
Flaxseed Polysaccharide Alters Colonic Gene Expression of Lipid Metabolism and Energy Metabolism in Obese Rats

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
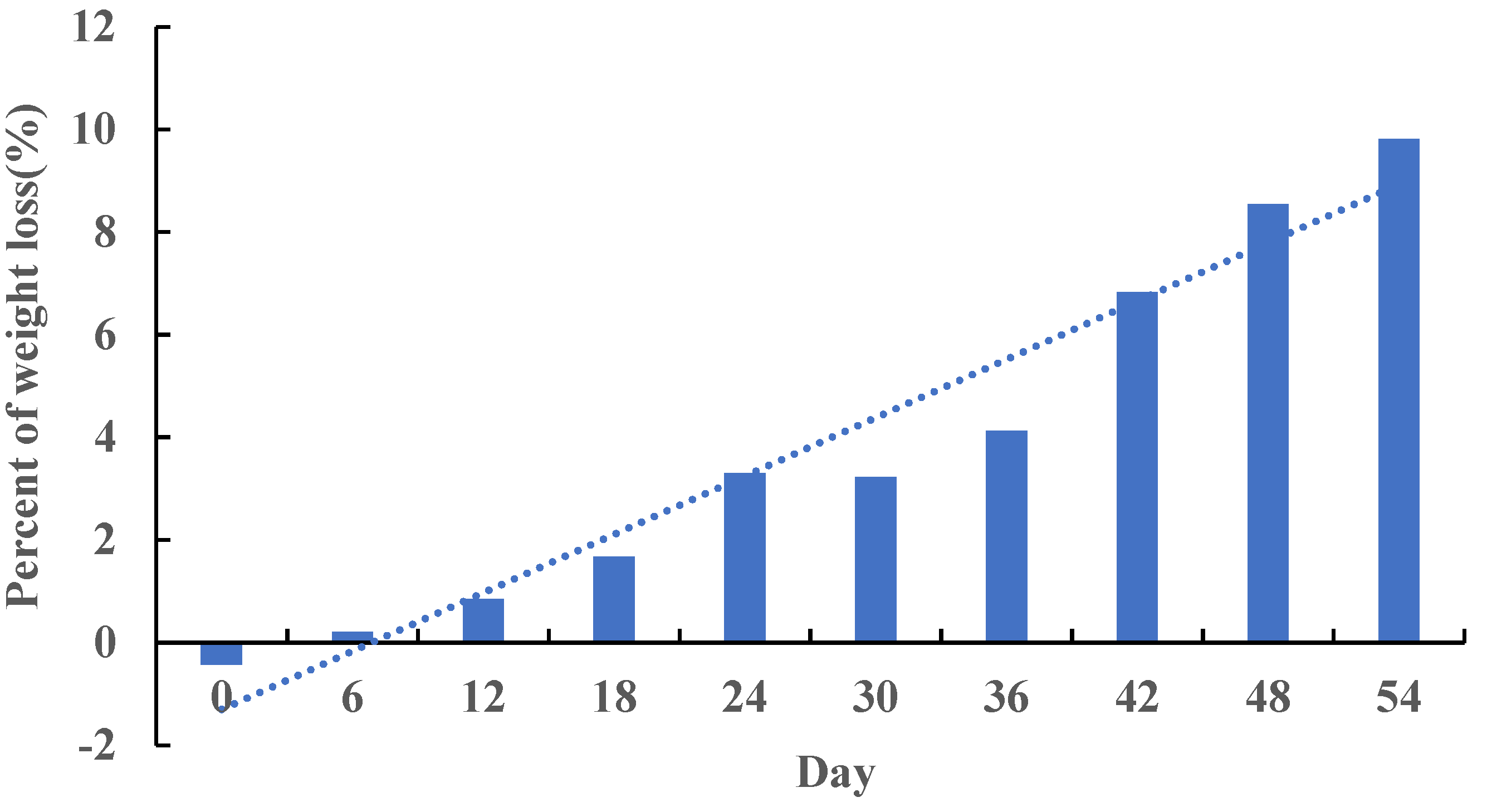
2.2. Animals, Diets, and Sample Preparation
