

Phosphorylated Trehalose Suppresses the Denaturation of Myofibrillar Proteins in Peeled Shrimp (Litopenaeus vannamei) during Long-Term Frozen Storage


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Phosphorylated Trehalose
2.3. Shrimp Sample and Treatments
2.4. Determination of Water-Holding Capacity (WHC)
2.5. Extraction of Myofibrillar Proteins (MP)
2.6. Determination of Ca2+-ATPase Activity
2.7. Determination of Total Sulfhydryl (T-SH) Content
2.8. Determination of Surface Hydrophobicity
2.9. Atomic Force Microscope (AFM) Analysis
2.10. Hematoxylin and Eosin (H&E) Staining Analysis
2.11. Differential Scanning Calorimetry (DSC) Analysis
2.12. Data Analysis
3. Results and Discussion
3.1. Water-Holding Capacity (WHC) Analysis
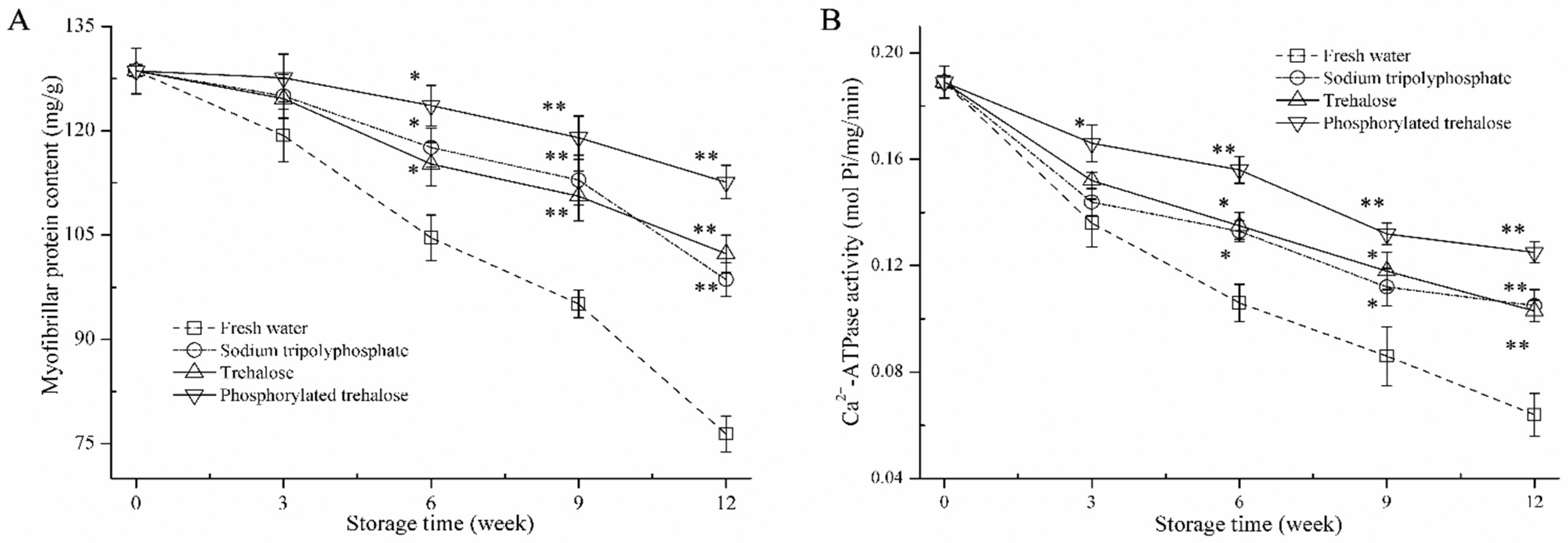
3.2. Myofibrillar Protein (MP) Content and Ca2+-ATPase Activity Analysis
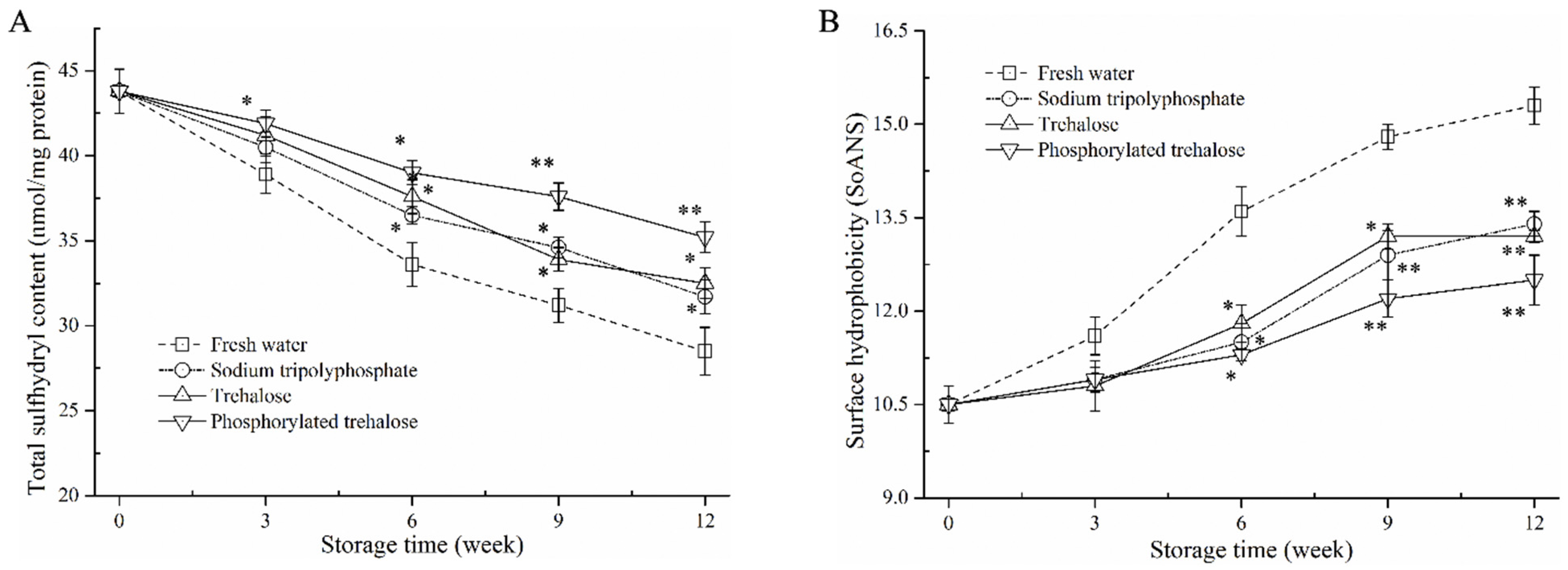
3.3. Total Sulfhydryl (T-SH) Content and Surface Hydrophobicity Analysis
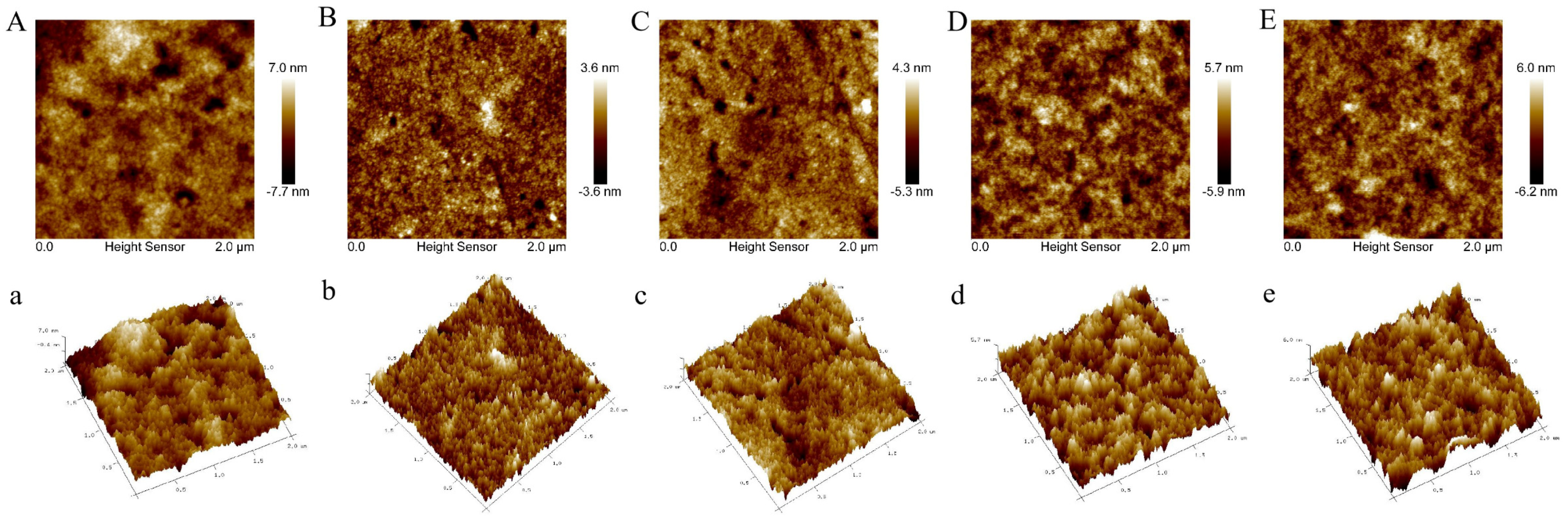
3.4. Atomic Force Microscope (AFM) Analysis
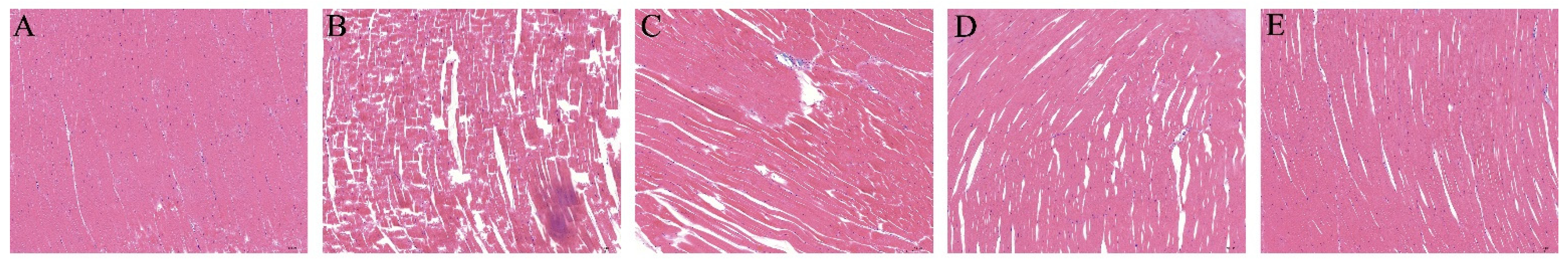
3.5. Hematoxylin and Eosin (H&E) Staining Analysis
3.6. Thermal Stability Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, B.; Yao, H.; Qi, H.; Ying, X.G. Cryoprotective characteristics of different sugar alcohols on peeled Pacific white shrimp (Litopenaeus vannamei) during frozen storage and their possible mechanisms of action. Int. J. Food Prop. 2020, 23, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.E.d.S.; Gonçalves, A.A. The effect of different food grade additives on the quality of Pacific white shrimp (Litopenaeus vannamei) after two freeze-thaw cycles. LWT 2019, 113, 108301. [Google Scholar] [CrossRef]
- Thangavelu, K.P.; Kerry, J.P.; Tiwari, B.K.; McDonnell, C.K. Novel processing technologies and ingredient strategies for the reduction of phosphate additives in processed meat. Trends Food Sci. Technol. 2019, 94, 43–53. [Google Scholar] [CrossRef]
