
Correlation Analysis between GlpQ-Regulated Degradation of Wall Teichoic Acid and Biofilm Formation Triggered by Lactobionic Acid in Staphylococcus aureus



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents, Bacterial Strains, and Growth Conditions
2.2. Minimum Inhibitory Concentration (MIC)
2.3. Biofilm Formation Assay
2.4. Light Microscopy of S. aureus Adhesion
2.5. Scanning Electron Microscopy (SEM) of S. aureus Biofilms
2.6. Determination of WTA by Native-Polyacrylamide gel Electrophoresis (Native-PAGE)
2.6.1. Extraction of WTA
2.6.2. Native-PAGE Analysis of WTA
2.7. RNA Isolation and qRT-PCR
2.8. Statistical Analysis
3. Results and Discussion
3.1. In Vitro Antibacterial Susceptibility Testing
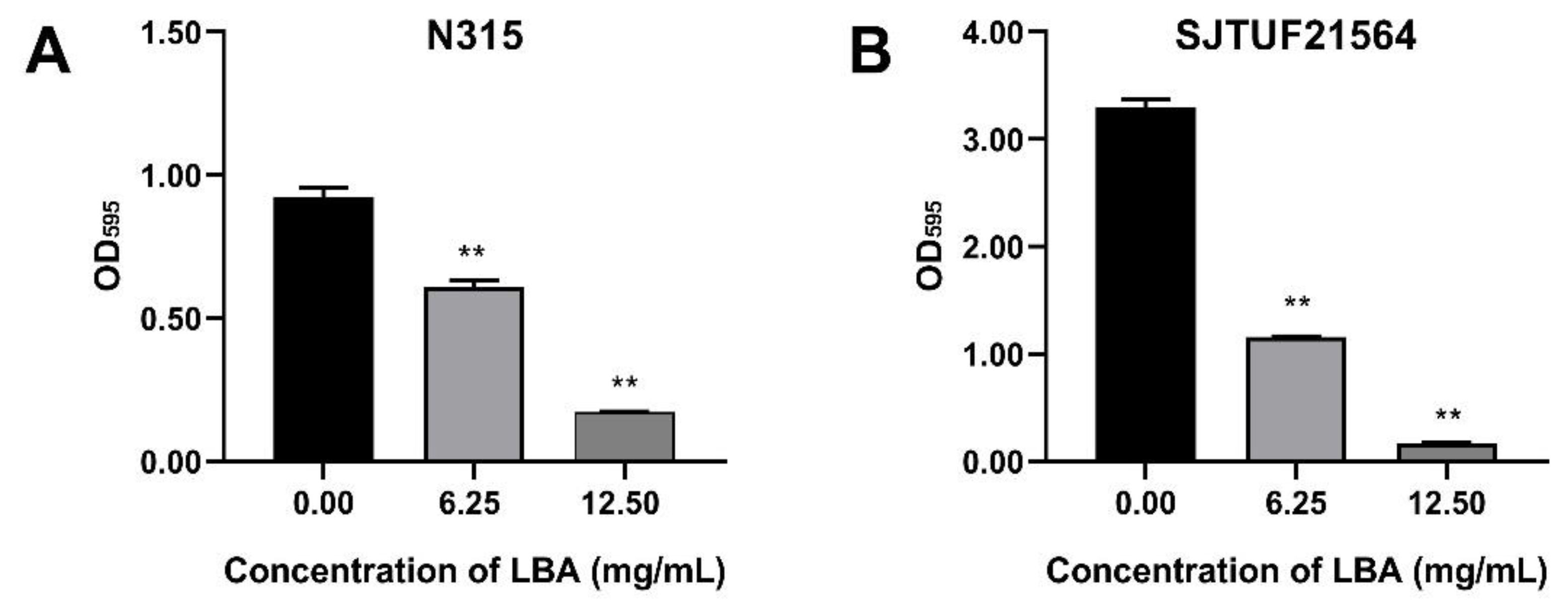
3.2. Effect of LBA on Biofilm Formation of S. aureus
3.3. Effect of LBA on Adherence of S. aureus
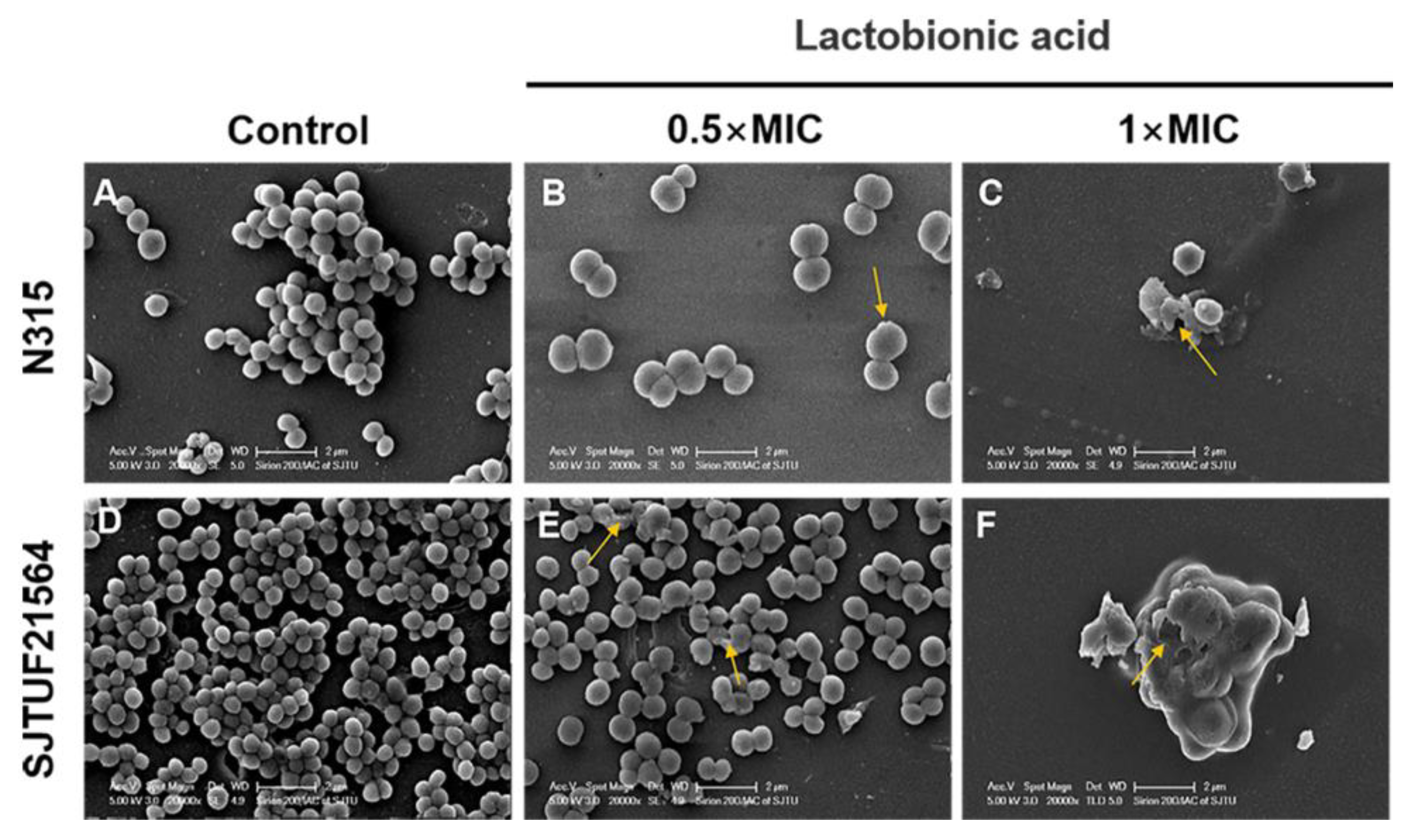
3.4. SEM Visualization of S. aureus Biofilm Formation
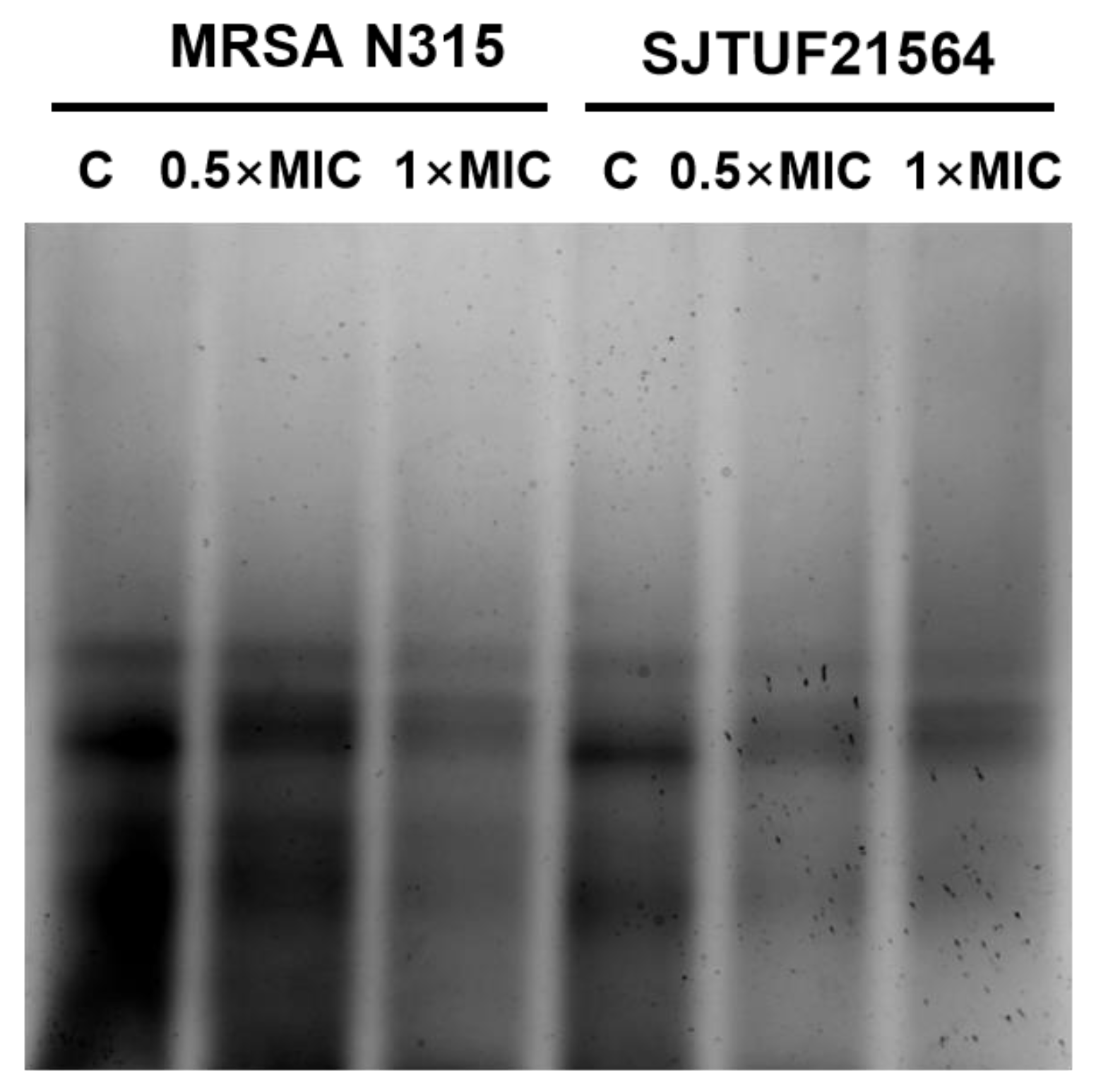
