
Impact of Protein Content on the Antioxidants, Anti-Inflammatory Properties and Glycemic Index of Wheat and Wheat Bran

 , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Materials
2.3. Proximate Composition
2.4. Phenolic Extracts Preparation
2.4.1. Release of Free Phenolic Compounds (FP)
2.4.2. Release of Bound Phenolic Compounds (BP)
2.5. Determination of Total Phenolics Compounds (TP)
2.6. Characterization of Phenolic Compounds by HPLC-ESI-QTOF-MS
2.7. Total Antioxidant Capacity (TAC)
2.7.1. DPPH Radical Scavenging Activity
2.7.2. ABTS•+•+ Radical Cation Scavenging Activity
2.7.3. Oxygen Radical Absorbance Capacity (ORAC)
2.7.4. Ferric Reducing Antioxidant Power (FRAP)
2.8. Glycemic Index (GI)
2.9. Determination of Anti-Inflammatory Activity (AIA)
2.10. Statistical Analysis
3. Results and Discussion
3.1. Proximal Composition
3.2. Determination of Total Phenolic Compounds (TP)
3.3. Characterization of Phenolic Compounds by HPLC-ESI-QTOF-MS
3.4. Total Antioxidant Capacity (TAC)
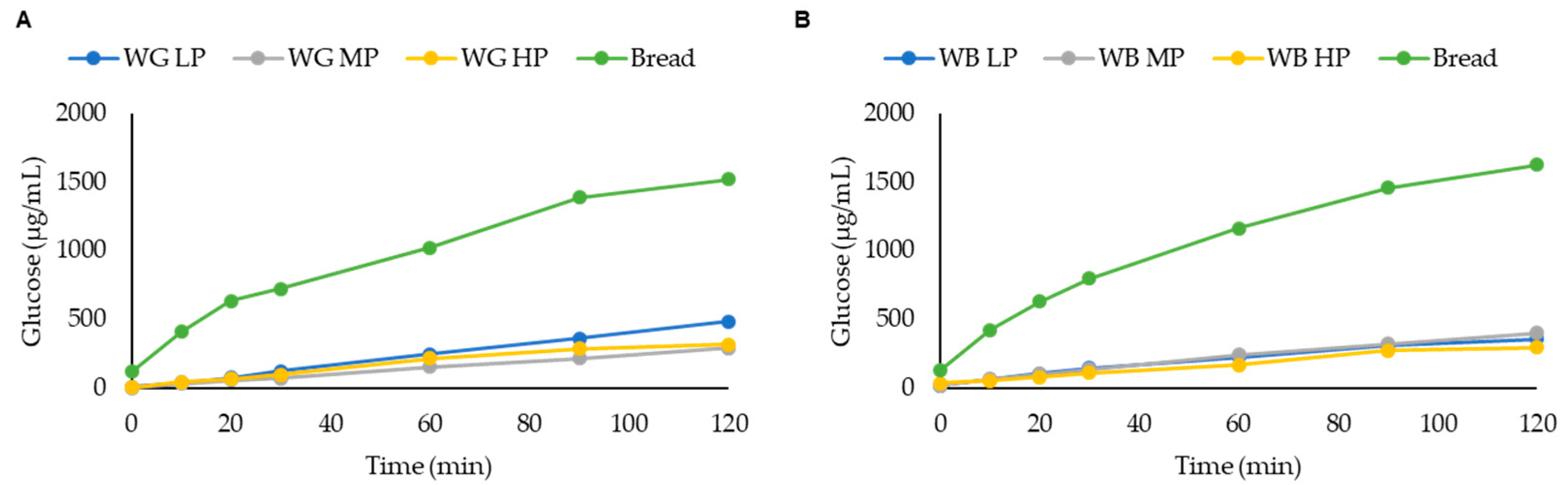
3.5. Glycemic Index (GI)
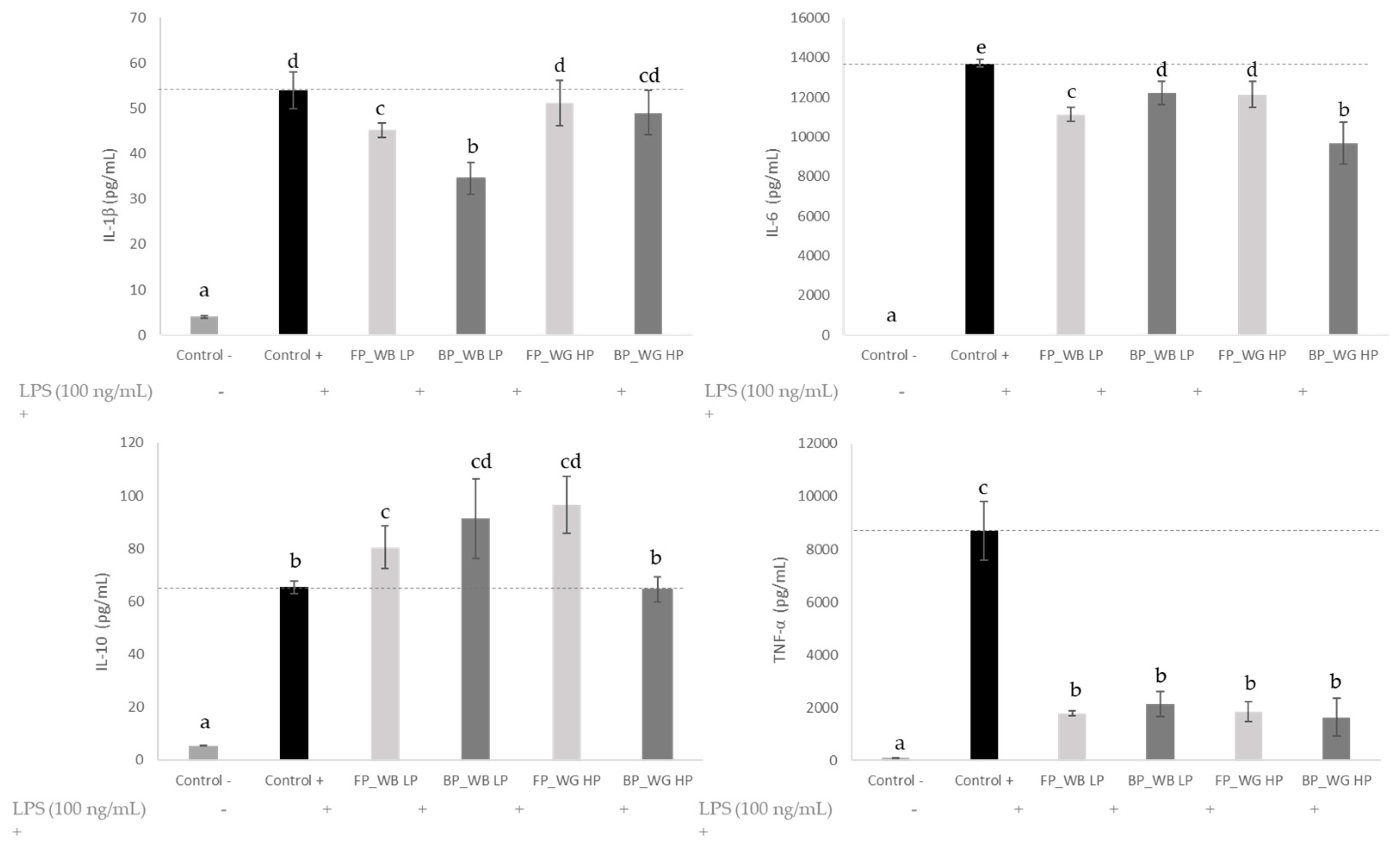
3.6. Anti-Inflammatory Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- EC. Circular Economy Action Plan. Available online: https://ec.europa.eu/environment/strategy/circular-economy-action-plan_en (accessed on 6 March 2022).
- Pandey, M.; Shrestha, J.; Subedi, S.; Shah, K.K. Role of nutrients in wheat: A review. Trop. Agrobiodiversity 2020, 1, 18–23. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- FAO. Cereal Supply and Demand Data May 2020. Available online: http://www.fao.org/fileadmin/templates/worldfood/Reports_and_docs/Cereal_supply_and_demand_data_may.xls (accessed on 4 March 2022).