2.3. cDNA Library Construction, RNA-seq and Bioinformatics Analysis
2.4. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.5. Statistical Analysis
3. Results
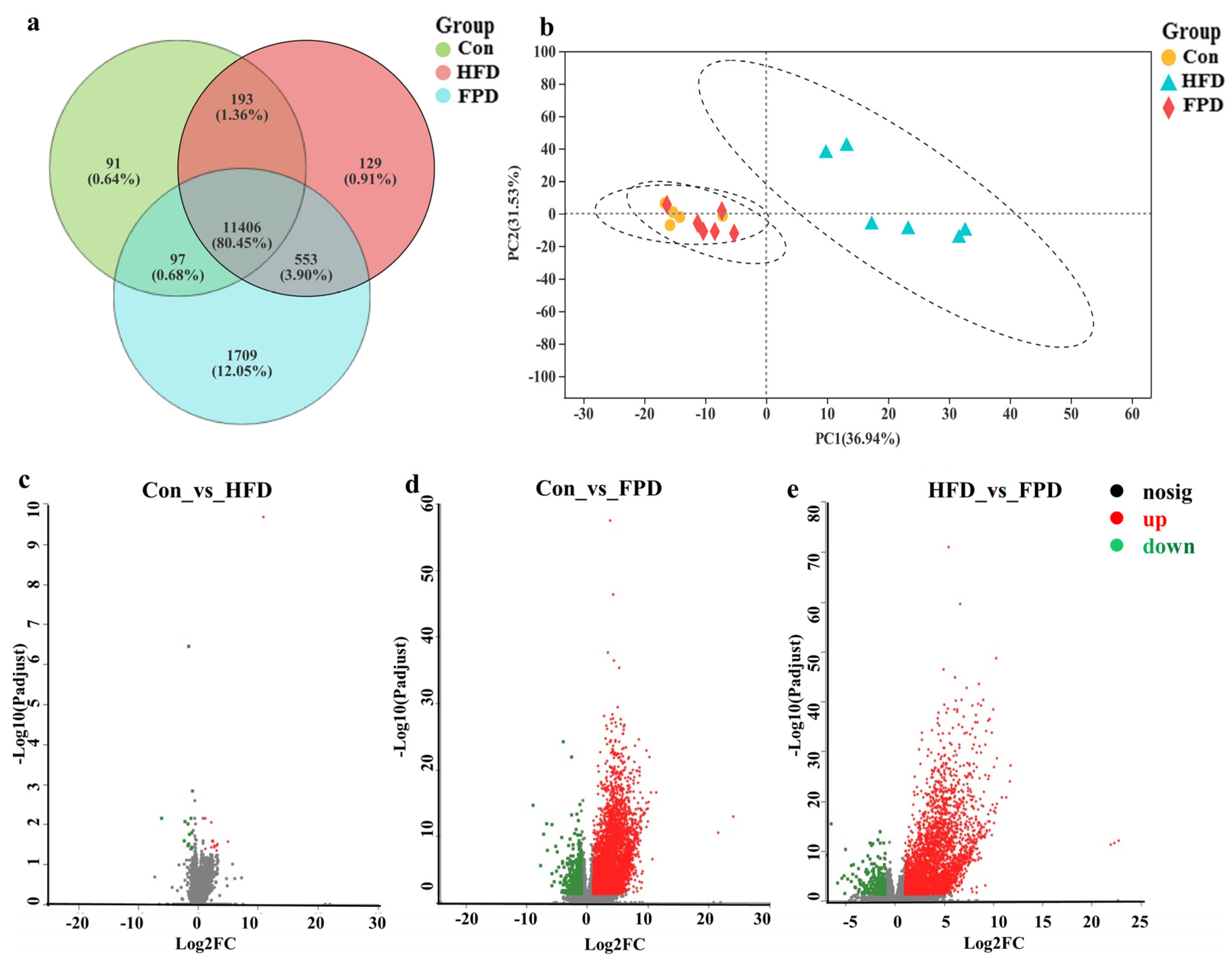
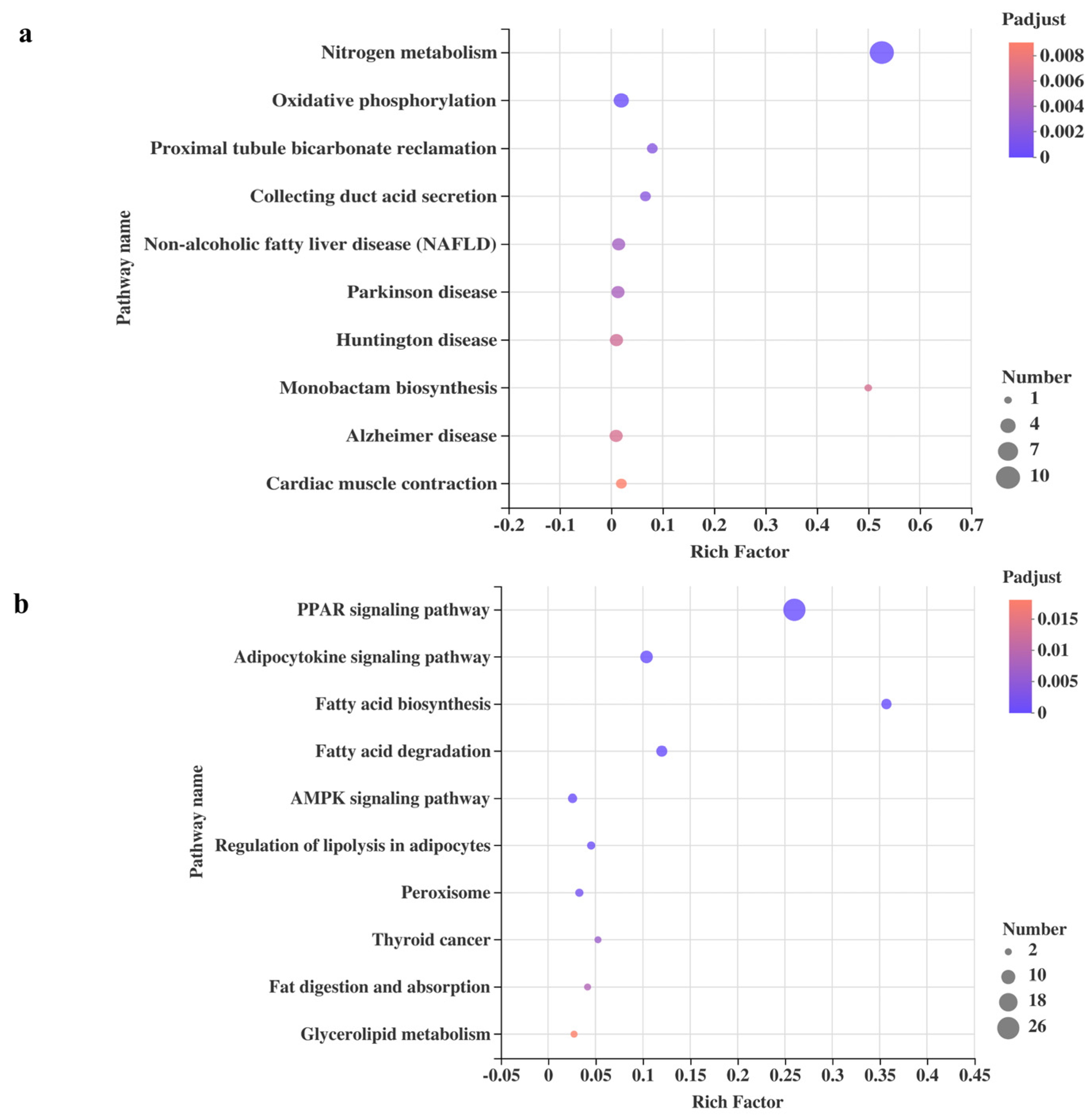
3.1. Gene Expression Profile of CETs