- Jain, N.K.; Roy, I. Effect of trehalose on protein structure. Protein Sci. 2009, 18, 24–36. [Google Scholar] [CrossRef]
- Zhang, B.; Yao, H.; Qi, H.; Zhang, X.L. Trehalose and alginate oligosaccharides increase the stability of muscle proteins in frozen shrimp (Litopenaeus vannamei). Food Funct. 2020, 11, 1270–1278. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wu, Y.J.; Sun, P.L.; Zhang, F.M.; Linhardt, R.J.; Zhang, A.Q. Chemically modified polysaccharides: Synthesis, characterization, structure activity relationships of action. Int. J. Biol. Macromol. 2019, 132, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Su, N.; Huang, Q.; Zhang, Q.; Wang, Y.; Li, J.; Ye, M. Phosphorylation and antiaging activity of polysaccharide from Trichosanthes peel. J. Food Drug Anal. 2017, 25, 976–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.Y.; Ji, Y.H.; Liao, A.M.; Huang, J.H.; Thakur, K.; Li, X.L.; Hu, F.; Zhang, J.G.; Wei, Z.J. Effects of sulfated, phosphorylated and carboxymethylated modifications on the antioxidant activities in-vitro of polysaccharides sequentially extracted from Amana edulis. Int. J. Biol. Macromol. 2020, 146, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xu, D.; Xia, N.; Zhao, X.; Kong, F.; Wang, S.; Fatehi, P. Preparation and application of phosphorylated xylan as a flocculant for cationic ethyl violet dye. Polymers 2018, 10, 317. [Google Scholar] [CrossRef] [Green Version]
- Teng, Q.; Hu, X.F.; Luo, F.; Cheng, C.; Ge, X.Y.; Yang, M.Y.; Liu, L.M. Influences of introducing frogs in the paddy fields on soil properties and rice growth. J. Soils Sediments 2016, 16, 51–61. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, H.X.; Yang, H.C.; Xiang, X.W.; Li, H.B.; Deng, S.G. Cryoprotective roles of trehalose and alginate oligosaccharides during frozen storage of peeled shrimp (Litopenaeus vannamei). Food Chem. 2017, 228, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, H.; Tang, H.; Hao, G.; Zhang, Y.; Deng, S. Insights into cryoprotective roles of carrageenan oligosaccharides in peeled whiteleg shrimp (Litopenaeus vannamei) during frozen storage. J. Agric. Food Chem. 2017, 65, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.G.; Wu, Q.J.; Shui, S.S.; Zhang, B.; Benjakul, S. Insights into the similarities and differences of whiteleg shrimp pre-soaked with sodium tripolyphosphate and sodium trimetaphosphate during frozen storage. Food Chem. 2021, 348, 129134. [Google Scholar] [CrossRef]
- Zuo, H.; Han, L.; Yu, Q.; Niu, K.; Zhao, S.; Shi, H. Proteome changes on water-holding capacity of yak longissimus lumborum during postmortem aging. Meat Sci. 2016, 121, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Sun, D.W. Novel methods for rapid freezing and thawing of foods—A review. J. Food Eng. 2002, 54, 175–182. [Google Scholar] [CrossRef]
- Reza, M.S.; Bapary, M.A.; Ahasan, C.T.; Islam, M.N.; Kamal, M. Shelf life of several marine fish species of Bangladesh during ice storage. Int. J. Food Sci. Technol. 2009, 44, 1485–1494. [Google Scholar] [CrossRef]
- Shui, S.S.; Qi, H.; Shaimaa, H.; Aubourg, S.P.; Zhang, B. Kappa-carrageenan and its oligosaccharides maintain the physicochemical properties of myofibrillar proteins in shrimp mud (Xia-Hua) during frozen storage. J. Food Sci. 2021, 86, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiong, Q.; Lai, X.; Li, X.; Wan, M.; Zhang, J.; Yan, Y.; Cao, M.; Lu, L.; Guan, J. Molecular modification of polysaccharides and resulting bioactivities. Compr. Rev. Food Sci. Food Saf. 2016, 15, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Hou, R.; Zeng, X.A. Synergistic effects of ultrasound and soluble soybean polysaccharide on frozen surimi from grass carp. J. Food Eng. 2018, 240, 1–8. [Google Scholar] [CrossRef]