- Rosa, N.N.; Dufour, C.; Lullien-Pellerin, V.; Micard, V. Exposure or release of ferulic acid from wheat aleurone: Impact on its antioxidant capacity. Food Chem. 2013, 141, 2355–2362. [Google Scholar] [CrossRef]
- Apprich, S.; Tirpanalan, Ö.; Hell, J.; Reisinger, M.; Böhmdorfer, S.; Siebenhandl-Ehn, S.; Novalin, S.; Kneifel, W. Wheat branbased biorefinery 2: Valorization of products. LWT Food Sci. Technol. 2014, 56, 222–231. [Google Scholar] [CrossRef]
- Andersson, A.A.; Dimberg, L.; Åman, P.; Landberg, R. Recent findings on certain bioactive components in whole grain wheat and rye. J. Cereal Sci. 2014, 59, 294–311. [Google Scholar] [CrossRef]
- Rico, D.; Villaverde, A.; Martinez-Villaluenga, C.; Gutierrez, A.L.; Caballero, P.A.; Ronda, F.; Peñas, E.; Frias, J.; Diana, A.B.M. Application of Autoclave Treatment for Development of a Natural Wheat Bran Antioxidant Ingredient. Foods 2020, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-W.; Hsu, C.-P.; Yang, B.B.; Wang, C.-Y. Advances in the extraction of natural ingredients by high pressure extraction technology. Trends Food Sci. Technol. 2013, 33, 54–62. [Google Scholar] [CrossRef]
- Bautista-Expósito, S.; Tomé-Sánchez, I.; Martín-Diana, A.B.; Frias, J.; Peñas, E.; Rico, D.; Casas, M.J.G.; Martínez-Villaluenga, C. Enzyme Selection and Hydrolysis under Optimal Conditions Improved Phenolic Acid Solubility, and Antioxidant and AntiInflammatory Activities of Wheat Bran. Antioxidants 2020, 9, 984. [Google Scholar] [CrossRef]
- Kaur, A.; Yadav, M.P.; Singh, B.; Bhinder, S.; Simon, S.; Singh, N. Isolation and characterization of arabinoxylans from wheat bran and study of their contribution to wheat flour dough rheology. Carbohydr. Polym. 2019, 221, 166–173. [Google Scholar] [CrossRef]
- Hemery, Y.; Xavier Rouau, X.; Lullien-Pellerin, V.; Barron, J.C.; Abecassis, J. Dry processes to develop wheat fractions and products with enhanced nutritional quality. J. Cereal Sci. 2007, 46, 327–347. [Google Scholar] [CrossRef]
- Anderson, J.W.; Baird, P.; Davis, R.H., Jr.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health benefits of dietary fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.L. Dietary fiber and body weight. Nutrition 2005, 21, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, D.W.; Kruger, J.E. Simple phenolic acids in flours prepared from Canadian wheat: Relationship to ash content, color and polyphenol oxidase activity. Cereal Chem. 1997, 74, 337–343. [Google Scholar] [CrossRef]
- Mhd Omar, N.A.; Wu, H.; Larsson, A.; Wolk, A.; Landberg, R. Long-Term Whole-Grain Rye and Wheat Consumption and Their Associations with Selected Biomarkers of Inflammation, Endothelial Function, and Cardiovascular Disease. Eur. J. Clin. Nutr. 2021, 75, 123–132. [Google Scholar] [CrossRef]
- Calabriso, N.; Massaro, M.; Scoditti, E.; Pasqualone, A.; Laddomada, B.; Carluccio, M.A. Phenolic Extracts from Whole Wheat Biofortified Bread Dampen Overwhelming Inflammatory Response in Human Endothelial Cells and Monocytes: Major Role of VCAM-1 and CXCL-10. Eur. J. Nutr. 2020, 59, 2603–2615. [Google Scholar] [CrossRef]
- Zhu, Y.; Sang, S. Phytochemicals in Whole Grain Wheat and Their Health-promoting Effects. Mol. Nutr. Food Res. 2017, 61, 1600852. [Google Scholar] [CrossRef]