3.5. Effect of LBA on WTA Content
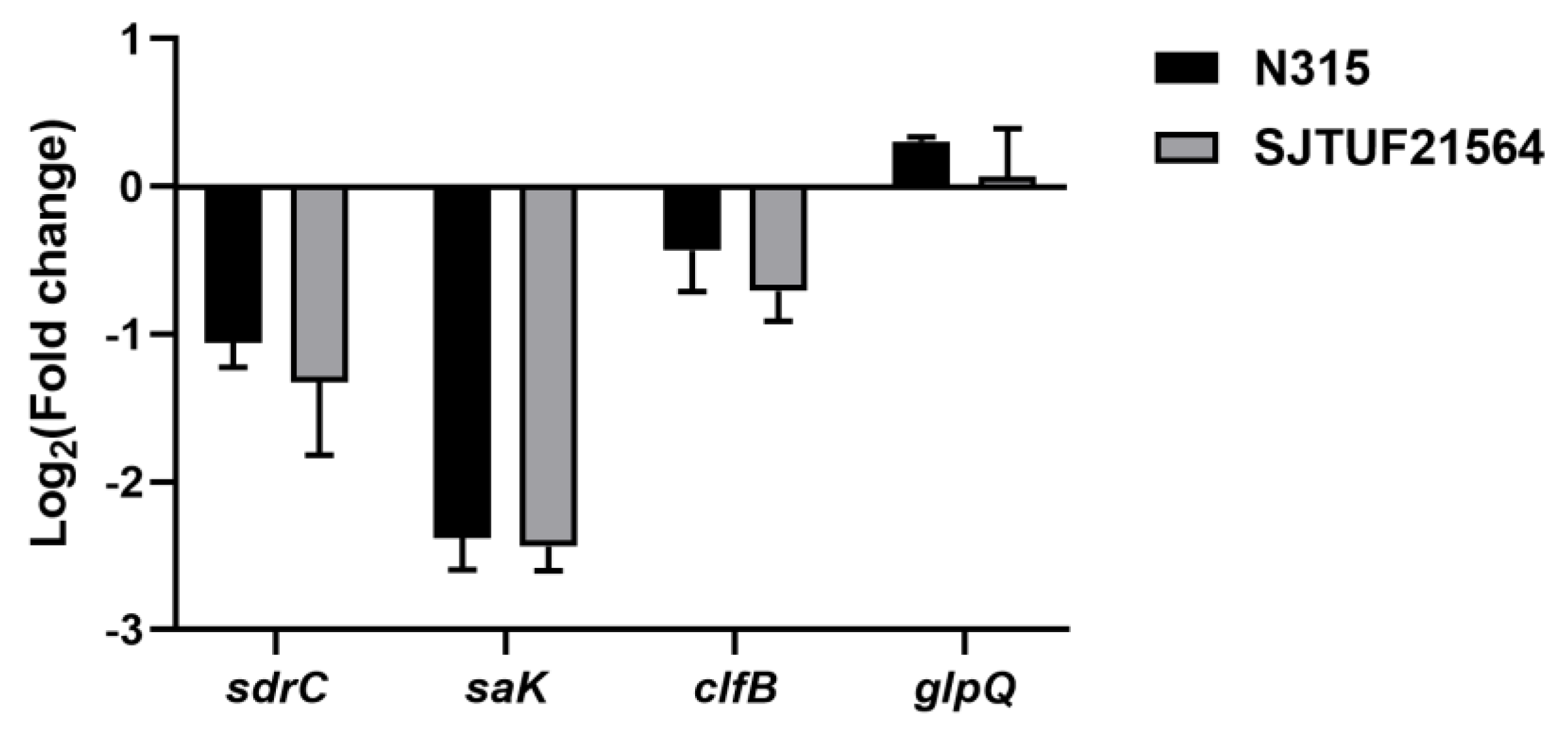
3.6. Analysis of WTA Hydrolase GlpQ Associated with S. aureus Biofilm Formation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, H.; Hou, X.; Shen, J.W.; Wang, W.H.; Ye, Y.W.; Yu, J.L.; Xue, T. Alternative sigma factor B reduces biofilm formation and stress response in milk-derived Staphylococcus aureus. LWT 2022, 162, 113515. [Google Scholar] [CrossRef]
- Ryu, S.; Shin, M.; Yun, B.; Lee, W.; Choi, H.; Kang, M.; Oh, S.; Kim, Y. Bacterial quality, prevalence of pathogens, and molecular characterization of biofilm-producing Staphylococcus aureus from Korean dairy farm environments. Animals 2021, 11, 1306. [Google Scholar] [CrossRef] [PubMed]
- Victoria, J.S.; Ian, C.; Alex, J.O. Staphylococcus aureus biofilms promote horizontal transfer of antibiotic resistance. Antimicrob. Agents Chemother. 2013, 57, 4. [Google Scholar]