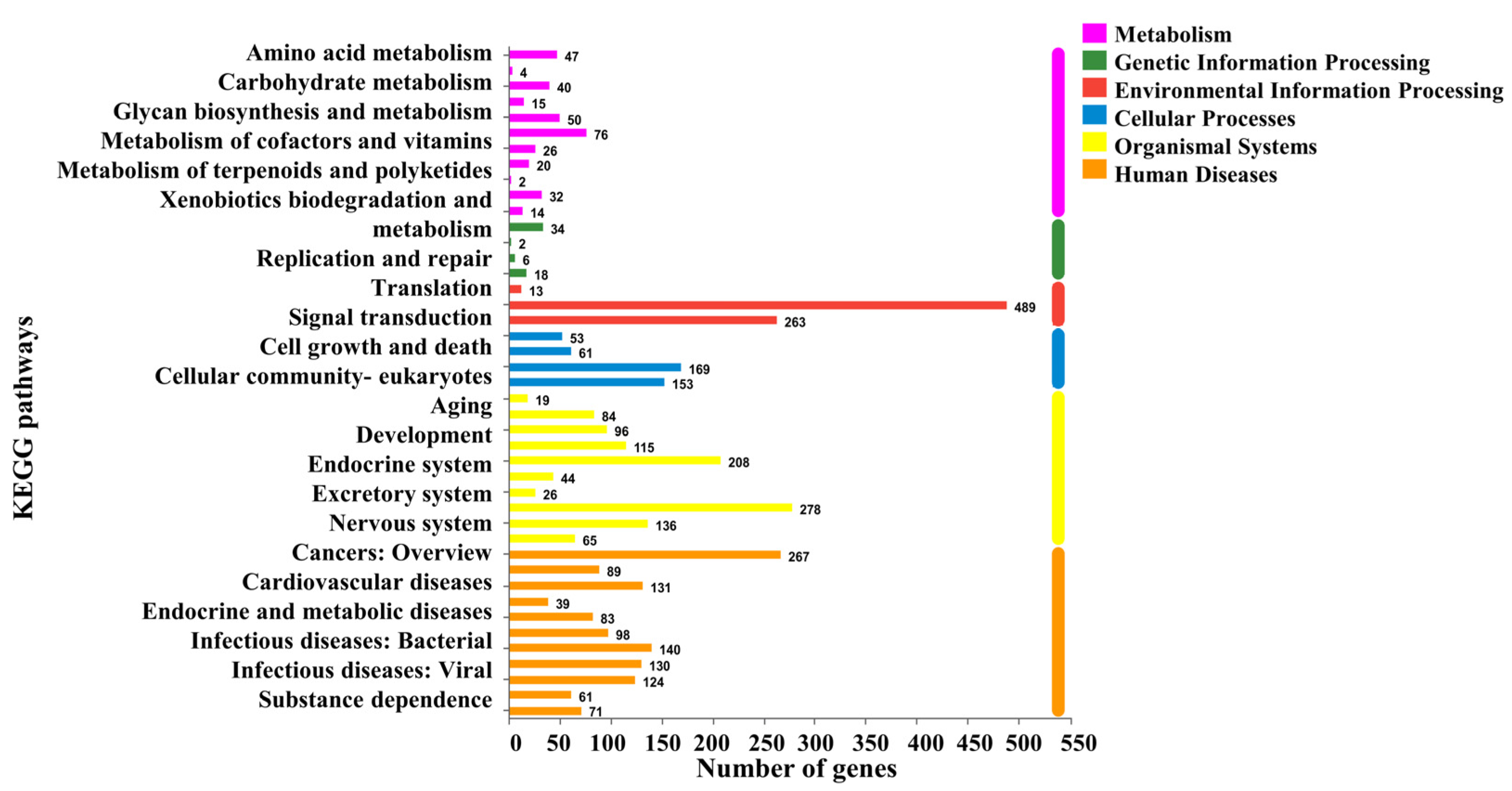
3.2. Analysis of DEGs Profile
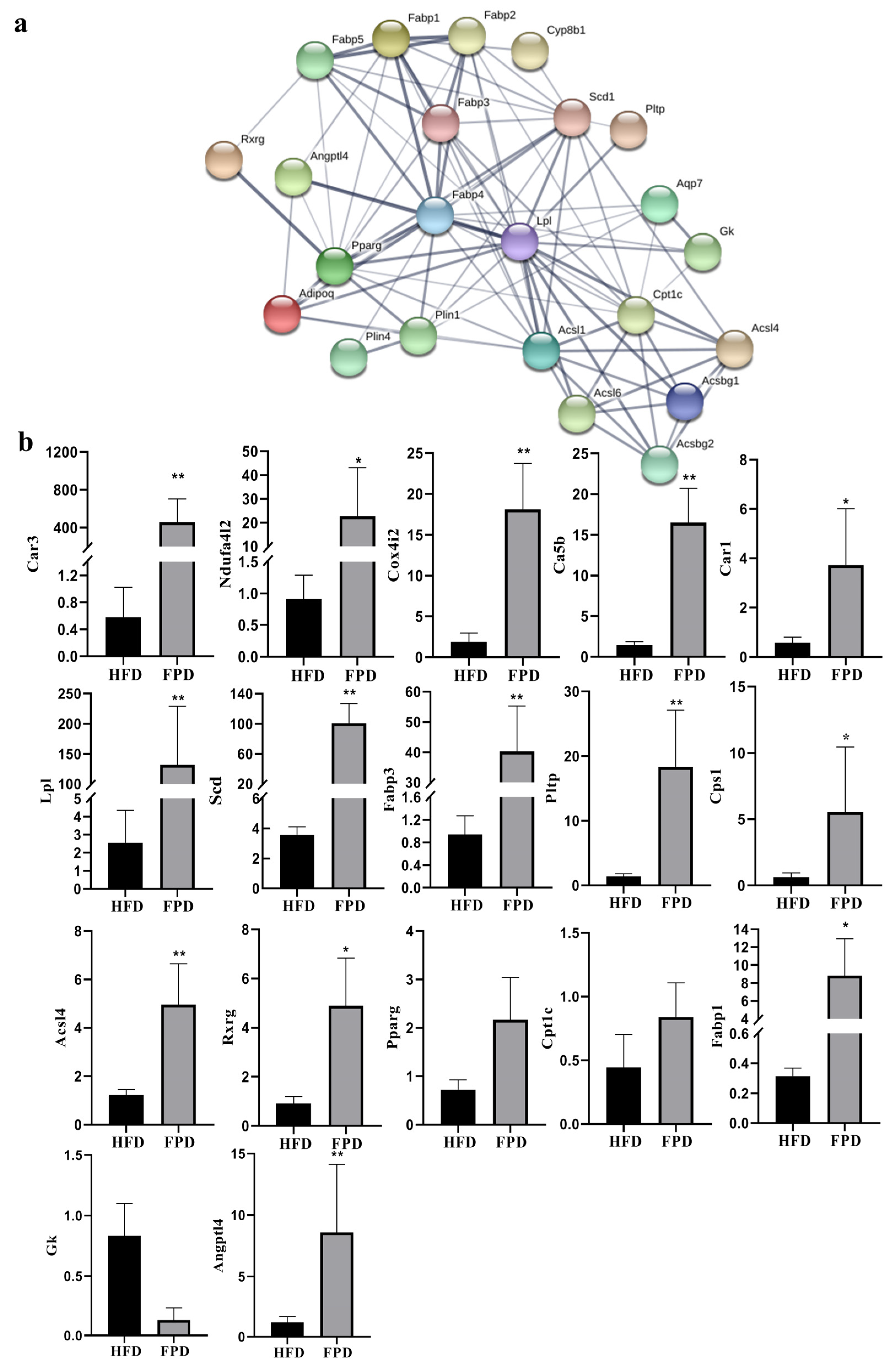
3.3. Verification of DEGs
4. Discussion
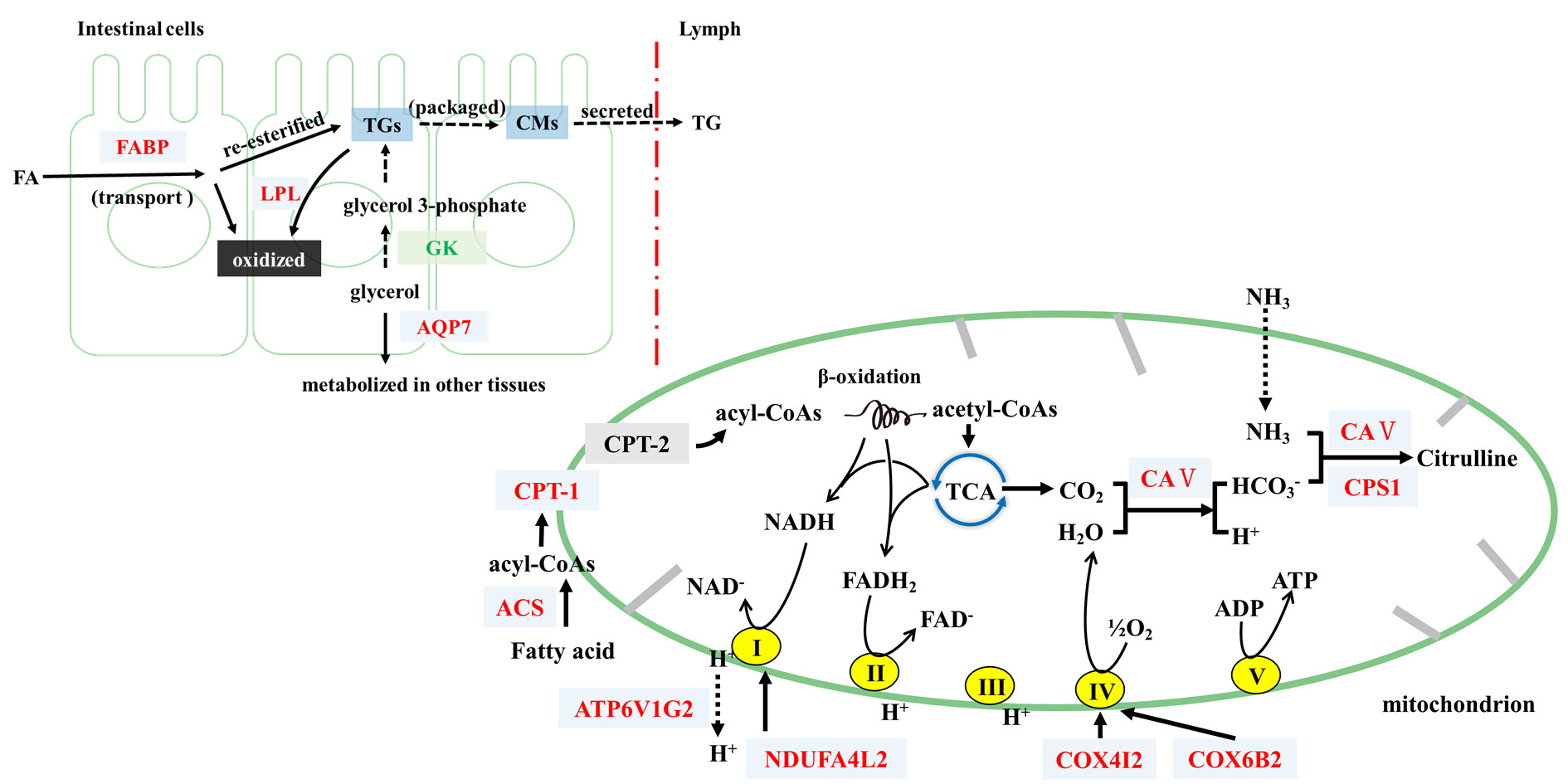
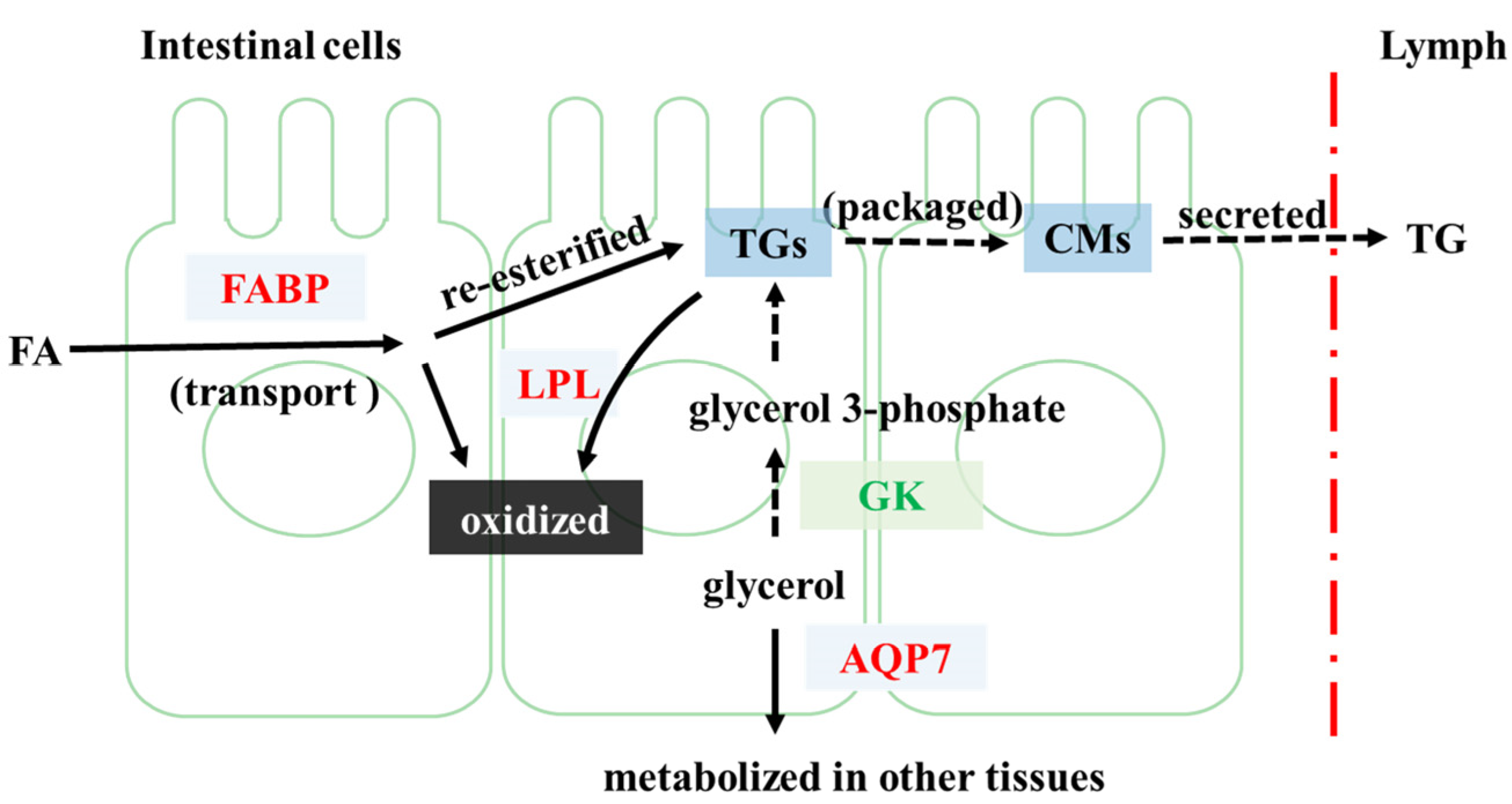
4.1. FP Intervention Regulated Lipid Metabolism of CETs