- Martín-Diana, A.B.; García-Casas, M.J.; Martínez-Villaluenga, C.; Frías, J.; Peñas, E.; Rico, D. Wheat and Oat Brans as Sources of Polyphenol Compounds for Development of Antioxidant Nutraceutical Ingredients. Foods 2021, 10, 115. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Saura-Calixto, F. Literature Data May Underestimate the Actual Antioxidant Capacity of Cereals. J. Agric. Food Chem. 2005, 53, 5036–5040. [Google Scholar] [CrossRef] [Green Version]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Bautista-Expósito, S.; Frias, J.; Rico, D.; González-Maillo, L.; Martinez-Villaluenga, C. Soluble Phenolic Composition Tailored by Germination Conditions Accompany Antioxidant and Anti-Inflammatory Properties of Wheat. Antioxidants 2020, 9, 426. [Google Scholar] [CrossRef]
- Martín-Diana, A.B.; Tomé-Sánchez, I.; García-Casas, M.J.; Martínez-Villaluenga, C.; Frías, J.; Rico, D. A Novel Strategy to Produce a Soluble and Bioactive Wheat Bran Ingredient Rich in Ferulic Acid. Antioxidants 2021, 10, 969. [Google Scholar] [CrossRef] [PubMed]
- Marconi, E.; Carcea, M. Pasta from non-traditional raw materials. Cereal Foods World 2001, 46, 522–530. [Google Scholar]
- Cros, D.L. Glutenin proteins and gluten strength in durum wheat. J. Cereal Sci. 1987, 5, 3–12. [Google Scholar] [CrossRef]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.A.; Beom Park, C.; Tsuda, H. Soluble and insoluble fiber influences on cancer development. Crit. Rev. Oncol. Hematol. 1998, 27, 229–242. [Google Scholar] [CrossRef]
- Moore, J.; Hao, Z.; Zhou, K.; Luther, M.; Costa, J.; Yu, L. Carotenoid, tocopherol, phenolic acid, and antioxidant properties of Maryland-grown soft wheat. J. Agric. Food Chem. 2005, 53, 6649–6657. [Google Scholar] [CrossRef]
- Yu, L.; Haley, S.; Perret, J.; Harris, M. Comparison of wheat flours grown at different locations for their antioxidant properties. Food Chem. 2004, 86, 11–16. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; AOAC: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Dinelli, G.; Segura-Carretero, A.; Di Silvestro, R.; Marotti, I.; Arráez-Román, D.; Benedettelli, S.; Ghiselli, L.; Fernadez-Gutierrez, A. Profiles of phenolic compounds in modern and old common wheat varieties determined by liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 7670–7681. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total Phenol Analysis: Automation and Comparison with Manual Methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS•+•+ radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Martin-Diana, A.B.; Izquierdo, N.; Albertos, I.; Sanchez, M.S.; Herrero, A.; Sanz, M.A.; Rico, D. Valorization of Carob’s Germ and Seed Peel as Natural Antioxidant Ingredients in Gluten-Free Crackers. J. Food Process. Preserv. 2016, 41, e12770. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Gularte, M.; Rosell, C. Physicochemical properties and enzymatic hydrolysis of different starches in the presence of hydrocolloids. Carbohydr. Polym. 2011, 85, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Granfeldt, Y. Foods Factors Affecting Metabolic Responses to Cereal Products. Ph.D. Thesis, University of Lund, Lund, Sweden, 1994. [Google Scholar]
- Chalamacharla, R.B.; Harsha, K.; Sheik, K.B.; Viswanatha, C.K. Wheat Bran-Composition and Nutritional Quality: A Review. Adv. Biotech. Micro. 2018, 9, 555754. [Google Scholar] [CrossRef]
- Brouns, F.; Hemery, Y.; Price, R.; Anson, N.M. Wheat aleurone: Separation, composition, health aspects, and potential food use. Crit. Rev. Food Sci. Nutr. 2012, 52, 553–568. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Lafiandra, D.; Tamás, L.; Békés, F. Genetic Manipulation of Gluten Structure and Function. In Gliadin and Glutenin: The Unique Balance of Wheat Quality; Bekes, F., Bushuk, W., Wrigley, C., Eds.; American Association of Cereal Chemists (AACC): St Paul, MN, USA, 2006; pp. 363–385. [Google Scholar]