- Campbell, J.; Singh, A.K.; Swoboda, J.G.; Gilmore, M.S.; Walker, S. An antibiotic that inhibits a late step in wall teichoic acid biosynthesis induces the cell wall stress stimulon in Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergara-Irigaray, M.; Maira-Litrán, T.; Merino, N.; Pier, G.B.; Penadés, J.; Iñigo, L. Wall teichoic acids are dispensable for anchoring the PNAG exopolysaccharide to the Staphylococcus aureus cell surface. Microbiology 2008, 154, 865–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kim, N.H.; Winstel, V.; Kurokawa, K.; Larsen, J.; An, J.H.; Khan, A.; Seong, M.H.; Lee, M.J.; Andersen, P.S.; et al. Surface glycopolymers are crucial for in vitro anti-wall teichoic acid IgG-mediated complement activation and opsonophagocytosis of Staphylococcus aureus. Infect. Immun. 2015, 83, 4247–4255. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Pfahler, N.M.; Vlpel, S.L.; Stehle, T. Cell wall glycosylation in Staphylococcus aureus: Targeting the tar glycosyltransferases. Curr. Opin. Struct. Biol. 2021, 68, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Dalen, R.V.; Peschel, A.; Sorge, N. Wall teichoic acid in Staphylococcus aureus host interaction. Trends Microbiol. 