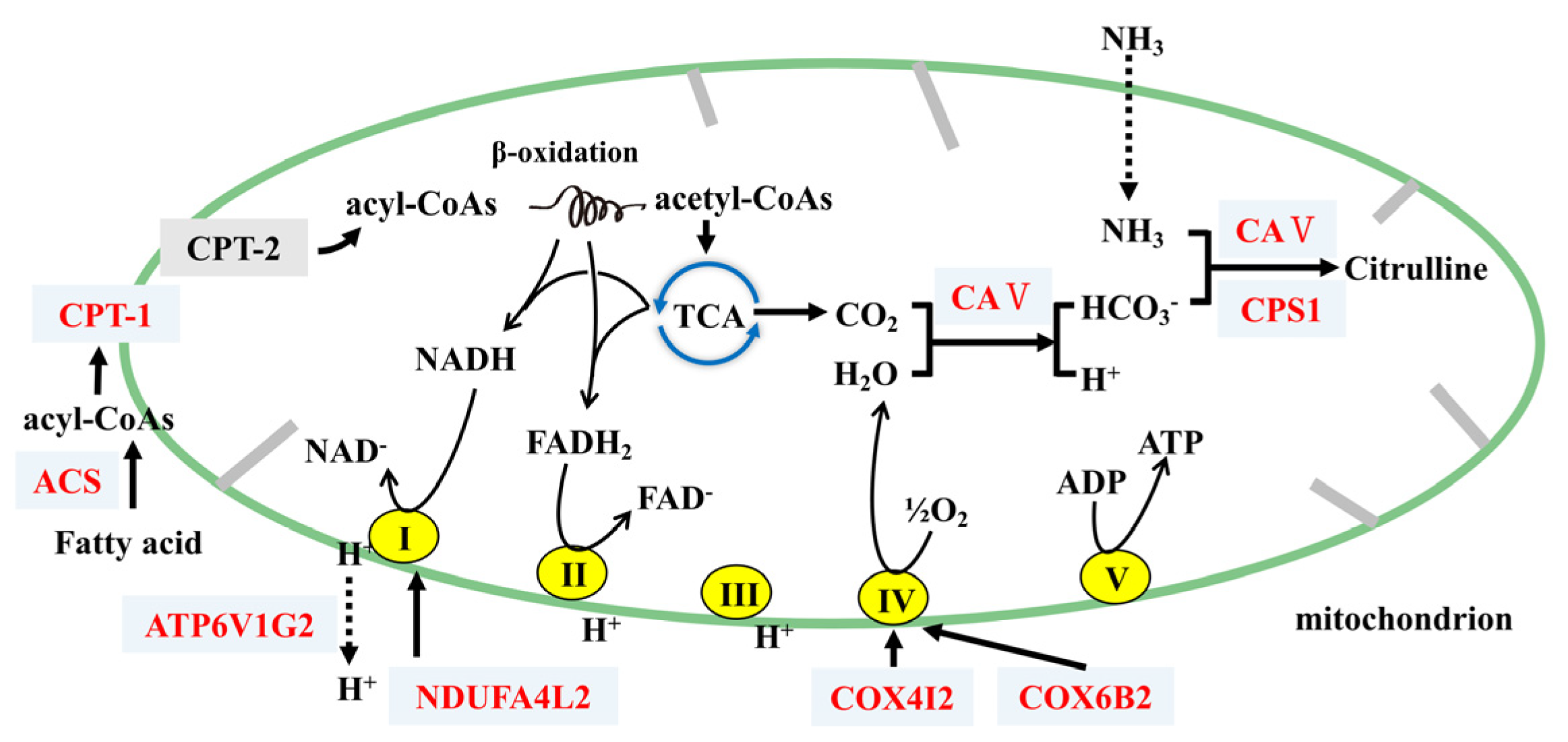
4.2. FP Intervention Regulated Energy Metabolism of CETs
 H2CO3 HCO3− + H+) [32]. Thus, FP intervention upregulated CAs and might accelerate the consumption of CO2, reducing its accumulation in cells and enhancing the tricarboxylic acid (TCA) cycle. This would further increase energy consumption. We also found that the upregulation of Cps1, which participated in the metabolism of metabolic ammonia and HCO3− into citrulline with Ca5b, plays an important part in the detoxification of intestinal ammonia [33,34].
