Sirtfoods: New Concept Foods, Functions, and Mechanisms


Abstract
:1. Introduction
2. SIRT Health Functions
2.1. SIRT Genes
2.2. SIRTs and Oxidative Stress
2.3. SIRTs and Apoptosis
2.4. SIRTs and Autophagy
2.5. SIRTs and Aging
2.6. SIRTs and Inflammation
2.7. SIRTs and Viral Infection
2.8. SIRTs and Energy Homeostasis
2.9. SIRTs and Cancer
3. Sirtfoods and SIRT Modulations
3.1. The Relationship between Sirtfoods and Gene Modulation
3.2. Natural Compounds Modulate SIRTs
4. Mechanisms of Sirtfoods
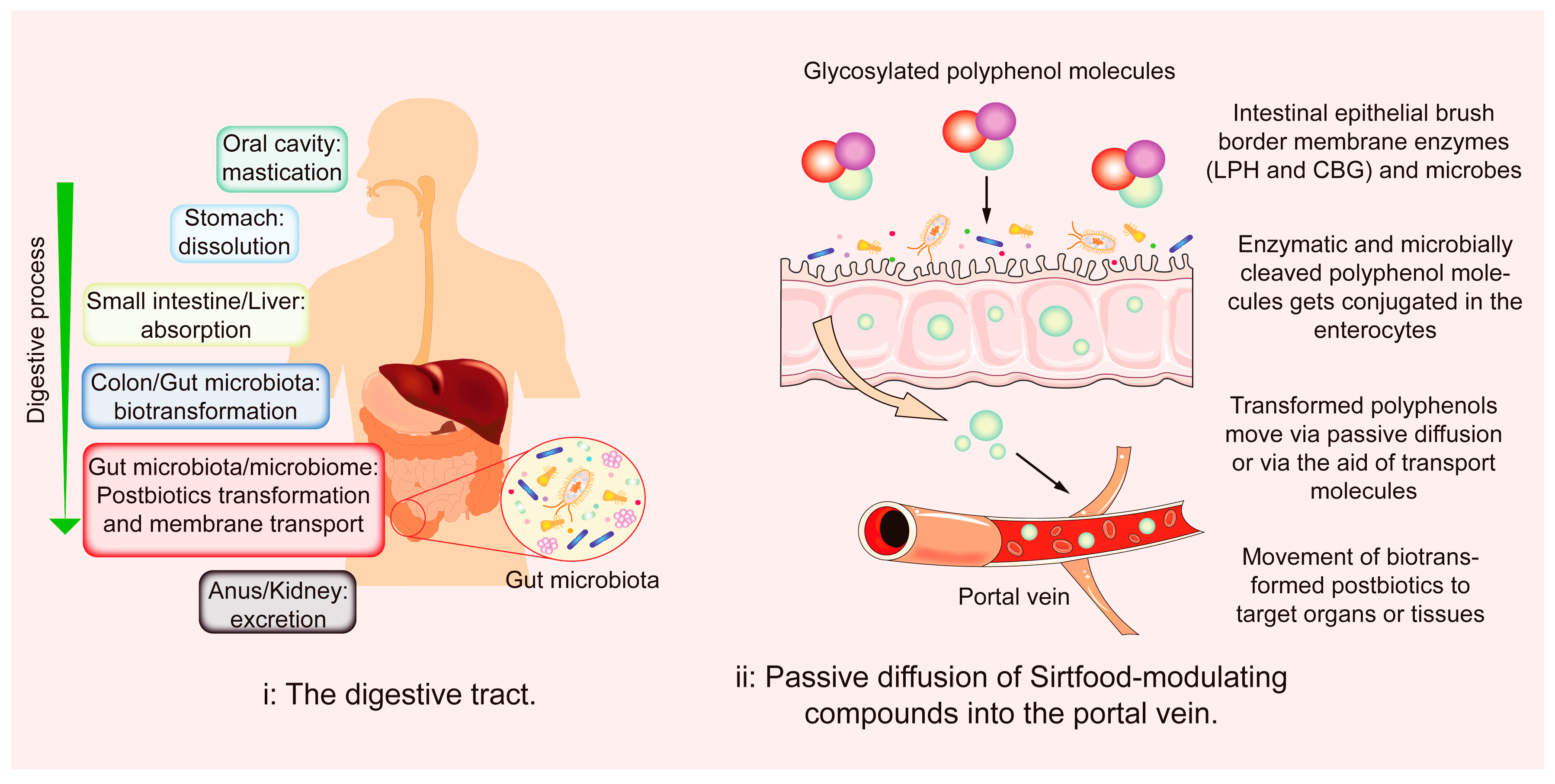
4.1. The Gut Environment
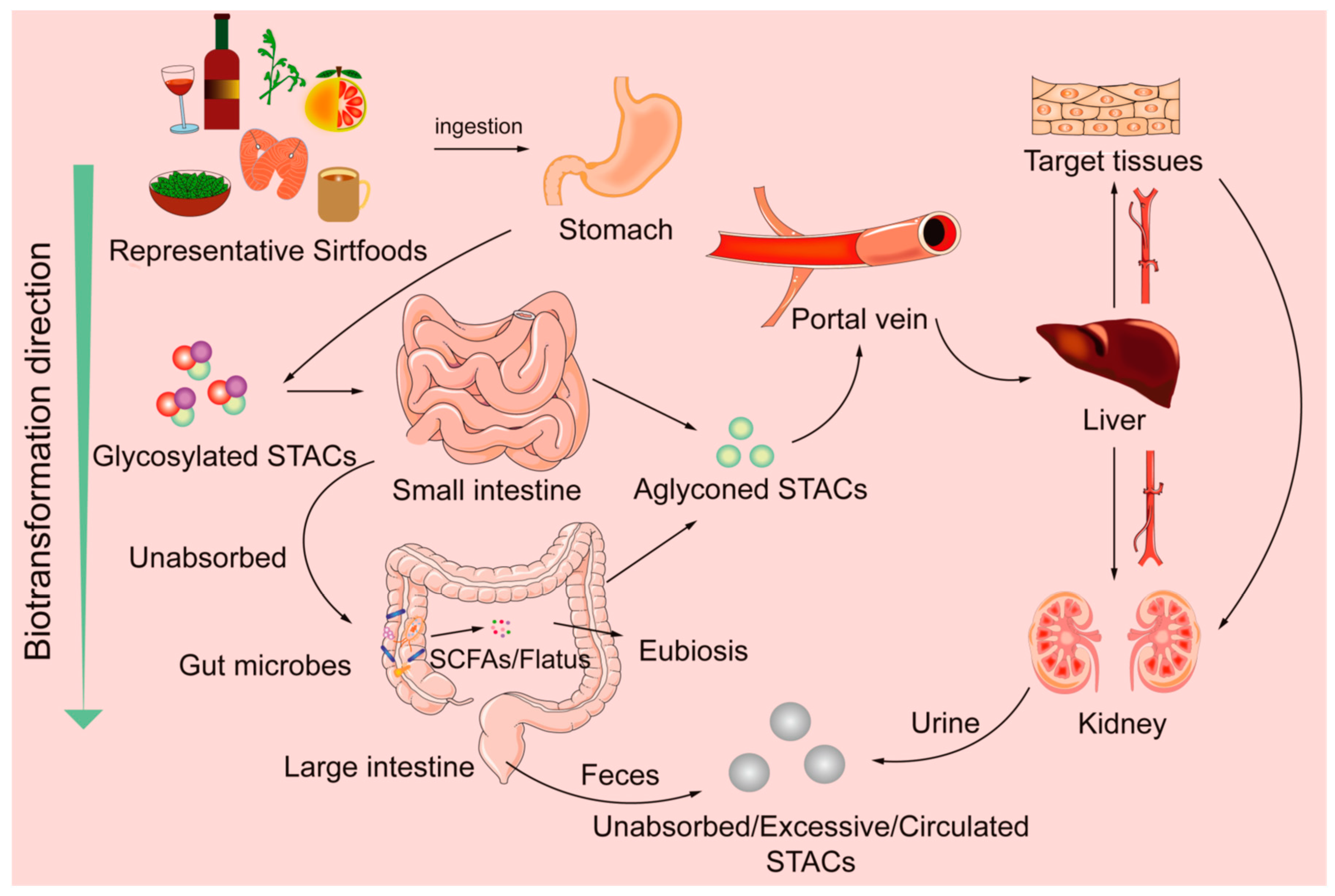
4.2. Abridged Sirtfoods’ Metabolisms
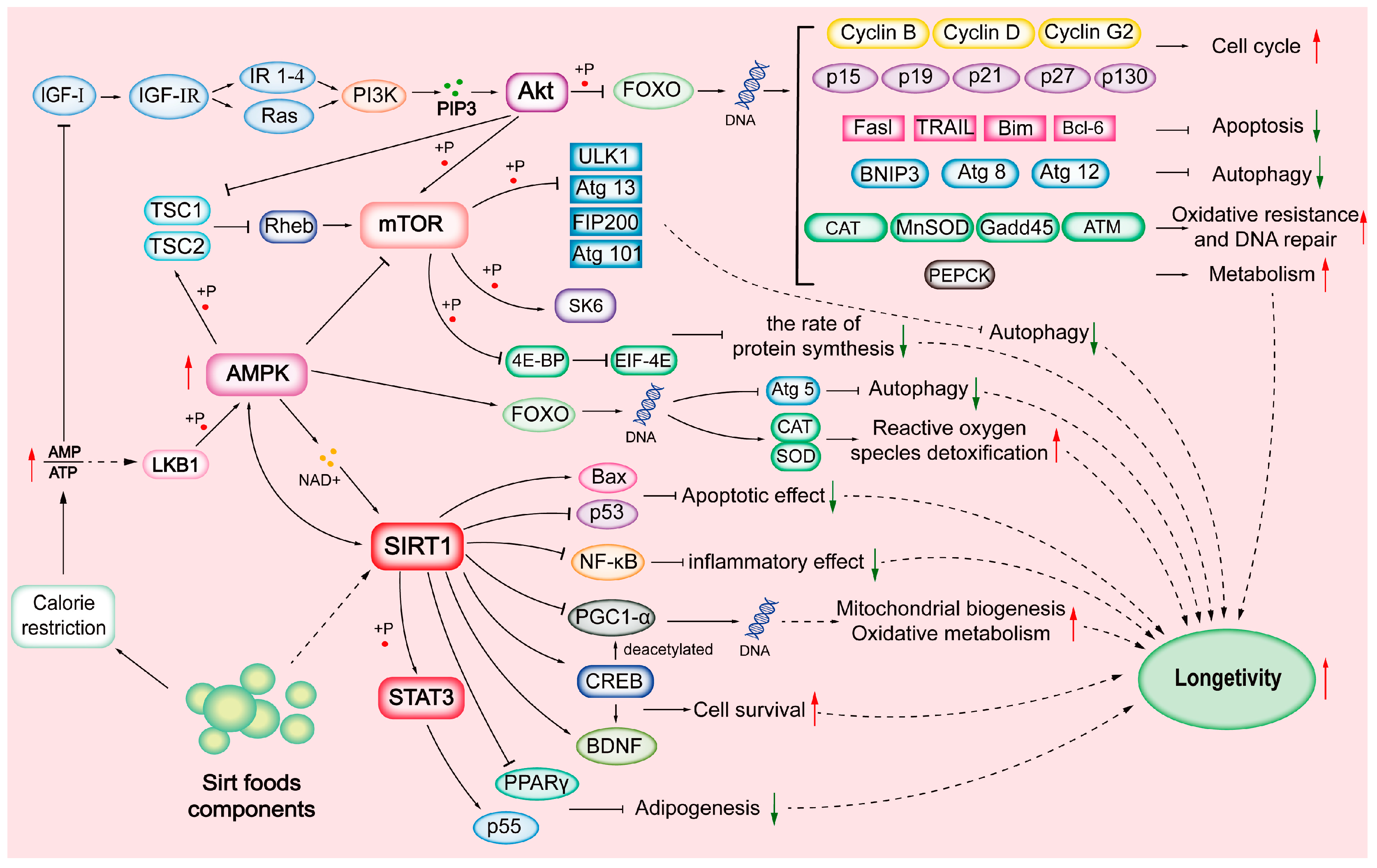
4.3. Bioactivity Regulations of SIRTs
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Betts, J.A.; Gonzalez, J.T. Personalized nutrition: What makes you so special? Nutr. Bull. 2017, 41, 353–359. [Google Scholar] [CrossRef]
- Rajasekaran, A.; Kalaivani, M. Designer foods and their benefits: A review. J. Food Sci. Technol. 2013, 50, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy effects of plant polyphenols: Molecular mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef]
- Olovo, C.V.; Udoekong, N.S.; Akan, O.D. Precision nutrition, diet and gut-microbiota in obesity. J. Biotech. Biores. 2021, 2, 1–3. [Google Scholar]
- Del Bo‘, C.; Bernardi, S.; Marino, M.; Porrini, M.; Tucci, M.; Guglielmetti, S.; Cherubini, A.; Carrieri, B.; Kirkup, B.; Kroon, P.; et al. Systematic review on polyphenol intake and health outcomes: Is there sufficient evidence to define a health-promoting polyphenol-rich dietary pattern? Nutrients 2019, 11, 1355. [Google Scholar]
- Jones, T. The Sirtfood Diet: A Detailed Beginner’s Guide. Healthline-Nutrition. 2020. Available online: https://www.healthline.com/nutrition/sirtfood-diet (accessed on 12 August 2022).