- Jain, N.K.; Roy, I. Trehalose and protein stability. Curr. Protoc. Protein Sci. 2010, 59, 4.9.1–4.9.12. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, L.; Chen, C.; Xiong, G.; Hu, Y.; Qiao, Y.; Wu, W.; Li, X.; Wang, J.; Liao, L. Antioxidant capacity of fermented soybeans and their protective effect on protein oxidation in largemouth bass (Micropterus salmoides) during repeated freezing-thawing (FT) treatments. LWT-Food Sci. Technol. 2018, 91, 213–221. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, L.; Li, Q.; Luo, Y. Comparison of gel properties and biochemical characteristics of myofibrillar protein from bighead carp (Aristichthys nobilis) affected by frozen storage and a hydroxyl radical-generation oxidizing system. Food Chem. 2017, 223, 96–103. [Google Scholar] [CrossRef]
- Nikoo, M.; Benjakul, S.; Rahmanifarah, K. Hydrolysates from marine sources as cryoprotective substances in seafoods and seafood products. Trends Food Sci. Technol. 2016, 57, 40–51. [Google Scholar] [CrossRef]
- Zhang, B.; Fang, C.; Hao, G.; Zhang, Y. Effect of kappa-carrageenan oligosaccharides on myofibrillar protein oxidation in peeled shrimp (Litopenaeus vannamei) during long-term frozen storage. Food Chem. 2018, 245, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhou, Y.; Zhao, L.; Chen, L.; He, Y.; Yang, H. Vacuum impregnation of fish gelatin combined with grape seed extract inhibits protein oxidation and degradation of chilled tilapia fillets. Food Chem. 2019, 294, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Nikoo, M.; Benjakul, S. Potential application of seafood-derived peptides as bifunctional ingredients, antioxidant-cryoprotectant: A review. J. Funct. Foods 2015, 19, 753–764. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, J.L.; Chen, S.J.; Zhang, X.L.; Wei, W.Y. Influence of trehalose and alginate oligosaccharides on ice crystal growth and recrystallization in whiteleg shrimp (Litopenaeus vannamei) during frozen storage with temperature fluctuations. Int. J. Refrig. 2019, 99, 176–185. [Google Scholar] [CrossRef]