- Cheryan, M. Phytic acid interactions in food systems. Crit. Rev. Food Sci. Nutr. 1980, 13, 297–335. [Google Scholar] [CrossRef]
- Rickard, E.S.; Thomson, L.U. Interactions and effects of phytic acid. In Antinutrients and Phytochemicals in Foods; Shahidi, F., Ed.; American Chemical Society: Washington, DC, USA, 1997; pp. 294–312. [Google Scholar]
- Yoon, J.H.; Thompson, L.U.; Jenkins, D.J. The effect of phytic acid on in vitro rate of starch digestibility and blood glucose response. Am. J. Clin. Nutr. 1983, 38, 835–842. [Google Scholar] [CrossRef]
- Katayama, T. Effect of dietary sodium phytate on the hepatic and serum levels of lipids and on the hepatic activities of NADPH-generating enzymes in rats fed on sucrose. Biosci. Biotechnol. Biochem. 1995, 59, 1159–1160. [Google Scholar] [CrossRef]
- Minihane, A.M.; Rimbach, G. Iron absorption and the iron binding and antioxidant properties of phytic acid. Int. J. Food Sci. Technol. 2002, 37, 741–748. [Google Scholar] [CrossRef]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Frias, J.; Rico, D.; Jiménez-Pulido, I.; Martínez-Villaluenga, C. Bioprocessed Wheat Ingredients: Characterization, Bioaccessibility of Phenolic Compounds, and Bioactivity During in vitro Digestion. Front. Plant Sci. 2021, 12, 790898. [Google Scholar] [CrossRef] [PubMed]
- Liyana-Pathirana, C.M.; Shahidi, F. Importance of Insoluble-Bound Phenolics to Antioxidant Properties of Wheat. J. Agr. Food Chem. 2006, 54, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Nanguet, A.L.; Beta, T. Comparison of Antioxidant Properties of Refined and Whole Wheat Flour and Bread. Antioxidants 2013, 2, 370–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harnly, J.M.; Bhagwat, S.; Lin, L.Z. Profiling Methods for the Determination of Phenolic Compounds in Foods and Dietary Supplements. Anal. Bioanal. Chem. 2007, 389, 47–61. [Google Scholar] [CrossRef] [Green Version]
- De Brier, N.; Gomand, S.V.; Celus, I.; Courtin, C.M.; Brijs, K.; Delcour, J.A. Extractability and chromatographic characterization of wheat (Triticum aestivum L.) bran protein. J. Food Sci. 2015, 80, C967–C974. [Google Scholar] [CrossRef]
- Idris, W.H.; Babiker, E.E.; El Tinay, A.H. Fractionation, solubility and functional properties of wheat bran proteins as influenced by pH and/or salt concentration. Nahrung/Food 2003, 47, 425–429. [Google Scholar] [CrossRef]
- Sęczyk, Ł.; Świeca, M.; Kapusta, I.; Gawlik-Dziki, U. Protein-Phenolic Interactions as a Factor Affecting the Physicochemical Properties of White Bean Proteins. Molecules 2019, 24, 408. [Google Scholar] [CrossRef] [Green Version]
- Gammoh, S.; Alu’datt, M.H.; Alhamad, M.N.; Rababah, T.; Al-Mahasneh, M.; Qasaimeh, A.; Johargy, A.; Kubow, S.; Hussein, N.M. The effects of protein-phenolic interactions in wheat protein fractions on allergenicity, antioxidant activity and the inhibitory activity of angiotensin I-converting enzyme (ACE). Food Biosci. 2018, 24, 50–55. [Google Scholar] [CrossRef]
- Gammoh, S.; Alu’datt, M.H.; Alhamad, M.N.; Rababah, T.; Ereifej, K.; Almajwal, A.; Ammari, Z.A.; Al Khateeb, W.; Hussein, N.M. Characterization of phenolic compounds extracted from wheat protein fractions using high-performance liquid chromatography/liquid chromatography mass spectrometry in relation to anti-allergenic, anti-oxidant, anti-hypertension, and anti-diabetic properties. Int. J. Food Prop. 2017, 20, 2383–2395. [Google Scholar] [CrossRef] [Green Version]