- Pallauf, K.; Giller, K.; Huebbe, P.; Rimbach, G. Nutrition and healthy ageing: Calorie restriction or polyphenol-rich “MediterrAsian” diet? Oxid. Medi. Cell. Longev. 2013, 2013, 707421. [Google Scholar] [CrossRef]
- Ahmed, M.A.; O’Callaghan, C.; Chang, E.D.; Jiang, H.; Vassilopoulos, A. Context-dependent roles for SIRT2 and SIRT3 in tumor development upon calorie restriction or high fat diet. Front. Oncol. 2020, 9, 1462. [Google Scholar] [CrossRef]
- Poulose, N.; Raju, R. Sirtuin regulation in aging and injury. BBA-Mol. Basis Dis. 2015, 1852, 2442–2455. [Google Scholar] [CrossRef] [PubMed]
- Lilja, S.; Pointner, A.; Bäck, H.; Duszka, K.; Hippe, B.; Suarez, L.; Höfinger, I.; Debebe, T.; König, J.; Haslberger, A.G. Fasting and fasting mimetic supplementation address sirtuin expression, miRNA and microbiota composition. Funct. Foods Health Dis. 2020, 10, 439–455. [Google Scholar] [CrossRef]
- Wei, M.; Brandhorst, S.; Shelehchi, M.; Mirzaei, H.; Cheng, C.W.; Budniak, J.; Groshen, S.; Mack, W.J.; Guen, E.; Di Biase, S.; et al. Fasting-mimicking diet and markers/risk factors for aging, diabetes, cancer, and cardiovascular disease. Sci. Transl. Med. 2017, 9, eaai8700. [Google Scholar] [CrossRef]
- Michan, S.; Sinclair, D. Sirtuins in mammals: Insights into their biological function. Biochem. J. 2007, 404, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupis, W.; Pałyga, J.; Tomal, E.; Niewiadomska, E. The role of sirtuins in cellular homeostasis. J. Physiol. Biochem. 2016, 72, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Mostoslavsky, R. Fine tuning our cellular factories: Sirtuins in mitochondrial biology. Cell Metab. 2011, 13, 621–626. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The role of sirtuins in antioxidant and redox signaling. Antioxid. Redox Sign. 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Karaman, B.M.; Sippl, W.; Ntie-Kang, F. Natural products as modulators of Sirtuins. Molecules 2020, 25, 3287. [Google Scholar] [CrossRef]
- Schiedel, M.; Robaa, D.; Rumpf, T.; Sippl, W.; Jung, M. The current state of NAD+-dependent histone deacetylases (Sirtuins) as novel therapeutic targets. Med. Res. Rev. 2017, 38, 147–200. [Google Scholar] [CrossRef]
- Alqarni, M.H.; Foudah, A.I.; Muharram, M.M.; Labrou, N.E. The pleiotropic function of human sirtuins as modulators of metabolic pathways and viral infections. Cells 2021, 10, 460. [Google Scholar] [CrossRef]
- Gertz, M.; Nguyen, G.T.; Fischer, F.; Suenkel, B.; Schlicker, C.; Fränzel, B.; Tomaschewski, J.; Aladini, F.; Becker, C.; Wolters, D.; et al. A molecular mechanism for direct sirtuin activation by resveratrol. PLoS ONE 2012, 7, e49761. [Google Scholar] [CrossRef]
- Palmeira, C.M.; Teodoro, J.S.; Amorim, J.A.; Steegborn, C.; Sinclair, D.A.; Rolo, A.P. Mitohormesis and metabolic health: The interplay between ROS, cAMP and sirtuins. Free Radic. Biol. Med. 2019, 141, 483–491. [Google Scholar] [CrossRef]
- Moustafa, E.M.; Rashed, E.R.; Rashed, R.R.; Omar, N.N. Piceatannol promotes hepatic and renal AMPK/SIRT1/PGC-1α mitochondrial pathway in rats exposed to reserpine or gamma-radiation. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211016194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.P.; Zhao, Z.X.; Pang, X.F.; Yang, J.; Yu, H.X.; Zhang, Y.H.; Zhou, H.; Zhao, J.H. MiR-34a/sirtuin-1/foxo3a is involved in genistein protecting against ox-LDL-induced oxidative damage in HUVECs. Toxicol. Lett. 2017, 277, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Poljsak, B.; Milisav, I. NAD+ as the link between oxidative stress, inflammation, caloric restriction, exercise, DNA repair, longevity, and health span. Rejuvenation Res. 2016, 19, 406–415. [Google Scholar] [CrossRef]
- D’Arcy, M. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Verdin, E. NAD⁺ in aging, metabolism, and neurodegeneration. Science 2015, 350, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Cikla-Suzgun, P.; Kucukguzel, S.G. Recent advances in apoptosis: The role of hydrazones. Mini Rev. Med. Chem. 2019, 19, 1427–1442. [Google Scholar]
- Zhang, Y.M.; Zhang, W.R.; Tao, L.N.; Zhai, J.H.; Gao, H.; Song, Y.Q.; Qu, X.Y. Quercetin protected against isoniazide-induced HepG2 cell apoptosis by activating SIRT1/ERK pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22369. [Google Scholar] [CrossRef]
- Liang, Y.S.; Qi, W.T.; Guo, W.; Wang, C.L.; Hu, Z.B.; Li, A.K. Genistein and daidzein induce apoptosis of colon cancer cells by inhibiting the accumulation of lipid droplets. Food Nutr. Res. 2018, 2018, 62. [Google Scholar] [CrossRef]
- Kim, H.S.; Patel, K.; Muldoon-Jacobs, K.; Bisht, K.S.; Aykin-Burns, N.; Pennington, J.D.; van der Meer, R.; Nguyen, P.; Savage, J.; Owens, K.M.; et al. SIRT3 is a mitochondria-localized tumor suppressor required for maintenance of mitochondrial integrity and metabolism during stress. Cancer Cell 2010, 17, 41–52. [Google Scholar] [CrossRef]
- Rajendran, R.; Garva, R.; Krstic-Demonacos, M.; Demonacos, C. Sirtuins: Molecular traffic lights in the crossroad of oxidative stress, chromatin remodeling, and transcription. J. Biomed. Biotechnol. 2011, 2011, 368276. [Google Scholar] [CrossRef]