H2CO3 HCO3− + H+) [32]. Thus, FP intervention upregulated CAs and might accelerate the consumption of CO2, reducing its accumulation in cells and enhancing the tricarboxylic acid (TCA) cycle. This would further increase energy consumption. We also found that the upregulation of Cps1, which participated in the metabolism of metabolic ammonia and HCO3− into citrulline with Ca5b, plays an important part in the detoxification of intestinal ammonia [33,34]. 5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, J.O. Understanding and addressing the epidemic of obesity: An energy balance perspective. Endocr. Rev. 2006, 27, 750–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.; Choi, H.-N.; Yim, J.-E. Effect of diet on the gut microbiota associated with obesity. J. Obes. Metab. Syndr. 2019, 28, 216. [Google Scholar] [CrossRef] [Green Version]
- Pi-Sunyer, F.X. The obesity epidemic: Pathophysiology and consequences of obesity. Obes. Res. 2002, 10, 97S–104S. [Google Scholar] [CrossRef]
- Han, S.-F.; Jiao, J.; Zhang, W.; Xu, J.-Y.; Zhang, W.; Fu, C.-L.; Qin, L.-Q. Lipolysis and thermogenesis in adipose tissues as new potential mechanisms for metabolic benefits of dietary fiber. Nutrition 2017, 33, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, J.; Gao, J.; Wu, X.; Cui, C.; Wei, H.; Zheng, R.; Peng, J. Combined soluble fiber-mediated intestinal microbiota improve insulin sensitivity of obese mice. Nutrients 2020, 12, 351. [Google Scholar] [CrossRef] [Green Version]
- Papathanasopoulos, A.; Camilleri, M. Dietary fiber supplements: Effects in obesity and metabolic syndrome and relationship to gastrointestinal functions. Gastroenterology 2010, 138, 65–72.e62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, K.; Hugenholtz, F.; Jonathan, M.C.; Schols, H.A.; Kleerebezem, M.; Smidt, H.; Müller, M.; Hooiveld, G.J. Comparison of the effects of five dietary fibers on mucosal transcriptional profiles, and luminal microbiota composition and SCFA concentrations in murine colon. Mol. Nutr. Food Res. 2015, 59, 1590–1602. [Google Scholar] [CrossRef]
- Bäckhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [Green Version]
- Safdar, B.; Pang, Z.; Liu, X.; Jatoi, M.A.; Mehmood, A.; Rashid, M.T.; Ali, N.; Naveed, M. Flaxseed gum: Extraction, bioactive composition, structural characterization, and its potential antioxidant activity. J. Food Biochem. 2019, 43, e13014. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.; Huang, F.; Yang, C.; Huang, Q. In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules 2020, 25, 4354. [Google Scholar] [CrossRef]
- Yu, J.; Hu, Q.; Liu, J.; Luo, J.; Liu, L.; Peng, X. Metabolites of gut microbiota fermenting Poria cocos polysaccharide alleviates chronic nonbacterial prostatitis in rats. Int. J. Biol. Macromol. 2022, 209 Pt B, 1593–1604. [Google Scholar] [CrossRef]
- Luo, J.; Li, Y.; Mai, Y.; Gao, L.; Ou, S.; Wang, Y.; Liu, L.; Peng, X. Flaxseed gum reduces body weight by regulating gut microbiota. J. Funct. Foods 2018, 47, 136–142. [Google Scholar] [CrossRef]
- Luo, J.; Qi, J.; Wang, W.; Luo, Z.; Liu, L.; Zhang, G.; Zhou, Q.; Liu, J.; Peng, X. Antiobesity effect of flaxseed polysaccharide via inducing satiety due to leptin resistance removal and promoting lipid metabolism through the AMP-activated protein kinase (AMPK) signaling pathway. J. Agric. Food Chem. 2019, 67, 7040–7049. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F. Purification, Structural Analysis and Its Macrophage Immunomodulatory and Anti-Hepatitis B Virus Activities. Master’s Thesis, Jinan University, Jinan, China, 2016. [Google Scholar]
- Zhang, C.; Xie, J.; Li, X.; Luo, J.; Huang, X.; Liu, L.; Peng, X. Alliin alters gut microbiota and gene expression of colonic epithelial tissues. J. Food Biochem. 2019, 43, e12795. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Xuan, Y.P.; Gao, B.; Sun, X.; Miao, S.C.; Lu, T.; Wang, Y.Y.; Jiao, W.J. Identification of an eight-gene prognostic signature for lung adenocarcinoma. Cancer Manag. Res. 2018, 10, 3383–3392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Fei, N.; Wu, G.; Zhang, C.; Zhao, L.; Zhang, M. Regulated inflammation and lipid metabolism in colon mRNA expressions of obese germfree mice responding to Enterobacter cloacae B29 combined with the high fat diet. Front. Microbiol. 2016, 7, 1786. [Google Scholar] [CrossRef] [Green Version]
- Amiri, M.; Yousefnia, S.; Forootan, F.S.; Peymani, M.; Ghaedi, K.; Esfahani, M.H.N. Diverse roles of fatty acid binding proteins (FABPs) in development and pathogenesis of cancers. Gene 2018, 676, 171–183. [Google Scholar] [CrossRef]
- Duszka, K.; Oresic, M.; Le May, C.; König, J.; Wahli, W. PPARγ modulates long chain fatty acid processing in the intestinal epithelium. Int. J. Mol. Sci. 2017, 18, 2559. [Google Scholar] [CrossRef] [Green Version]
- Demignot, S.; Beilstein, F.; Morel, E. Triglyceride-rich lipoproteins and cytosolic lipid droplets in enterocytes: Key players in intestinal physiology and metabolic disorders. Biochimie 2014, 96, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Lambert, J.E.; Parks, E.J. Postprandial metabolism of meal triglyceride in humans. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2012, 1821, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Li, J.; Zou, J.; Qiu, B.; Liu, W.; Lin, X.; Li, D.; Liu, Z.; Du, F. Rat Small Intestinal Mucosal Epithelial Cells Absorb Dietary 1, 3-Diacylglycerol Via Phosphatidic Acid Pathways. Lipids 2018, 53, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Iena, F.M.; Lebeck, J. Implications of aquaglyceroporin 7 in energy metabolism. Int. J. Mol. Sci. 2018, 19, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dávalos-Salas, M.; Montgomery, M.K.; Reehorst, C.M.; Nightingale, R.; Ng, I.; Anderton, H.; Al-Obaidi, S.; Lesmana, A.; Scott, C.M.; Ioannidis, P. Deletion of intestinal Hdac3 remodels the lipidome of enterocytes and protects mice from diet-induced obesity. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of energy metabolism by long-chain fatty acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef]
