 2018, 70, 22–27. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ghislain, M.; Simon, R.; Jansky, S.H.; Gavrilenko, T. Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot. Rev. 2014, 80, 283–383. [Google Scholar] [CrossRef]
- Bashir, I.; Nicolao, R.; Heiden, G. Wild potatoes: A genetic reservoir for potato breeding. In Wild Germplasm for Genetic Improvement in Crop Plants; Academic Press: Cambridge, MA, USA, 2021; pp. 215–240. [Google Scholar]
- Aversano, R.; Contaldi, F.; Adelfi, M.; D’Amelia, V.; Diretto, G.; de Tommasi, N.; Vaccaro, C.; Vassallo, A.; Carputo, D. Comparative metabolite and genome analysis of tuber-bearing potato species. Phytochemistry 2017, 137, 42–51. [Google Scholar] [CrossRef]
- Drapal, M.; Lindqvist-Kreuze, H.; Mihovilovich, E.; Aponte, M.; Bonierbale, M.; Fraser, P.D. Cooking dependent loss of metabolites in potato breeding lines and their wild and landrace relatives. J. Food Compos. Anal. 2020, 88, 103432. [Google Scholar] [CrossRef]
- Rasheed, H.; Ahmad, D.; Bao, J. Genetic diversity and health properties of polyphenols in potato. Antioxidants 2022, 11, 603. [Google Scholar] [CrossRef]
- Sulli, M.; Mandolino, G.; Sturaro, M.; Onofri, C.; Diretto, G.; Parisi, B.; Giuliano, G. Molecular and biochemical characterization of a potato collection with contrasting tuber carotenoid content. PLoS ONE 2017, 12, e0184143. [Google Scholar] [CrossRef]
- Keita, F.; Karpukhin, M.Y. Development of cultivar technologies for cultivation of potato in the Middle Urals. Agrar. Bull. Urals. 2020, S14, 35–39. [Google Scholar] [CrossRef]
- Ngobese, N.Z.; Workneh, T.S.; Alimi, B.A.; Tesfay, S. Nutrient composition and starch characteristics of eight European potato cultivars cultivated in South Africa. J. Food Compos. Anal. 2017, 55, 1–11. [Google Scholar] [CrossRef]
- Zaheer, K.; Akhtar, M. Potato production, usage, and nutrition. Crit. Rev. Food Sci. Nutr. 2016, 56, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Mu, T.; Ma, M.; Zhang, R.; Sun, Q.; Xu, Y. Nutritional evaluation of different cultivars of potatoes (Solanum tuberosum L.) from China by grey relational analysis (GRA) and its application in potato steamed bread making. J. Integr. Agric. 2019, 18, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Londhe, S. Sustainable Potato Production and the Impact of Climate Change; IGI Global: Hershey, PA, USA, 2016; 323p. [Google Scholar]
- Parameswaran, R.; Bhuvaneswar, P. Detection of macro and micro nutrients in potatoes using elemental analysis techniques. Int. J. Recent Technol. Eng. IJRTE 2020, 8, 1033–1040. [Google Scholar] [CrossRef]
- Akyol, H.; Riciputi, Y.; Capanoglu, E.; Caboni, M.F.; Verardo, V. Phenolic compounds in the potato and its byproducts: An Overview. Int. J. Mol. Sci. 2016, 17, 835. [Google Scholar] [CrossRef]
- Habeebullah, S.F.K.; Grejsen, H.D.; Jacobsen, C. Potato peel extract as a natural antioxidant in chilled storage of minced horse mackerel (Trachurus trachurus): Effect on lipid and protein oxidation. Food Chem. 2012, 131, 843–851. [Google Scholar]