- Zhang, D.X.; Zhang, J.P.; Hu, J.Y.; Huang, Y.S. The potential regulatory roles of NAD+ and its metabolism in autophagy. Metabolism 2016, 65, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.H.; Jiang, T.L.; Wang, Y.; Guo, L. The role and mechanism of SIRT1 in resveratrol-regulated osteoblast autophagy in osteoporosis rats. Sci. Rep. 2019, 9, 18424. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ma, X.R.; Ouyang, T.X.; Chen, H.P.; Lin, J.; Liu, J.; Xiao, Y.; Yu, J.; Huang, Y.Y. SIRT1 reverses senescence via enhancing autophagy and attenuates oxidative stress-induced apoptosis through promoting p53 degradation. Int. J. Biol. Macromol. 2018, 117, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.X.; Jia, C.F.; Zhang, S.P.; Fan, G.J.; Li, Y.B.; Shan, P.P.; Sun, L.H.; Xiao, W.Z.; Li, L.; Zheng, Y.; et al. The REGγ proteasome regulates hepatic lipid metabolism through inhibition of autophagy. Cell Metab. 2013, 18, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H. Mechanisms and disease implications of sirtuin-mediated autophagic regulation. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef]
- Inoue, T.; Nakayama, Y.; Li, Y.; Matsumori, H.; Takahashi, H.; Kojima, H.; Wanibuchi, H.; Katoh, M.; Oshimura, M. SIRT2 knockdown increases basal autophagy and prevents postslippage death by abnormally prolonging the mitotic arrest that is induced by microtubule inhibitors. FEBS J. 2014, 281, 2623–2637. [Google Scholar] [CrossRef]
- Liu, G.; Park, S.H.; Imbesi, M.; Nathan, W.J.; Zou, X.; Zhu, Y.; Jiang, H.; Parisiadou, L.; Gius, D. Loss of NAD-dependent protein deacetylase sirtuin-2 alters mitochondrial protein acetylation and dysregulates mitophagy. Antioxid. Redox Signal. 2017, 26, 849–863. [Google Scholar] [CrossRef]
- Cho, C.S.; Lombard, D.B.; Lee, J.H. SIRT3 as a regulator of hepatic autophagy. Hepatology 2017, 66, 700–702. [Google Scholar] [CrossRef]
- Li, S.T.; Dou, X.B.; Ning, H.; Song, Q.; Wei, W.; Zhang, X.M.; Shen, C.; Li, J.X.; Sun, C.H.; Song, Z.Y. Sirtuin 3 acts as a negative regulator of autophagy dictating hepatocyte susceptibility to lipotoxicity. Hepatology 2017, 66, 936–952. [Google Scholar] [CrossRef]
- Lee, H.J.; Kang, M.G.; Cha, H.Y.; Kim, Y.M.; Lim, Y.; Yang, S.J. Effects of piceatannol and resveratrol on sirtuins and hepatic inflammation in high-fat diet-fed mice. J. Med. Food 2019, 22, 833–840. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD+ metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell Biol. 2020, 22, 119–141. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, J.G. Resveratrol inhibits proliferation and promotes apoptosis of neuroblastoma cells: Role of sirtuin 1. Neurochem. Res. 2011, 36, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Yao, H.; Caito, S.; Sundar, I.K.; Rahman, I. Redox regulation of SIRT1 inflammation and cellular senescence. Free Radic. Biol. Med. 2013, 61, 95–110. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Zhu, Y.; McGowan, S.J.; Angelini, L.; Fuhrmann-Stroissnigg, H.; Xu, M.; Ling, Y.Y.; Melos, K.I.; Pirtskhalava, T.; Inman, C.L.; et al. Fisetin is a senotherapeutic that extends health and lifespan. EBioMedicine 2018, 36, 18–28. [Google Scholar] [CrossRef]
- Anwar, T.; Khosla, S.; Ramakrishna, G. Increased expression of SIRT2 is a novel marker of cellular senescence and is dependent on wild type p53 status. Cell Cycle 2016, 15, 1883–1897. [Google Scholar] [CrossRef]
- Jing, E.; Emanuelli, B.; Hirschey, M.D.; Boucher, J.; Lee, K.Y.; Lombard, D.; Verdin, E.M.; Kahn, C.R. Sirtuin-3 (Sirt3) regulates skeletal muscle metabolism and insulin signaling via altered mitochondrial oxidation and reactive oxygen species production. Proc. Natl. Acad. Sci. USA 2011, 108, 14608–14613. [Google Scholar] [CrossRef]
- Zhong, L.; D’Urso, A.; Toiber, D.; Sebastian, C.; Henry, R.E.; Vadysirisack, D.D.; Guimaraes, A.; Marinelli, B.; Wikstrom, J.D.; Nir, T.; et al. The histone deacetylase Sirt6 regulates glucose homeostasis via Hif1alpha. Cell 2010, 140, 280–293. [Google Scholar] [CrossRef]
- Kanfi, Y.; Naiman, S.; Amir, G.; Peshti, V.; Zinman, G.; Nahum, L.; Bar-Joseph, Z.; Cohen, H.Y. The sirtuin SIRT6 regulates lifespan in male mice. Nature 2012, 483, 218–221. [Google Scholar] [CrossRef]
- Vazquez, B.N.; Thackray, J.K.; Simonet, N.G.; Kane-Goldsmith, N.; Martinez-Redondo, P.; Nguyen, T.; Bunting, S.; Vaquero, A.; Tischfield, J.A.; Serrano, L. SIRT7 promotes genome integrity and modulates non-homologous end joining DNA repair. EMBO J. 2016, 35, 1488–1503. [Google Scholar] [CrossRef]
- Vachharajani, V.T.; Liu, T.F.; Wang, X.F.; Hoth, J.J.; Yoza, B.K.; McCall, C.E. Sirtuins link inflammation and metabolism. J. Immunol. Res. 2016, 2016, 8167273. [Google Scholar] [CrossRef] [PubMed]
- Mendes, K.L.; Lelis, D.F.; Santos, S. Nuclear sirtuins and inflammatory signaling pathways. Cytokine Growth Factor Rev. 2017, 38, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Suuronen, T.; Ojala, J.; Kaarniranta, K.; Salminen, A. Antagonistic crosstalk between NF-κB and SIRT1 in the regulation of inflammation and metabolic disorders. Cell. Signal. 2013, 25, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.W.; Feng, J.; Zhang, J.Y.; Kang, X.; Qian, D. Quercetin modulates AMPK/SIRT1/NF κB signaling to inhibit inflammatory/oxidative stress responses in diabetic high fat diet induced atherosclerosis in the rat carotid artery. Exp. Ther. Med. 2020, 20, 280. [Google Scholar] [CrossRef]
- Hung, C.H.; Chan, S.H.; Chu, P.M.; Tsai, K.L. Quercetin is a potent anti-atherosclerotic compound by activation of SIRT1 signaling under oxLDL stimulation. Mol. Nutr. Food Res. 2015, 59, 1905–1917. [Google Scholar] [CrossRef]
- Chen, M.L.; Yi, L.; Jin, X.; Liang, X.Y.; Zhou, Y.; Zhang, T.; Xie, Q.; Zhou, X.; Chang, H.; Fu, Y.J.; et al. Resveratrol attenuates vascular endothelial inflammation by inducing autophagy through the cAMP signaling pathway. Autophagy 2013, 9, 2033–2045. [Google Scholar] [CrossRef]