- Justesen, U. Collision-induced fragmentation of deprotonated methoxylated flavonoids, obtained by electrospray ionization mass spectrometry. J. Mass Spectrom. 2001, 36, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Gruz, J.; Novák, O.; Miroslav Strnad, M. Rapid analysis of phenolic acids in beverages by UPLC–MS/MS. Food Chem. 2008, 111, 789–794. [Google Scholar] [CrossRef]
- Mattila, P.; Pihlava, J.M.; Hellström, J. Contents of phenolic acids, alkyl- and alkenylresorcinols, and avenanthramides in commercial grain products. J. Agric. Food Chem. 2005, 53, 8290–8295. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Chiodelli, G.; Giuberti, G.; Gallo, A.; Masoero, F.; Trevisan, M. Phenolic profile and fermentation patterns of different commercial gluten-free pasta during in vitro large intestine fermentation. Food Res. Int. 2017, 97, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal dietary fiber: A natural functional ingredient to deliver phenolic compounds into the gut. Trends Food Sci. Technol. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Awika, J.M.; Rose, D.J.; Simsek, S. Complementary effects of cereal and pulse polyphenols and dietary fiber on chronic inflammation and gut health. Food Funct. 2018, 9, 1389–1409. [Google Scholar] [CrossRef]
- Martín-García, B.; Gómez-Caravaca, A.M.; Marconi, E.; Verardo, V. Distribution of free and bound phenolic compounds, and alkylresorcinols in wheat aleurone enriched fractions. Food Res. Int. 2021, 140, 109816. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef] [Green Version]
- Siddeeg, A.; AlKehayez, N.M.; Abu-Hiamed, H.A.; Al-Sanea, E.A.; AL-Farga, A.M. Mode of action and determination of antioxidant activity in the dietary sources: An overview. Saudi J. Biol. Sci. 2021, 28, 1633–1644. [Google Scholar] [CrossRef]
- Sevgi, K.; Tepe, B.; Sarikurkcu, C. Antioxidant and DNA damage protection potentials of selected phenolic acids. Food Chem. Toxicol. 2015, 77, 12–21. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh, A.; Ghasemzadeh, N. Flavonoids and phenolic acids: Role and biochemical activity in plants and human. J. Med. Plants Res. 2011, 5, 6697–6703. [Google Scholar] [CrossRef]
- Adom, K.K.; Sorrells, M.; Liu, R. Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [Google Scholar] [CrossRef]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic acids in wheat varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef] [PubMed]
- Przybylski, P.; Konopko, A.; Łętowski, P.; Jodko-Piórecka, K.; Litwinienko, G. Concentration-dependent HAT/ET mechanism of the reaction of phenols with 2,2-diphenyl-1-picrylhydrazyl (dpph) in methanol. RSC Adv. 2022, 12, 8131–8136. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Giuberti, G.; Bhumireddy, S.R.; Mandal, R.; Trevisan, M.; Wishart, D.S. Transformation of polyphenols found in pigmented gluten-free flours during in vitro large intestinal fermentation. Food Chem. 2019, 298, 125068. [Google Scholar] [CrossRef]
- Chung, H.J.; Lim, H.S.; Lim, S.T. Effect of partial gelatinization and retrogradation on the enzymatic digestion of waxy rice starch. J. Cereal Sci. 2006, 43, 353–359. [Google Scholar] [CrossRef]
- Brennan, C.S.; Blake, D.E.; Ellis, P.R.; Schofield, J.D. Effects of Guar Galactomannan on Wheat Bread Microstructure and on the In vitro and In vivo Digestibility of Starch in Bread. J. Cereal Sci. 1996, 24, 151–160. [Google Scholar] [CrossRef]
- Thompson, L.U. Antinutrients and blood glucose. Food Technol. 1988, 42, 123–132. [Google Scholar]