- Zhang, B.; Qi, X.E.; Mao, J.L.; Ying, X.G. Trehalose and alginate oligosaccharides affect the stability of myosin in whiteleg shrimp (Litopenaeus vannamei): The water-replacement mechanism confirmed by molecular dynamic simulation. LWT-Food Sci. Technol. 2020, 127, 109393. [Google Scholar] [CrossRef]
- Zhang, B.; Cao, H.J.; Lin, H.M.; Deng, S.G.; Wu, H. Insights into ice-growth inhibition by trehalose and alginate oligosaccharides in peeled Pacific white shrimp (Litopenaeus vannamei) during frozen storage. Food Chem. 2019, 278, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.k.; Zhang, B.; Deng, S.g.; Xie, C. Comparison of the cryoprotective effects of trehalose, alginate, and its oligosaccharides on peeled shrimp (Litopenaeus vannamei) during frozen storage. J. Food Sci. 2015, 80, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kong, B.; Li, F.; Liu, Q.; Zhang, H.; Xia, X. Changes in the thermal stability and structure of protein from porcine longissimus dorsi induced by different thawing methods. Food Chem. 2020, 316, 126375. [Google Scholar] [CrossRef] [PubMed]
- Harnkarnsujarit, N.; Charoenrein, S. Influence of collapsed structure on stability of β-carotene in freeze-dried mangoes. Food Res. Int. 2011, 44, 3188–3194. [Google Scholar] [CrossRef]
- Harnkarnsujarit, N.; Kawai, K.; Suzuki, T. Effects of freezing temperature and water activity on microstructure, color, and protein conformation of freeze-dried bluefin tuna (Thunnus orientalis). Food Bioprocess Technol. 2015, 8, 916–925. [Google Scholar] [CrossRef]
- Hu, Z.; Qiu, L.; Sun, Y.; Xiong, H.; Ogra, Y. Improvement of the solubility and emulsifying properties of rice bran protein by phosphorylation with sodium trimetaphosphate. Food Hydrocoll. 2019, 96, 288–299. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WHC | Storage Time (Week) | Fresh Water | Sodium Tripolyphosphate | Trehalose | Phosphorylated Trehalose |