- Koyuncu, E.; Budayeva, H.G.; Miteva, Y.V.; Ricci, D.P.; Silhavy, T.J.; Shenk, T.; Cristea, I.M. Sirtuins are evolutionarily conserved viral restriction factors. mBio 2014, 5, e2214–e2249. [Google Scholar] [CrossRef]
- Lin, S.C.; Ho, C.T.; Chuo, W.H.; Li, S.; Wang, T.T.; Lin, C.C. Effective inhibition of MERS-CoV infection by resveratrol. BMC Infect. Dis. 2017, 17, 144. [Google Scholar] [CrossRef]
- Saeedi-Boroujeni, A.; Mahmoudian-Sani, M.R. Anti-inflammatory potential of quercetin in COVID-19 treatment. J. Inflamm. 2021, 18, 3. [Google Scholar] [CrossRef]
- Okeke, E.S.; Ita, R.E.; Egong, E.J.; Udofia, L.E.; Mgbechidinma, C.L.; Akan, O.D. Metaproteomic insights into fermented fish and vegetable products and associated microbes. Food Chem. Mol. Sci. 2021, 3, 100045. [Google Scholar] [CrossRef]
- Rodgers, J.T.; Puigserver, P. Fasting-dependent glucose and lipid metabolic response through hepatic sirtuin 1. Proc. Natl. Acad. Sci. USA 2007, 104, 12861–12866. [Google Scholar] [CrossRef] [PubMed]
- Higashida, K.; Kim, S.H.; Jung, S.R.; Asaka, M.; Holloszy, J.O.; Han, D.H. Effects of resveratrol and SIRT1 on PGC-1α activity and mitochondrial biogenesis: A reevaluation. PLoS Biol. 2013, 11, e1001603. [Google Scholar] [CrossRef] [PubMed]
- Kalliora, C.; Kyriazis, I.D.; Oka, S.; Lieu, M.J.; Yue, Y.J.; Area-Gomez, E.; Pol, C.J.; Tian, Y.; Mizushima, W.; Chin, A.; et al. Dual PPARα/γ activation inhibits SIRT1-PGC-1α axis and causes cardiac dysfunction. JCI Insight 2019, 4, e129556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, E.; Gesta, S.; Kahn, C.R. SIRT2 regulates adipocyte differentiation through FOXO1 acetylation/deacetylation. Cell Metab. 2007, 6, 105–114. [Google Scholar] [CrossRef]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.; Ilkayeva, O.R.; et al. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 2010, 464, 121–125. [Google Scholar] [CrossRef]
- Kelly, G.S. A review of the sirtuin system, its clinical implications, and the potential role of dietary activators like resveratrol: Part 2. Altern. Med. Rev. 2010, 15, 313–338. [Google Scholar]
- Liang, F.; Kume, S.; Koya, D. SIRT1 and insulin resistance. Nat. Rev. Endocrinol. 2009, 5, 367–373. [Google Scholar] [CrossRef]
- Rodgers, J.T.; Lerin, C.; Gerhart-Hines, Z.; Puigserver, P. Metabolic adaptations through the PGC-1 alpha and SIRT1 pathways. FEBS Lett. 2008, 582, 46–53. [Google Scholar] [CrossRef]
- Zhu, X.F.; Yang, J.Y.; Zhu, W.J.; Yin, X.X.; Yang, B.B.; Wei, Y.H.; Guo, X.F. Combination of berberine with resveratrol improves the lipid-lowering efficacy. Int. J. Mol. Sci. 2018, 19, 3903. [Google Scholar] [CrossRef]
- Danaei, G.; Hoorn, S.V.; Lopez, A.D.; Murray, C.J.L.; Ezzati, M. Comparative risk assessment collaborating group (Cancers). Causes of cancer in the world: Comparative risk assessment of nine behavioural and environmental risk factors. Lancet 2005, 366, 1784–1793. [Google Scholar] [CrossRef]
- Bosch-Presegué, L.; Vaquero, A. The dual role of sirtuins in cancer. Genes Cancer 2011, 2, 648–662. [Google Scholar] [CrossRef]
- George, J.; Ahmad, N. Mitochondrial sirtuins in cancer: Emerging roles and therapeutic potential. Cancer Res. 2016, 76, 2500–2506. [Google Scholar] [CrossRef] [PubMed]
- Chao, S.C.; Chen, Y.J.; Huang, K.H.; Kuo, K.L.; Yang, T.H.; Huang, K.Y.; Wang, C.C.; Tang, C.H.; Yang, R.S.; Liu, S.H. Induction of sirtuin-1 signaling by resveratrol induces human chondrosarcoma cell apoptosis and exhibits antitumor activity. Sci. Rep. 2017, 7, 3180. [Google Scholar] [CrossRef] [PubMed]
- González-Vallinas, M.; González-Castejón, M.; Rodríguez-Casado, A.; Ramírez de Molina, A. Dietary phytochemicals in cancer prevention and therapy: A complementary approach with promising perspectives. Nutr. Rev. 2013, 71, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.S.; Prasad, S.K.; Shivamallu, C.; Prasad, K.S.; Syed, A.; Reddy, P.; Cull, C.A.; Amachawadi, R.G. Genistein: A potent anti-breast cancer agent. Curr. Issues Mol. Biol. 2021, 43, 1502–1517. [Google Scholar] [CrossRef]
- Thomas, R.; Butler, E.; Macchi, F.; Williams, M. Phytochemicals in cancer prevention and management. Brit. J. Med. Pract. 2015, 8, a815. [Google Scholar]
- Connell, N.J.; Houtkooper, R.H.; Schrauwen, P. NAD+ metabolism as a target for metabolic health: Have we found the silver bullet? Diabetologia 2019, 62, 888–899. [Google Scholar] [CrossRef]
- Agerholm, M.; Dall, M.; Jensen, B.; Prats, C.; Madsen, S.; Basse, A.L.; Graae, A.S.; Risis, S.; Goldenbaum, J.; Quistorff, B.; et al. Perturbations of NAD salvage systems impact mitochondrial function and energy homeostasis in mouse myoblasts and intact skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2017, 314, E377–E395. [Google Scholar] [CrossRef]
- Griffiths, H.B.S.; Williams, C.; King, S.J.; Allison, S.J. Nicotinamide adenine dinucleotide (NAD+): Essential redox metabolite, co-substrate and an anti-cancer and anti-ageing therapeutic target. Biochem. Soc. Trans. 2020, 48, 733–744. [Google Scholar] [CrossRef]
- Libert, S.; Guarente, L. Metabolic and neuropsychiatric effects of calorie restriction and sirtuins. Annu. Rev. Physiol. 2013, 75, 669–684. [Google Scholar] [CrossRef]