- Mohdaly, A.; Sarhan, M.; Mahmoud, A.; Ramadan, M.; Smetanska, I. Antioxidant efficacy of potato peels and sugar beet pulp extracts in vegetable oils protection. Food Chem. 2010, 123, 1019–1026. [Google Scholar] [CrossRef]
- Mäder, J.; Rawel, H.; Kroh, L. Composition of phenolic compounds and glycoalkaloids α-solanine and α-chaconine during commercial potato processing. J. Agric. Food Chem. 2009, 57, 6292–6297. [Google Scholar] [CrossRef]
- Mattila, P.; Hellström, J. Phenolic acids in potatoes, vegetables, and some of their products. J. Food Comp. Anal. 2007, 20, 152–160. [Google Scholar] [CrossRef]
- Deußer, H.; Guignard, C.; Hoffmann, L.; Evers, D. Polyphenol and glycoalkaloid contents in potato cultivars grown in Luxembourg. Food Chem. 2012, 135, 2814–2824. [Google Scholar] [CrossRef]
- Uri, C.; Juhász, Z.; Polgár, Z.; Bánfalvi, Z. A GC-MS-based metabolomics study on the tubers of commercial potato cultivars upon storage. Food Chem. 2014, 159, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Dobson, G.; Shepherd, T.; Verrall, S.R.; Griffiths, W.D.; Ramsay, G.; McNicol, J.W.; Davies, H.V.; Stewart, D.A. Metabolomics study of cultivated potato (Solanum tuberosum) groups andigena, phureja, stenotomum, and tuberosum using gas chromatography−mass spectrometry. J. Agric. Food Chem. 2010, 58, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Pęksa, A.; Miedzianka, J.; Nemś, A.; Rytel, E. The free-amino-acid content in six potatoes cultivars through storage. Molecules 2021, 26, 1322. [Google Scholar] [CrossRef] [PubMed]
- Nacheva, E. Achievements and trends in potato breeding in Maritsa Vegetable Crops Research Institute Plovdiv during the new millennium. Acta Hortic. 2009, 830, 121–126. [Google Scholar] [CrossRef]
- Nacheva, E.; Blagoeva, V. Bulgarian potato cultivars. J. Agric. Plus 2010, 4, 10–11. (In Bulgarian) [Google Scholar]
- Nacheva, E. Rozhen—A new Bulgarian potato cultivar suitable for fresh consumption and processing. In Proceedings of the EAPR Agronomy Section Meeting, Mamaia, Romania, 23–27 June 2004; pp. 57–65. [Google Scholar]
- Nacheva, E. Peroun—A new Bulgarian potato cultivar suitable for fresh consumption and processing. In Proceedings of the Stara Zagora Union of Scientists, Scientific Conference with International Participation, Stara Zagora, Bulgaria, 3–4 June 2004; Volume 2, pp. 178–183. (In Bulgarian). [Google Scholar]
- Nacheva, E.; Petkova, V.; Pevicharova, G.; Samaliev, H. Orfei—A new Bulgarian potato cultivar resistant to potato cyst nematodes Globodera rostochiensis and Globodera pallida. In Proceedings of the XIV International Scientific and Practical Conference “Agricultural Science—Agricultural Production in Siberia, Mongolia, Kazakhstan and Bulgaria”, Krasnoyarsk, Russia, 25–28 July 2011; pp. 128–131. [Google Scholar]
- Nacheva, E.; Petkova, V.; Pevicharova, G.; Samaliev, H. Pavelsko—A new Bulgarian potato cultivar suitable for chips. Turk. J. Agric. Nat. Sci. 2014, 1, 1554–1558. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- EN ISO/IEC 17025:2001; Bulgarian State Standard Method. Bulgarian Institute for Standardization: Sofia, Bulgaria, 2001.