|---|---|---|---|---|---|
| Thawing loss (%) | 0 | 5.69 ± 0.21 Ab | 4.79 ± 0.18 Aa | 4.92 ± 0.20 Aa | 4.86 ± 0.17 Aa |
| 3 | 6.23 ± 0.19 Bb | 4.93 ± 0.16 Aa | 5.01 ± 0.13 Aa | 4.90 ± 0.19 Aa | |
| 6 | 6.79 ± 0.18 Cc | 5.72 ± 0.14 Bb | 5.40 ± 0.16 Bab | 5.32 ± 0.11 Ba | |
| 9 | 7.34 ± 0.21 Dc | 6.12 ± 0.19 Cb | 5.66 ± 0.20 Ba | 5.44 ± 0.17 BCa | |
| 12 | 8.52 ± 0.20 Ed | 6.55 ± 0.17 Dc | 5.92 ± 0.15 Cb | 5.59 ± 0.16 Ca | |
| Cooking loss (%) | 0 | 8.65 ± 0.24 Ab | 5.34 ± 0.19 Aa | 5.43 ± 0.21 Aa | 5.19 ± 0.25 Aa |
| 3 | 9.53 ± 0.26 Bb | 5.53 ± 0.21 Aa | 5.61 ± 0.25 Aa | 5.33 ± 0.23 Aa | |
| 6 | 10.44 ± 0.36 Cc | 6.24 ± 0.23 Bab | 6.35 ± 0.26 Bb | 5.77 ± 0.19 Ba | |
| 9 | 11.89 ± 0.41 Dc | 6.89 ± 0.20 Cab | 7.11 ± 0.19 Cb | 6.50 ± 0.21 Ca | |
| 12 | 13.25 ± 0.37 Ec | 7.46 ± 0.19 Db | 7.54 ± 0.29 Cb | 6.97 ± 0.17 Da |
| MPs | Storage Time (Week) | Fresh Water | Sodium Tripolyphosphate | Trehalose | Phosphorylated Trehalose | ||||
|---|---|---|---|---|---|---|---|---|---|
| Tmax (°C) | ΔH (J/g) | Tmax (°C) | ΔH (J/g) | Tmax (°C) | ΔH (J/g) | Tmax (°C) | ΔH (J/g) | ||
| Myosin | 0 | 50.05 ± 0.21 Ea | 0.582 ± 0.012 Ea | 50.61 ± 0.24 Da | 0.600 ± 0.010 Ea | 50.49 ± 0.19 Da | 0.590 ± 0.011 Da | 50.66 ± 0.27 Da | 0.602 ± 0.009 Da |
| 3 | 49.38 ± 0.20 Da | 0.534 ± 0.010 Da | 50.17 ± 0.19 Db | 0.576 ± 0.008 Db | 50.11 ± 0.28 Db | 0.579 ± 0.009 Db | 50.23 ± 0.21 Db | 0.586 ± 0.010 Db | |
| 6 | 48.11 ± 0.18 Ca | 0.491 ± 0.012 Ca | 49.26 ± 0.22 Cb | 0.528 ± 0.009 Cb | 49.10 ± 0.25 Cb | 0.529 ± 0.010 Cb | 49.38 ± 0.14 Cb | 0.536 ± 0.007 Cb | |
| 9 | 46.56 ± 0.23 Ba | 0.450 ± 0.011 Ba | 48.04 ± 0.23 Bb | 0.490 ± 0.011 Bb | 48.16 ± 0.18 Bbc | 0.499 ± 0.008 Bb | 48.60 ± 0.18 Bc | 0.512 ± 0.005 Bc | |
| 12 | 44.21 ± 0.29 Aa | 0.413 ± 0.012 Aa | 46.11 ± 0.26 Ab | 0.455 ± 0.010 Ab | 46.08 ± 0.27 Ab | 0.458 ± 0.011 Ab | 47.22 ± 0.23 Ac | 0.478 ± 0.007 Ac | |