- Cortes-Martin, A.; Selma, M.V.; Tomás-Barberán, F.A.; González-Sarrías, A.; Espín, J.C. Where to look into the puzzle of polyphenols and health? The postbiotics and gut microbiota associated with human metabotypes. Mol. Nutr. Food Res. 2020, 64, 1900952. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Sinclair, D.A.; Ellis, J.L.; Steegborn, C. Sirtuin activators and inhibitors: Promises, achievements, and challenges. Pharmacol. Ther. 2018, 188, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Crous-Bou, M.; Molinuevo, J.L.; Sala-Vila, A. Plant-rich dietary patterns, plant foods and nutrients, and telomere length. Adv. Nutr. 2019, 10, S296–S303. [Google Scholar] [CrossRef]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef]
- Hold, G.L. Western lifestyle: A ‘master’ manipulator of the intestinal microbiota? Gut 2014, 63, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Mueller, N.T.; Whyatt, R.; Hoepner, L.; Oberfield, S.; Dominguez-Bello, M.G.; Widen, E.M.; Hassoun, A.; Perera, F.; Rundle, A. Prenatal exposure to antibiotics, cesarean section and risk of childhood obesity. Int. J. Obes. 2014, 39, 665–670. [Google Scholar] [CrossRef]
- Giuliani, C.; Marzorati, M.; Innocenti, M.; Vilchez-Vargas, R.; Vital, M.; Pieper, D.H.; Van de Wiele, T.; Mulinacci, N. Dietary supplement based on stilbenes: A focus on gut microbial metabolism by the in vitro simulator M-SHIME®. Food Funct. 2016, 7, 4564–4575. [Google Scholar] [CrossRef]
- van Dorsten, F.A.; Grün, C.H.; van Velzen, E.J.; Jacobs, D.M.; Draijer, R.; van Duynhoven, J.P. The metabolic fate of red wine and grape juice polyphenols in humans assessed by metabolomics. Mol. Nutr. Food Res. 2010, 54, 897–908. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Chalet, C.; Rubbens, J.; Tack, J.; Duchateau, G.S.; Augustijns, P. Intestinal disposition of quercetin and its phase-II metabolites after oral administration in healthy volunteers. J. Pharm. Pharmacol. 2018, 70, 1002–1008. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Demchuk, O.M. New perspectives for fisetin. Front. Chem. 2019, 7, 697. [Google Scholar] [CrossRef] [PubMed]
- Banik, K.; Ranaware, A.M.; Harsha, C.; Nitesh, T.; Girisa, S.; Deshpande, V.; Fan, L.; Nalawade, S.P.; Sethi, G.; Kunnumakkara, A.B. Piceatannol: A natural stilbene for the prevention and treatment of cancer. Pharmacol. Res. 2020, 153, 104635. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Zhang, L.M.; Liao, P.F.; Xiao, Z.L.; Zhang, F.; Sindaye, D.; Xin, Z.Q.; Tan, C.Q.; Deng, J.P.; Yin, Y.L.; et al. Impact of gallic acid on gut health: Focus on the gut microbiome, immune response, and mechanisms of action. Front. Immunol. 2020, 11, 580208. [Google Scholar] [CrossRef]
- Kane, A.E.; Sinclair, D.A. Sirtuins and NAD+ in the development and treatment of metabolic and cardiovascular diseases. Circ. Res. 2018, 123, 868–885. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2019, 60, 626–659. [Google Scholar] [CrossRef]
- Chitchumroonchokchai, C.; Riedl, K.M.; Suksumrarn, S.; Clinton, S.K.; Kinghorn, A.D.; Failla, M.L. Xanthones in mangosteen juice are absorbed and partially conjugated by healthy adults. J. Nutr. 2012, 142, 675–680. [Google Scholar] [CrossRef]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the human gut microbiome and host metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef]
- Schwer, B.; Verdin, E. Conserved metabolic regulatory functions of Sirtuins. Cell Metab. 2008, 7, 104–112. [Google Scholar] [CrossRef]
- Cueva, C.; Silva, M.; Pinillos, I.; Bartolomé, B.; Moreno-Arribas, M.V. Interplay between dietary polyphenols and oral and gut microbiota in the development of colorectal cancer. Nutrients 2020, 12, 625. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of intestinal microbiota in the bioavailability and physiological functions of dietary polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [PubMed]
- Kemperman, R.A.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel approaches for analyzing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef]
- Koudoufio, M.; Desjardins, Y.; Feldman, F.; Spahis, S.; Delvin, E.; Levy, E. Insight into polyphenol and gut microbiota crosstalk: Are their metabolites the key to understand protective effects against metabolic disorders? Antioxidants 2020, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Clifford, M.N. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of polysaccharides in food, digestion, and health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef]
- Zhao, J.F.; Zhang, X.Y.; Liu, H.B.; Brown, M.A.; Qiao, S.Y. Dietary protein and gut microbiota composition and function. Curr. Protein Pept. Sci. 2019, 20, 145–154. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type (Class) | Localization (Mol. Weight) | Tissue Expression | Enzyme Activity | Substrates | Functions | Activators | Inhibitors |
|---|---|---|---|---|---|---|---|
| SIRT1 (I) | Nuclear; cytosolic. (81.7 kDa) | Brain; Skeletal muscle; Heart; Kidney; Uterus. | NAD+-dependent deacetylases catalyze the deacetylation of histones and nonhistone proteins. | Acetyl-CoA synthetase 1;Akt; Androgene receptor; APE1; Atg 5; Atg 7; Atg 8; BCL6; BMAL1; c-MYC; cortactin; CRABPII; CRTC2; DNMT1; eNOS; FOXA2; FOXO family; FXR; HIF1α; HIF2α; Histone H1K26; Histone H3K9; Histone H4K16; Histone K14; Histone K56; HIV Tat; HMGB1; HMGCS1; hMOF; HSF1; IRF-1; KAP1; KDAC1; Ku70; LKB1; LXR; MEF2; NBS1; NF-κB(p65); Notch1; p300; p53; p73; PARP1; PCAF; PER2; PGAM1; PGC1α; PML; PPAR-γ; RFX-5; SATB1; Smad7; SREBP-1c; SUV39H1; TDG; TFAM; TIP60; TopBP1; TORC1; UCP2; WRN; XPA. | Apoptosis; Cell cycle; Cell migration; Cell survival; Circadian control; Inflammation; Insulin signaling; Lifespan regulation; Metabolism regulation; Neuroprotection; Oxidative stress response; Mitohormesis; Tumor suppression; Viral transcription. | Resveratrol; Piceatannol; Quercetin; Fisetin; Daidzein; Genistein; Berberine; Flavonoid mulberrin; The xanthone gartanin; The alkaloids quinidine and quinine. | Tanikolide dimer; Chalcones; Biochalcones. |