- Dai, J.; Liang, K.; Zhao, S.; Jia, W.; Liu, Y.; Wu, H.; Lv, J.; Cao, C.; Chen, T.; Zhuang, S. Chemoproteomics reveals baicalin activates hepatic CPT1 to ameliorate diet-induced obesity and hepatic steatosis. Proc. Natl. Acad. Sci. USA 2018, 115, E5896–E5905. [Google Scholar] [CrossRef] [Green Version]
- Kuwata, H.; Nakatani, E.; Shimbara-Matsubayashi, S.; Ishikawa, F.; Shibanuma, M.; Sasaki, Y.; Yoda, E.; Nakatani, Y.; Hara, S. Long-chain acyl-CoA synthetase 4 participates in the formation of highly unsaturated fatty acid-containing phospholipids in murine macrophages. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 1606–1618. [Google Scholar] [CrossRef]
- Ducheix, S.; Peres, C.; Härdfeldt, J.; Frau, C.; Mocciaro, G.; Piccinin, E.; Lobaccaro, J.-M.; De Santis, S.; Chieppa, M.; Bertrand-Michel, J. Deletion of stearoyl-CoA desaturase-1 from the intestinal epithelium promotes inflammation and tumorigenesis, reversed by dietary oleate. Gastroenterology 2018, 155, 1524–1538.e1529. [Google Scholar] [CrossRef]
- Phua, T.; Sng, M.K.; Tan, E.H.P.; Chee, D.S.L.; Li, Y.; Wee, J.W.K.; Teo, Z.; Chan, J.S.K.; Lim, M.M.K.; Tan, C.K. Angiopoietin-like 4 mediates colonic inflammation by regulating chemokine transcript stability via tristetraprolin. Sci. Rep. 2017, 7, 1–16. [Google Scholar]
- Smetnev, S.; Klimushina, M.; Kutsenko, V.; Kiseleva, A.; Gumanova, N.; Kots, A.; Skirko, O.; Ershova, A.; Yarovaya, E.; Metelskaya, V. Associations of SNPs of the ADIPOQ Gene with Serum Adiponectin Levels, Unstable Angina, and Coronary Artery Disease. Biomolecules 2019, 9, 537. [Google Scholar] [CrossRef] [Green Version]
- Sohn, J.H.; Lee, Y.K.; Han, J.S.; Jeon, Y.G.; Kim, J.I.; Choe, S.S.; Kim, S.J.; Yoo, H.J.; Kim, J.B. Perilipin 1 (Plin1) deficiency promotes inflammatory responses in lean adipose tissue through lipid dysregulation. J. Biol. Chem. 2018, 293, 13974–13988. [Google Scholar] [CrossRef] [Green Version]
- Renner, S.W.; Walker, L.M.; Forsberg, L.J.; Sexton, J.Z.; Brenman, J.E. Carbonic anhydrase III (Car3) is not required for fatty acid synthesis and does not protect against high-fat diet induced obesity in mice. PLoS ONE 2017, 12, e0176502. [Google Scholar]
- Saarnio, J.; Parkkila, S.; Parkkila, A.-K.; Waheed, A.; Karttunen, T.; Sly, W.S. Cell-specific expression of mitochondrial carbonic anhydrase in the human and rat gastrointestinal tract. J. Histochem. Cytochem. 1999, 47, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillman, J.B.; Dhahbi, J.M.; Mote, P.L.; Walford, R.L.; Spindler, S.R. Dietary calorie restriction in mice induces carbamyl phosphate synthetase I gene transcription tissue specifically. J. Biol. Chem. 1996, 271, 3500–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, S.E.; Sena, L.A.; Chandel, N.S. Mitochondria in the regulation of innate and adaptive immunity. Immunity 2015, 42, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.; Yang, X.; Xie, X.; Wang, M. Mitochondrial NDUFA4L2 protein promotes the vitality of lung cancer cells by repressing oxidative stress. Thorac. Cancer 2019, 10, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Nie, K.; Li, J.; He, X.; Wang, Y.; Zhao, Q.; Du, M.; Sun, H.; Wang, J.; Lyu, J.; Fang, H. COX6B2 drives metabolic reprogramming toward oxidative phosphorylation to promote metastasis in pancreatic ductal cancer cells. Oncogenesis 2020, 9, 1–13. [Google Scholar] [CrossRef]
- Kane, P.M. Targeting reversible disassembly as a mechanism of controlling V-ATPase activity. Curr. Protein Pept. Sci. 2012, 13, 117–123. [Google Scholar] [CrossRef]
- Fuhrmann, D.C.; Brüne, B. Mitochondrial composition and function under the control of hypoxia. Redox Biol. 2017, 12, 208–215. [Google Scholar] [CrossRef]
- Evans, A.M.; Mahmoud, A.D.; Moral-Sanz, J.; Hartmann, S. The emerging role of AMPK in the regulation of breathing and oxygen supply. Biochem. J. 2016, 473, 2561–2572. [Google Scholar] [CrossRef] [Green Version]
- Adiyaman, S.C.; Ozer, M.; Saydam, B.O.; Akinci, B. The Role of Adiponectin in Maintaining Metabolic Homeostasis. Curr. Diabetes Rev. 2020, 16, 95–103. [Google Scholar] [CrossRef]
- Ohashi, K.; Yuasa, D.; Shibata, R.; Murohara, T.; Ouchi, N. Adiponectin as a Target in Obesity-related Inflammatory State. Endocr. Metab. Immune Disord. Drug Targets 2015, 15, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Guan, X. The CNS glucagon-like peptide-2 receptor in the control of energy balance and glucose homeostasis. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2014, 307, R585–R596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, G.S.; Putaala, H.; Hibberd, A.A.; Alhoniemi, E.; Tiihonen, K.; Mäkelä, K.A.; Herzig, K.-H. Polydextrose changes the gut microbiome and attenuates fasting triglyceride and cholesterol levels in Western diet fed mice. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Zapata, R.C.; Pezeshki, A.; Reidelberger, R.D.; Chelikani, P.K. Inulin fiber dose-dependently modulates energy balance, glucose tolerance, gut microbiota, hormones and diet preference in high-fat-fed male rats. J. Nutr. Biochem. 2018, 59, 142–152. [Google Scholar] [CrossRef]
- Franzago, M.; Santurbano, D.; Vitacolonna, E.; Stuppia, L. Genes and Diet in the Prevention of Chronic Diseases in Future Generations. Int. J. Mol. Sci. 2020, 21, 2633. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Standard Diet (D12450B) | High-Fat Diet (D12492) | Control Diet (AIN-93M) | Flaxseed Polysaccharide Diet |
|---|---|---|---|---|
| flaxseed polysaccharide | 0.00 | 0.00 | 0.00 | 10.00 |