- Gabriele, M.; Pucci, L.; Árvay, J.; Longo, V. Anti-inflammatory and antioxidant effect of fermented whole wheat on TNFα-stimulated HT-29 and NF-κB signaling pathway activation. J. Funct. Foods 2018, 45, 392–400. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, X.; Qiang, S.; Su, J.; Li, J. Anti-Oxidation and Anti-Inflammatory Potency Evaluation of Ferulic Acid Derivatives Obtained through Virtual Screening. Int. J. Mol. Sci. 2021, 22, 11305. [Google Scholar] [CrossRef] [PubMed]
- El-Gogary, R.I.; Nasr, M.; Rahsed, L.A.; Hamzawy, M.A. Ferulic acid nanocapsules as a promising treatment modality for colorectal cancer: Preparation and in vitro/in vivo appraisal. Life Sci. 2022, 298, 120500. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WG LP | WG MP | WG HP | WB LP | WB MP | WB HP | |
|---|---|---|---|---|---|---|
| Ash | 1.79 ± 0.09 a | 1.81 ± 0.19 ab | 2.03 ± 0.23 b | 6.32 ± 0.17 c | 6.26 ± 0.08 c | 7.16 ± 0.30 d |
| TDF | 15.46 ± 0.91 a | 14.41 ± 0.79 a | 14.64 ± 0.06 a | 50.88 ± 19.90 b | 45.24 ± 13.93 b | 46.04 ± 17.86b |
| Fat | 1.73 ± 0.01 a | 1.90 ± 0.40 a | 1.80 ± 0.33 a | 3.87 ± 0.06 b | 3.86 ± 0.20 b | 4.17 ± 0.28 b |
| Moisture | 10.13 ± 0.13b | 10.34 ± 0.16 b | 9.28 ± 0.18 a | 12.59 ± 0.93 c | 12.58 ± 0.61 c | 12.62 ± 1.04 c |
| Proteins | 10.75 ± 0.46 a | 11.82 ± 0.56 b | 17.95 ± 0.57 d | 12.04 ± 0.58 b | 15.16 ± 0.40 c | 19.31 ± 0.28 e |
| Carbohydrates | 85.73 ± 0.38 f | 84.47 ± 0.03 e | 78.21 ± 0.01 d | 77.73 ± 0.35 c | 74.73 ± 0.12 b | 69.36 ± 0.30 a |
| PA | 0.75 ± 0.00 b | 0.70 ± 0.01 a | 0.83 ± 0.02 c | 3.29 ± 0.03 d | 3.55 ± 0.02 e | 3.74 ± 0.00 f |
| TSC | 50.24 ± 2.00 d | 48.30 ± 1.69 c | 54.37 ± 1.22 e | 8.77 ± 0.62 a | 11.56 ± 0.83 b | 6.92 ± 0.30 a |
| Class | Sub-Class | Compound | Molecular Formula | Ion Fragments | WB | WG | ||
|---|---|---|---|---|---|---|---|---|
| Error | Fraction | Error | Fraction | |||||
| Phenolic acids | Hydroxybenzoic acids | Protocatechuic acid | C7H6O4 | 151. 136. 112 | −7.58 | BP | −4.34 | BP |
| Hydroxybenzoic acid | C7H6O3 | 135. 125. 121 | −7.12 | BP | −4.22 | BP | ||
| Hydroxycinnamic acids | Ferulic acid | C10H10O4 | 178. 149. 134 | −3.96 | FP-BP | 0.17 | BP | |
| p-Coumaric acid | C9H8O3 | 119 | 8.95 | BP | n.d. | n.d. | ||
| Sinapic acid | C11H12O5 | 208 | n.d. | n.d. | −1.8 | BP | ||
| Diferulic isomer 1 | C20H18O8 | 341. (282). 193. (112) | −1.84 | BP | −2.1 | BP | ||
| Diferulic isomer 2 | C20H18O8 | 359. 341. 326 | −1.58 | BP | −2.35 | BP | ||
| Diferulic isomer 3 | C20H18O8 | 369. 355. 341. 313. 271 | −3.39 | BP | 1.01 | BP | ||
| Diferulic isomer 4 | C20H18O8 | 341. 326. 282. 248. 227 | 4.38 | BP | −2.87 | BP | ||
| Diferulic isomer 5 | C20H18O8 | 347. 313. 261. 217. 193. 178 | 0.49 | BP | −2.35 | BP | ||
| Diferulic isomer 6 | C20H18O8 | 341. 303. 239. 193. 178 | −1.32 | BP | n.d. | n.d. | ||
| Caffeic acid | C9H8O4 | (165). 135. 127 | −2.32 | FP-BP | −5.65 | BP | ||
| Isoferulic acid | C10H10O4 | 178. 149. 134 | 4.29 | BP | −0.86 | BP | ||
| 1-O-Sinapoyl-beta-D-glucose | C17H22O10 | 216. 162. 119 | −4.87 | FP | −2.54 | FP | ||
| Flavonoids | Flavones | Apigenin-6-C-arabinoside-8-C-hexoside I | C26H28O14 | 397. 316. 216. 119 | 1.82 | FP | 0.94 | FP |
| Apigenin-6-C-arabinoside-8-C-hexoside II | C26H28O14 | 432. 245. 164. 149 | −1.37 | FP | −1.01 | FP | ||
| Apigenin-6-C-galactosyl-8-C-glucosyl-O-glucuropyranoside | C33H38O21 | 577. 343 | 5.3 | BP | n.d. | n.d. | ||
| Lignans | Lignans | Syringaresinol | C22H26O8 | 387. 353. 341. 257. 193. 119 | n.d. | n.d. | −1.93 | B |
| Others | Hydroxybenzaldehyde acids | 4-Hydroxybenzaldehyde | C7H6O2 | (112) | −4.89 | BP | −4.07 | B |
| Alkylphenols | 5-Nonadecenylresorcinol | C25H42O2 | 347. 309. 283 | −6.67 | FP | n.d. | n.d. | |