| SIRT2 (I) | Nuclear; cytosolic. (41.5 kDa) | Brain. | NAD+-dependent HDAC and ADP-ribosyl transferase activities. | Akt; ALDH1A1; APCCDH1; ATRIP; BubR1; CDC20; CDK9; FOXO1; FOXO3a; G6PD; H4K16; HIF1α; Histone H3K18; Histone H3K56; Histone H4K16; K-RAS; NF-κB (p65); p300; p53; Par-3; PEPCK1; PGAM; TUG; α-tubulin; β-secretase 1. | Adipocyte differentiation; Autophagy; Bacterial infections; Cell cycle regulation; Energy homeostasis Fibrosis; Genome stability; Immune response; Longevity; Myelination; Neurodegeneration. | Resveratrol. | Tanikolide dimer. |
| SIRT3 (I) | Mitochondrial; Nuclear. (43.6 kDa) | Brain; Heart; Liver; Kidney; Brown adipose tissue. | NAD+-dependent deacetylases catalyze the deacetylation of histones and nonhistone proteins. | Acetyl-CoA synthetase 2; Aconitase 2; ALDH2; ATP-synthase F1; Complex 1; Cyclophilin D; FOXO3a; GDH; GOT2; GSK3β; Histone H4K16; HMGCS2; Hsp10; Long-chain acyl-CoA dehydrogenase; HMGCS; Isocitrate dehydrogenase 2; Ku70; LCAD; LKB1; MDH; Mitochondrial ribosomal protein L10; MnSOD; OGG1; OPA1; OTC; PDH; PGC-1α; SDH; Skp2; SOD2; UCP-1; VLCAD. | Fatty acid oxidation; Fibrosis; Longevity; Metabolism regulation; Oxidative phosphorylation; Oxidative stress TCA cycle; Thermogenesis; Urea cycle. | Resveratrol; Trans-(−)-ε-viniferin; Piceatannol. | - |
| SIRT4 (II) | Mitochondrial (35.2 kDa) | Brain; Liver; Kidney; Heart; Pancreatic β-cells. | NAD+-dependent HDAC and ADP-ribosyl transferase activities. | ANT2/3; GDH; Hsp60; MCD; Pyruvate dehydrogenase complex; Stress-70. | Fatty acid oxidation; Insulin secretion; Metabolism regulation; TCA cycle; Tumor suppression. | - | |
| SIRT5 (III) | Mitochondrial; Nuclear (33.9 kDa) | Brain; Heart; Muscle; Testis; Lymphoblast. | NAD+-dependent deacetylases catalyze the deacetylation of histones; Desuccinylase; Demalonylase; Deglutarylase. | Cytochrome c; Carbamoyl phosphate; GAPDH; HMGCS2; Hsp70; IDH; PML; Prx-1; SOD1; Synthetase 1; UOX; VLCAD. | Apoptosis; Fatty acid oxidation; Ketone body synthesis; Oxidative stress; Urea cycle. | Resveratrol; Piceatannol. | - |
| SIRT6 (IV) | Nuclear (39.1 kDa) | Brain; Heart; Muscles; Ovaries; Bone cells. | NAD+-dependent ADP ribosyltransferase (ART) mediating mitochondrial protein ribosylation; Demyristoylase; Depalmitoylase. | GCN5; Histone H3K9; Histone H3K56; H3k18ac; TNF-α; KAP1; NF-κB; p70; PAPR1. | Apoptosis; DNA repair; Genome stability; Longevity; Protein secretion. | Anthocyanidins; Gallic acid derivatives. | Catechins; Epicatechins; Phytoestrogens. |
| SIRT7 (IV) | Nucleolus; Cytoplasm (44.9 kDa) | Peripheral blood cells; CD33+ myeloid bone marrow precursor cells. | NAD+ dependent deacetylase; Desuccinylase; Regulates the RNA polymerase I (Pol I) transcriptional machinery. | DNA-PK; GABPβ1; Histone H3K122; Histone H3K18; H3K36ac; p53; PAF53; RNA polymerase I; U3–55k. | Cell cycle regulation; Genome stability; Regulation of rDNA transcription; Tumor promotion. | - |
| Polyphenol | Diet Source | Postbiotics | Health Proffering Mechanisms | Microbes Involved | References | |
|---|---|---|---|---|---|---|
| 1 | Resveratrol (non-flavonoid stilbene) | Red wine, peanuts, red grapes, and selected teas. | Dihydroresveratrol, Lunularin, 3,4′-dihydroxy-trans-stilbene. | AMPK activity ↑→lipid lowering TG plasma ↓ Carcinogenesis ↓ Bacteroidetes↑ SCFAs ↑ | Bacteroidetes, Actinobacteria, Verrucomicrobia, Cyanobacteria, Slackia equolifaciens, and Adlercreutzia equolifaciens. | [88]. |
| 2 | Trans-(−)-ε-viniferin (Stilbene) | Grapes and wines. | ε-viniferin mono-sulfate, ε-viniferin mono-glucuronide. | Anti-fungal ↑ Intestinal sodium-dependent glucose uptake via the SGLT1↓ | Enterobacteriales. | [89]. |
| 3 | Quercetin (dietary flavonols) | Tea, red wine, berries, apples, tomatoes, beans, and onions. | Quercetin-3-glucuronide, Quercetin-7-glucuronide, 3,4-dihydroxyphenylacetic acid, 3-(3-hydroxyphenyl) propionic acid, 3,4-dihydroxybenzoic acid, 4-hydroxybenzoic acid. | Cellular barrier ↓ antioxidative pathway ↑. serum IL-6↓ Synthesis of polyamines ↓ Anti-inflammation ↑ Bacteroidetes ↑ SCFAs ↑. | Eubacterium ramulus, Eubacterium oxidoreducens, Clostridium orbiscindens, and Butyrovibrio spp. | [90]. |
| 4 | Fisetin (dietary flavonol) | Strawberries, apples, persimmons, grapes, peach, lotus root, cucumber, teas, onions, kiwi, and kale. | Glucuronidated fisetin, Geraldol (3,4′,7-trihydroxy-3′-methoxyflavone), and Glucuronidated geraldol. | Anti-pathogenic ↑ Butyrate production ↑ Apoptosis ↑ MAPK/NF-κB ↓. | Lachnospiraceae. | [91]. |
| 5 | Piceatannol (Stilbene) | Grapes, passionfruit, white tea, wines, Japanese knotweed, Asian legumes, and Korean rhubarb. | Piceatannol disulfate, piceatannol monosulfate-1, and piceatannol monosulfate-2. | Cell survival or proliferation ↓ Cell cytotoxicity ↑ Reactive oxygen species level ↓ Autophagy ↑ Cell cycle proteins ↑. | Lactobacillus spp, Lachnospiraceae, and Bacillus megaterium CYP102A1. | [89]. |
| 6 | Daidzein (isoflavones) | Soybeans, legumes, whole grains, berries, and nuts. | Dihydrodaidzein; can further be converted to S-equol. | Hormone-dependent diseases ↓ Antioxidation ↑ Anti-cancer ↑. | Bifidobacteria sp. (B. breve and B. longum), Lactococcus strain 20–92, Eggerthella sp. Julong 732, and Eubacterium limosum. | [92]. |