| Actin | 0 | 78.76 ± 0.31 Ea | 0.312 ± 0.004 Da | 79.01 ± 0.32 Dab | 0.315 ± 0.003 Da | 78.89 ± 0.26 Da | 0.318 ± 0.005 Da | 79.38 ± 0.30 Db | 0.322 ± 0.003 Da |
| 3 | 77.21 ± 0.22 Da | 0.308 ± 0.005 Da | 78.90 ± 0.24 Db | 0.311 ± 0.004 Da | 78.62 ± 0.31 Db | 0.310 ± 0.003 Da | 78.99 ± 0.22 Db | 0.313 ± 0.002 Ca | |
| 6 | 76.06 ± 0.27 Ca | 0.286 ± 0.004 Ca | 77.67 ± 0.36 Cb | 0.298 ± 0.002 Cb | 77.88 ± 0.29 Cb | 0.301 ± 0.003 Cb | 77.70 ± 0.34 Cb | 0.300 ± 0.004 Bb | |
| 9 | 74.36 ± 0.35 Ba | 0.264 ± 0.003 Ba | 75.83 ± 0.27 Bb | 0.280 ± 0.004 Bb | 76.65 ± 0.34 Bc | 0.290 ± 0.002 Bc | 76.77 ± 0.28 Bc | 0.292 ± 0.003 Bc | |
| 12 | 71.15 ± 0.28 Aa | 0.240 ± 0.005 Aa | 74.53 ± 0.24 Ab | 0.263 ± 0.003 Ab | 75.09 ± 0.21 Ac | 0.271 ± 0.004 Ab | 75.41 ± 0.20 Ad | 0.281 ± 0.002 Ac | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shui, S.; Yang, H.; Lu, B.; Zhang, B. Phosphorylated Trehalose Suppresses the Denaturation of Myofibrillar Proteins in Peeled Shrimp (Litopenaeus vannamei) during Long-Term Frozen Storage. Foods 2022, 11, 3189. https://doi.org/10.3390/foods11203189
Shui S, Yang H, Lu B, Zhang B. Phosphorylated Trehalose Suppresses the Denaturation of Myofibrillar Proteins in Peeled Shrimp (Litopenaeus vannamei) during Long-Term Frozen Storage. Foods. 2022; 11(20):3189. https://doi.org/10.3390/foods11203189
Chicago/Turabian StyleShui, Shanshan, Huicheng Yang, Baiyi Lu, and Bin Zhang. 2022. "Phosphorylated Trehalose Suppresses the Denaturation of Myofibrillar Proteins in Peeled Shrimp (Litopenaeus vannamei) during Long-Term Frozen Storage" Foods 11, no. 20: 3189. https://doi.org/10.3390/foods11203189
APA StyleShui, S., Yang, H., Lu, B., & Zhang, B. (2022). Phosphorylated Trehalose Suppresses the Denaturation of Myofibrillar Proteins in Peeled Shrimp (Litopenaeus vannamei) during Long-Term Frozen Storage. Foods, 11(20), 3189. https://doi.org/10.3390/foods11203189