| 5-Nonadecylresorcinol | C25H44O2 | (355. 337. 311. 279. 248) | −4.37 | FP | −3.04 | FP | ||
| 5-Heneicosylresorcinol | C27H48O2 | (379). 339. 248 | −2.83 | FP | −4.32 | FP | ||
| Class | Sub-Class | Compound | OG | OB | SO | OH1 | OH2 | |||
|---|---|---|---|---|---|---|---|---|---|---|
| FP | BP | FP | BP | FP | BP | |||||
| Phenolic acids | Hydroxybenzoic acids | Protocatechuic acid | 1.05 ± 0.04 a | n.d. | n.d. | n.d. | n.d. | 1.71 ± 0.17 b | n.d. | n.d. |
| Hydroxybenzoic acid | n.d. | 0.75 ± 0.01 c | 0.53 ± 0.02 a | n.d. | n.d. | 0.66 ± 0.10 b | n.d. | 0.74 c | ||
| Hydroxycinnamic acids | Ferulic acid | 0.36 + 0.01 | 28.30 ± 1.56 c | 0.50 + 0.13 | 213.76 ± 4.39 e | n.d. | 32.36 ± 3.09 d | 6.15 ± 0.87b | 3.38 a | |
| p-Coumaric acid | n.d. | n.d. | n.d. | 130.67 ± 1.43 c | n.d. | n.d. | 11.47 ± 2.11 b | 7.32 a | ||
| Sinapic acid | n.d. | 2.93 ± 0.18 b | n.d. | n.d. | n.d. | 2.50 ± 0.14 a | n.d. | n.d. | ||
| Avenanthramide C | n.d. | n.d. | 0.21 ± 0.02 a | n.d. | 13.40 ± 0.34 b | 0.19 ± 0.04 a | n.d. | n.d. | ||
| Avenanthramide 2p | n.d. | n.d. | 1.58 ± 0.14 a | n.d. | 14.35 ± 0.27 b | n.d. | n.d. | n.d. | ||
| Avenanthramide 2f | 0.23 ± 0.07 a | n.d. | 1.48 ± 0.10 b | n.d. | 19.24 ± 0.19d | 1.67 ± 0.01 c | n.d. | n.d. | ||
| Diferulic isomer 1 | n.d. | 4.42 ± 0.22 b | n.d. | n.d. | n.d. | 3.80 ± 0.38 a | n.d. | n.d. | ||
| Diferulic isomer 2 | n.d. | 5.02 ± 0.63 b | n.d. | n.d. | n.d. | 3.85 ± 0.38 a | n.d. | n.d. | ||
| Diferulic isomer 4 | n.d. | 5.70 ± 0.39 a | n.d. | n.d. | n.d. | 8.35 ± 0.82 b | n.d. | n.d. | ||
| Diferulic isomer 5 | n.d. | 6.83 ± 0.59 a | n.d. | n.d. | n.d. | 7.30 ± 0.62 a | n.d. | n.d. | ||
| Diferulic isomer 6 | n.d. | 0.74 ± 0.06 a | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
| Caffeic acid | 2.32 ± 0.01 c | 1.50 ± 0.11 a | n.d. | 6.42 ± 0.26 e | n.d. | 3.44 ± 0.41 d | 2.41 ± 0.25 c | 1.72 b | ||
| Isoferulic acid | n.d. | 3.36 ± 0.67 a | n.d. | n.d. | n.d. | 2.96 ± 0.39 a | n.d. | n.d. | ||
| 1-O-Sinapoyl-beta-D-glucose | 1.55 ± 0.02 a | n.d. | n.d. | n.d. | 2.45 ± 0.02 b | n.d. | n.d. | n.d. | ||
| Flavonoids | Flavones | Apigenin-6-C-arabinoside-8-C-hexoside III | n.d. | n.d. | <LOD | n.d. | <LOD | n.d. | n.d. | n.d. |
| Others | Hydroxybenzaldehide acids | 4-Hydroxybenzaldehyde | n.d. | 1.10 ± 0.11 a | 4.17 ± 0.06 c | 27.88 ± 1.44 e | n.d. | 1.71 ± 0.11 b | 9.10 ± 1.05 d | 9.91 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimenez-Pulido, I.J.; Daniel, R.; Perez, J.; Martínez-Villaluenga, C.; De Luis, D.; Martín Diana, A.B. Impact of Protein Content on the Antioxidants, Anti-Inflammatory Properties and Glycemic Index of Wheat and Wheat Bran. Foods 2022, 11, 2049. https://doi.org/10.3390/foods11142049
Jimenez-Pulido IJ, Daniel R, Perez J, Martínez-Villaluenga C, De Luis D, Martín Diana AB. Impact of Protein Content on the Antioxidants, Anti-Inflammatory Properties and Glycemic Index of Wheat and Wheat Bran. Foods. 2022; 11(14):2049. https://doi.org/10.3390/foods11142049
Chicago/Turabian StyleJimenez-Pulido, Ivan Jesus, Rico Daniel, Jara Perez, Cristina Martínez-Villaluenga, Daniel De Luis, and Ana Belén Martín Diana. 2022. "Impact of Protein Content on the Antioxidants, Anti-Inflammatory Properties and Glycemic Index of Wheat and Wheat Bran" Foods 11, no. 14: 2049. https://doi.org/10.3390/foods11142049
APA StyleJimenez-Pulido, I. J., Daniel, R., Perez, J., Martínez-Villaluenga, C., De Luis, D., & Martín Diana, A. B. (2022). Impact of Protein Content on the Antioxidants, Anti-Inflammatory Properties and Glycemic Index of Wheat and Wheat Bran. Foods, 11(14), 2049. https://doi.org/10.3390/foods11142049