| 7 | Anthocyanidin | Berries, currants, grapes, colored leafy vegetables, grains, roots, and tubers. | 4-hydroxybenzoic acid, protocatechuic acid, gallic acid, vanillic acid, syringic acid, catechol, pyrogallol, resorcinol, tyrosol, 3-(3′-hydroxyphenyl) propionic acid, dihydrocaffeic acid, and 3-(4′-hydroxyphenyl) lactic acid. | Modulation of gut microbiota → Anti-pathogenicity ↑ SCFAs ↑ Vitamin production ↑ Anti-inflammatory ↑ Gut microbiota metabolites ↑→ macrophage reverse cholesterol transport ↑ Atherosclerotic lesion ↓. | Bifidobaterium spp., Lactobacillus spp., Eubacterium ramulus, and Clostridium saccbarogumia. | [85,93]. |
| 8 | Gallic acid derivatives | Grapes, gallnuts, pomegranates, and tea leaves. | Pyrogallol-1-O-glucuronide, 4-OMeGA, 4-OMeGA-3-O-sulfate, pyrogallol-O-sulfate, deoxypyrogallol-O-sulfate, and O-methylpyrogallol-O-sulfate. | Gut microbial modulation, Helicobacter pylori ↓ Anti-pathogenic ↑ Anti-cancer ↑ anti-inflammatory ↑ SCFAs ↑. | Pseudomonas and Atopobium spp. | [94]. |
| 9 | Genistein (isoflavones) | Soy, raisins, currants, prunes, mango, passionfruit, quinoa seeds, and peanuts. | p-ethyl phenol and 4-hydroxy-phenyl-2-propionic acid. | Binding to estrogenic receptors → estrogenic/anti-estrogenic activities. Carcinogenesis ↓. | Lactobacillum, Lactoccocus, Enterococcus, Bifidobacterium, and Bacteroides. | [90]. |
| 10 | Xanthone gartanin | Mangosteen. | Alpha- and γ-mangostin. | Apoptosis ↑ Akt, MAPK, NF-κB pathways ↓ Anti-microbial ↑. | Enterobacteriaceae and Enterococcaceae. | [9]. |
| 11 | Quinine | Cinchona tree bark. | 3-hydroxyquinine. | Anti-malarial ↑ T2R family receptors ↑. | Subdoligranulum spp., Akkermansia muciniphila, Roseburia inulinivorans, Methanobrevibacter smithii, and Roseburia intestinalis. | [95]. |
| 12 | Catechins | Acacia catechu L., Camellia sinensis, red wine, and chocolate. | UDP-glucuronosyltransferases (UGTs); sulphotransferases (SULTs); and catechol-O-methyltransferase (COMT). | Anti-cancer ↑ Antioxidation ↑ Anti-inflammatory activities ↑ Immune activity ↑ Receptor tyrosine kinase ↓. | Aspergillus, Penicillium, Rhizopus, Mucor, Yeasts, and Bacterium. | [96]. |
| 13 | Epicatechins | Apples, blackberries, broad beans, cherries, grapes, pears, raspberries, dark chocolates, cocoa, and tea leaves. | 1-(3′,4′-dihydroxyphenyl)-3-(2″,4″,6″-dihydroxyphenyl)-2-propanol (3,4-diHPP-2-ol) and 5-(3′,4′-dihydroxyphenyl)-γ-valerolactone (3,4-diHPV) | Antioxidant activities ↑ Modulates NO and ROS→improves arterial vessels endothelial functions. | Eubacterium SDG-2, Lactobacillus plantarum, Eggerthella lenta, and Adlercreutzia equolifaciens | [97]. |
| 14 | Phytoestrogens | Flax seeds, grapes, soybeans, kidney beans, apples, cabbage, spinach, hops, garlic, onion, wine, and tea. | Equol, urolithins, and enterolignans. | Cardiovascular diseases ↓ Diabetes ↓ Breast cancer ↓ Osteoporosis ↓ Anti-inflammation ↑. | Bacteroides, Clostridium strains, Lactobacillus strains, Gordonibacter urolithinfaciens, Gordonibacter pamelaeae DSM 19378(T), Bifidobacterium, Lactobacillus, Enterococcus faecalis, and Streptococcus bovis. | [98]. |
| 15 | Tanikolide dimer | Lyngbya majuscule: a Madagascar marine Cyanobacterium. | - | Anti-fungal activity against Candida albicans Anti-bacterial activity against Mycobacterium smegmatis and Streptococcus pyogenes Anti-cancer ↑. | - | [9]. |
| 16 | Chalcones | Citruses, Ochna sp., apples, tomatoes, shallots, bean sprouts, and potatoes. | Dihydrochalcone | Anti-cancer ↑ Antioxidative ↑ Anti-microbial ↑ anti-inflammatory ↑. | Eubacterium ramulus. | [99]. |
| 17 | Biochalcones | Ochna sp. and Rhus pyroides. | - | Anti-protozoal ↑ Anti-viral ↑. | - | [97]. |
| 18 | Berberine | Chinese herb (Rhizoma coptidis). | Raisanberine and CPU86017. | Anti-viral ↑ Anti-cancer ↑ Antioxidative ↑; Anti-inflammation↑ Cardioprotective effects ↑. | Clostridium hiranonis, C. scindens, and C. hylemonae. | [99]. |
| 19 | Undigested polysaccharides/ dietary fiber | SCFAs. | Acetate, propionate, and butyrate. | Colonocytes and epithelia cells Apoptosis ↑ Histone deacetylases ↑ →FFAR2 and FFAR3 →intestinal gluconeogenesis ↑ → satiety ↑ Glucose → adiposity ↓ Lipogenesis ↑ Gut microbial eubiosis↑. | Bifidobacterium longum, Eubacterium hallii, Lachnospiraceae, Faecalibacterium prausnitzii, Negativicutes, Clostridium species, Roseburia inulinivorans, and Ruminococcus obeum. | [100]. |
| 20 | Undigested polysaccharides/dietary fiber | Gases. | Mercaptans, sulphated mucins, and hydrogen sulfide. | Microbial redox reactions ↑ Anaerobic fermentation ↑ Lubricate gastrointestinal tracts ↑ Cell signaling molecules ↑. | Bacteroides, Clostridium, and Desulfovibrio. | [101]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akan, O.D.; Qin, D.; Guo, T.; Lin, Q.; Luo, F. Sirtfoods: New Concept Foods, Functions, and Mechanisms. Foods 2022, 11, 2955. https://doi.org/10.3390/foods11192955
Akan OD, Qin D, Guo T, Lin Q, Luo F. Sirtfoods: New Concept Foods, Functions, and Mechanisms. Foods. 2022; 11(19):2955. https://doi.org/10.3390/foods11192955
Chicago/Turabian StyleAkan, Otobong Donald, Dandan Qin, Tianyi Guo, Qinlu Lin, and Feijun Luo. 2022. "Sirtfoods: New Concept Foods, Functions, and Mechanisms" Foods 11, no. 19: 2955. https://doi.org/10.3390/foods11192955
APA StyleAkan, O. D., Qin, D., Guo, T., Lin, Q., & Luo, F. (2022). Sirtfoods: New Concept Foods, Functions, and Mechanisms. Foods, 11(19), 2955. https://doi.org/10.3390/foods11192955