| corn starch | 33.00 | 0.00 | 46.57 | 36.57 |
| dextrin | 3.35 | 16.35 | 15.50 | 15.50 |
| casein | 19.13 | 26.17 | 14.00 | 14.00 |
| sucrose | 34.47 | 9.00 | 10.00 | 10.00 |
| cellulose | 4.78 | 6.54 | 5.00 | 5.00 |
| soybean oil | 2.39 | 3.27 | 4.00 | 4.00 |
| lard | 1.91 | 32.06 | 0.00 | 0.00 |
| mineral mix ain-93 | 3.35 | 4.58 | 3.50 | 3.50 |
| vitamin mix ain-93 | 0.96 | 1.31 | 1.00 | 1.00 |
| L-cystine | 0.29 | 0.39 | 0.18 | 0.18 |
| choline bitartrate | 0.24 | 0.33 | 0.25 | 0.25 |
| Gene | Accession Number | F/P | Sequence (5′-3′) | Product Length |
|---|---|---|---|---|
| β-actin | NM_031144.3 | Forward Primer | ATTGCTGACAGGATGCAGAA | 109 bp |
| Reverse Primer | TAGAGCCACCAATCCACACAG | |||
| Pparg | NM_013124.3 | Forward Primer | GTCTCACAATGCCATCAGGT | 87 bp |
| Reverse Primer | AGCTGGTCGATATCACTGGA | |||
| Fabp1 | NM_012556.2 | Forward Primer | CCAAGAGAACTTTGAGCCCT | 151 bp |
| Reverse Primer | ATTGTGGATCACCTTGGACC | |||
| Rxrg | NM_031765.1 | Forward Primer | ATCCCAGCTACACAGATACCC | 111 bp |
| Reverse Primer | GACCCATGGCAGAAGTGATG | |||
| Cpt1c | NM_001034925.2 | Forward Primer | GTCTTCACTCAGTTCCGACG | 117 bp |
| Reverse Primer | AAGTCATTCCAGACACGCC | |||
| Pltp | NM_001168543.1 | Forward Primer | TTGTACCATCAAGCCGTCAG | 89 bp |
| Reverse Primer | GCTCGACTTCAGGCATTGTA | |||
| Ca5b | NM_001005551.2 | Forward Primer | CTTGAACAGTTTCGGACCCT | 103 bp |
| Reverse Primer | GGACAGTGCGATTCATCAGA | |||
| Cps1 | NM_017072.2 | Forward Primer | AGCCGTTGTTTGGAATCAGT | 80 bp |
| Reverse Primer | GGCCATGGACATCTTGTAGG | |||
| Scd | NM_139192.2 | Forward Primer | CGTCAGCACCTTCTTGAGATA | 168 bp |
| Reverse Primer | GCGTGATGGTAGTTGTGGAA | |||
| Angptl4 | NM_199115.2 | Forward Primer | CCACCAATGTTTCCCCCAAT | 92 bp |
| Reverse Primer | GCTCTTGGCACAGTTAAGGT | |||
| Fabp3 | NM_024162.2 | Forward Primer | GCTGGGAGTAGAGTTTGACG | 139 bp |
| Reverse Primer | CCCATCACTTAGTTCCCGTG | |||
| Car1 | NM_001107660.1 | Forward Primer | GTCCTGACCAATGGAGCAAA | 92 bp |
| Reverse Primer | GGAATCATGTTTGGCTTCGC | |||
| Gk | NM_024381.2 | Forward Primer | ATGGCCTAATGAAAGCTGGG | 222 bp |
| Reverse Primer | CAGGTTTGTCTCTGCCAAGT | |||
| Ndufa4l2 | NM_001271272.1 | Forward Primer | ATGGCAGGAACCAGTCTAGG | 79 bp |
| Reverse Primer | TGAAGCCGATCATTGGGATG | |||
| Cox4i2 | NM_053472.1 | Forward Primer | TTCGCAGAGATGAACCATCG | 87 bp |
| Reverse Primer | AATCACCAGAGCCGTGAATC | |||
| Lpl | NM_012598.2 | Forward Primer | TTGGCTCCAGAGTTTGAC | 145 bp |
| Reverse Primer | TGCTAATCCAGGAATCAGA | |||
| Acsl4 | NM_053623.1 | Forward Primer | CCGAGTGAATAACTTTGGA | 200 bp |
| Reverse Primer | AGGAAGCCTCAGACTCAT | |||
| Car3 | NM_019292.4 | Forward Primer | GCTCTGCTAAGACCATCC | 117 bp |
| Reverse Primer | ATTGGCGAAGTCGGTAGG |
| KEGG Pathway | Gene Name | Gene Description | Regulation |
|---|---|---|---|
| Oxidative phosphorylation | Ndufa4l2 | NDUFA4, mitochondrial complex associated like 2 | Upregulated |
| Atp6v1g2 | ATPase H+ transporting V1 subunit G2 | Upregulated | |
| Cox4i2 | cytochrome c oxidase subunit 4i2 | Upregulated | |
| Cox6b2 | cytochrome c oxidase subunit VI b polypeptide 2 | Upregulated | |
| Nitrogen metabolism | Cps1 | carbamoyl-phosphate synthase 1 | Upregulated |
| Car3 | carbonic anhydrase 3 | Upregulated | |
| Car1 | carbonic anhydrase I | Upregulated | |
| Ca5b | carbonic anhydrase 5B | Upregulated | |
| Car8 | carbonic anhydrase 8 | Upregulated | |
| Car13 | carbonic anhydrase 13 | Upregulated | |
| Car2 | carbonic anhydrase 2 | Upregulated | |
| Car12 | carbonic anhydrase 12 | Upregulated | |
| Car14 | carbonic anhydrase 14 | Upregulated | |
| Car4 | carbonic anhydrase 4 | Upregulated | |
| PPAR-signaling pathway | Plin4 | perilipin 4 | Upregulated |
| Fabp1 | fatty acid binding protein 1 | Upregulated | |
| Fabp3 | fatty acid binding protein 3 | Upregulated | |
| Cyp8b1 | cytochrome P450, family 8, subfamily b, polypeptide 1 | Upregulated | |
| Scd | stearoyl-CoA desaturase | Upregulated | |
| Fabp4 | fatty acid binding protein 4 | Upregulated | |
| Lpl | lipoprotein lipase | Upregulated | |
| Acsl6 | acyl-CoA synthetase long-chain family member 6 | Upregulated | |
| Plin1 | perilipin 1 | Upregulated | |
| Pltp | phospholipid transfer protein | Upregulated | |
| Angptl4 | angiopoietin-like 4 | Upregulated | |
| Rxrg | retinoid X receptor gamma | Upregulated | |
| Adipoq | adiponectin, C1Q and collagen domain containing | Upregulated | |
| Acsbg2 | acyl-CoA synthetase bubblegum family member 2 | Upregulated | |
| Fabp5 | fatty acid binding protein 5, epidermal | Upregulated | |
| Fabp2 | fatty acid binding protein 2 | Upregulated | |
| Aqp7 | aquaporin 7 | Upregulated | |
| Cpt1c | carnitine palmitoyl transferase 1c | Upregulated | |
| Acsl4 | acyl-CoA synthetase long-chain family member 4 | Upregulated | |
| Pparg | peroxisome proliferator-activated receptor gamma | Upregulated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Lin, X.; Liu, L.; Peng, X. Flaxseed Polysaccharide Alters Colonic Gene Expression of Lipid Metabolism and Energy Metabolism in Obese Rats. Foods 2022, 11, 1991. https://doi.org/10.3390/foods11131991
Wei H, Lin X, Liu L, Peng X. Flaxseed Polysaccharide Alters Colonic Gene Expression of Lipid Metabolism and Energy Metabolism in Obese Rats. Foods. 2022; 11(13):1991. https://doi.org/10.3390/foods11131991
Chicago/Turabian StyleWei, Hua, Xiaohong Lin, Liu Liu, and Xichun Peng. 2022. "Flaxseed Polysaccharide Alters Colonic Gene Expression of Lipid Metabolism and Energy Metabolism in Obese Rats" Foods 11, no. 13: 1991. https://doi.org/10.3390/foods11131991
APA StyleWei, H., Lin, X., Liu, L., & Peng, X. (2022). Flaxseed Polysaccharide Alters Colonic Gene Expression of Lipid Metabolism and Energy Metabolism in Obese Rats. Foods, 11(13), 1991. https://doi.org/10.3390/foods11131991