- Wichrowska, D.; Rogozińska, I.; Pawelzik, E. Concentrations of some organic acids in potato tubers depending on weed control method, cultivar and storage conditions. Pol. J. Environ. Stud. 2009, 18, 487–491. [Google Scholar]
- Liu, P.; Xu, N.; Liu, R.; Liu, J.; Peng, Y.; Wang, Q. Exogenous proline treatment inhibiting enzymatic browning of fresh-cut potatoes during cold storage. Postharvest Biol. Technol. 2022, 184, 111754. [Google Scholar] [CrossRef]
- Burton, W.G.; van Es, A.; Hartmans, K.S. The physics and physiology of storage. In The Potato Crop, 2nd ed.; Harris, P.M., Ed.; Chapman and Hall: London, UK, 1992; pp. 608–727. [Google Scholar]
- Duarte-Delgado, D.; Ñústez-López, C.E.; Narváez-Cuenca, C.E.; Restrepo-Sánchez, L.P.; Melo, S.E.; Sarmiento, F.; Kushalappa, A.C.; Mosquera-Vásquez, T. Natural variation of sucrose, glucose and fructose contents in Colombian genotypes of Solanum tuberosum group Phureja at harvest. J. Sci. Food Agric. 2016, 96, 4288–4294. [Google Scholar] [CrossRef] [Green Version]
- Andre, C.M.; Ghislain, M.; Bertin, P.; Oufir, M.; del Rosario Herrera, M.; Hoffmann, L.; Hausman, J.F.; Larondelle, Y.; Evers, D. Andean potato cultivars (Solanum tuberosum L.) as a source of antioxidant and mineral micronutrients. J. Agric. Food Chem. 2007, 55, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Kanatt, S.; Chander, R.; Radhakrishna, P.; Sharma, A. Potato peel extracts natural antioxidant for retarding lipid peroxidation in radiation processed lamb meat. J. Agric. Food Chem. 2005, 53, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Volnová, B.; Musilová, J.; Hrabovská, D.; Bystrická, J.; Kavalcová, P.; Kopernická, M. The research of total polyphenols content and their changes in different cultivars of potatoes. J. Microbiol. Biotechnol. Food Sci. 2015, 3, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.R. Antioxidants in potato. Am. J. Potato Res. 2005, 82, 163–172. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potato Cultivar (Analysed Part of Tuber) | Phenolics (mg/100 g FW) | Flavonoids (mg/100 g FW) | Flavonoids/Phenolics Ratio |
|---|---|---|---|
| “Iverce” (skin) | 3130.09 ± 11.9 | 64.61 ± 0.68 | 0.021 |
| “Iverce” (flesh) | 346.24 ± 1.7 | 1.68 ± 0.15 | 0.005 |
| “Orfei” (skin) | 3246.42 ± 70.68 | 74.03 ± 1.01 | 0.023 |
| “Orfei” (flesh) | 318.12 ± 0.47 | 1.22 ± 0.04 | 0.004 |
| “Nadezhda” (skin) | 4120.06 ± 10.48 | 80.93 ± 1.05 | 0.020 |
| “Nadezhda” (flesh) | 385.82 ± 1.83 | 2.26 ± 0.06 | 0.006 |
| “Pavelsko” (skin) | 3522.44 ± 27.07 | 57.51 ± 0.41 | 0.016 |
| “Pavelsko” (flesh) | 352.19 ± 2.04 | 2.07 ± 0.1 | 0.006 |
| “Perun” (skin) | 4015.26 ± 46.8 | 84.38 ± 0.86 | 0.021 |
| “Perun” (flesh) | 636.5 ± 1.52 | 2.04 ± 0.09 | 0.003 |
| “Rozhen” (skin) | 2846.89 ± 15.91 | 62.51 ± 1.09 | 0.022 |
| “Rozhen” (flesh) | 363.78 ± 3.65 | 1.49 ± 0.05 | 0.004 |
| “Bor” (skin) | 3004.95 ± 29.57 | 32.69 ± 0.39 | 0.011 |
| “Bor” (flesh) | 420.7 ± 0.97 | 1.64 ± 0.11 | 0.004 |
| “Kalina” (skin) | 3597.55 ± 57.56 | 32.54 ± 0.31 | 0.009 |
| “Kalina” (flesh) | 423.86 ± 1.54 | 1.21 ± 0.05 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomlekova, N.; Mladenov, P.; Dincheva, I.; Nacheva, E. Metabolic Profiling of Bulgarian Potato Cultivars. Foods 2022, 11, 1981. https://doi.org/10.3390/foods11131981
Tomlekova N, Mladenov P, Dincheva I, Nacheva E. Metabolic Profiling of Bulgarian Potato Cultivars. Foods. 2022; 11(13):1981. https://doi.org/10.3390/foods11131981
Chicago/Turabian StyleTomlekova, Nasya, Petko Mladenov, Ivayla Dincheva, and Emilya Nacheva. 2022. "Metabolic Profiling of Bulgarian Potato Cultivars" Foods 11, no. 13: 1981. https://doi.org/10.3390/foods11131981
APA StyleTomlekova, N., Mladenov, P., Dincheva, I., & Nacheva, E. (2022). Metabolic Profiling of Bulgarian Potato Cultivars. Foods, 11(13), 1981. https://doi.org/10.3390/foods11131981