2020, 28, 985–998. [Google Scholar] [CrossRef]
- Gross, M.; Cramton, S.E.; Götz, F.L.; Peschel, A. Key role of teichoic acid net charge in Staphylococcus aureus colonization of artificial surfaces. Infect. Immun. 2001, 69, 3423–3426. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Han, J.; Gong, G.L.; Koffas, M.A.G.; Zha, J. Wall teichoic acids: Physiology and applications. FEMS Microbiol. Rev. 2020, 45, fuaa064. [Google Scholar] [CrossRef]
- Jorge, A.M.; Schneider, J.; Unsleber, S.; Xia, G.Q.; Mayer, C.; Peschel, A. Staphylococcus aureus counters phosphate limitation by scavenging wall teichoic acids from other staphylococci via the teichoicase GlpQ. J. Biol. Chem. 2018, 293, 14916–14924. [Google Scholar] [CrossRef]
- Walter, A.; Unsleber, S.; Rismondo, J.; Rismondo, J.; Jorge, A.M.; Peschel, A.; Gründling, A.; Mayer, C. Phosphoglycerol-type wall and lipoteichoic acids are enantiomeric polymers differentiated by the stereospecific glycerophosphodiesterase GlpQ. J. Biol. Chem. 2020, 295, 4024–4034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.M.; Kong, F.H.; Liang, X.N.; Li, M.H.; Yang, N.; Cao, X.Y.; Yang, M.; Tao, D.B.; Yue, X.Q.; Zheng, Y. Label-free quantitative proteomics reveals the multitargeted antibacterial mechanisms of lactobionic acid against methicillin-resistant Staphylococcus aureus (MRSA) using SWATH-MS technology. J. Agric. Food Chem. 2019, 67, 12322–12332. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagai, Y.; et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Krmusaolu, S. The Methods for Detection of Biofilm and Screening Antibiofilm Activity of Agents, 15th ed.; IntechOpen: Britain, UK, 2019; pp. 1–17. [Google Scholar]
- Wayne, P.A. Methods for Dilution Antimicrobial Susceptibility Test for Bacteria that Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; M07. [Google Scholar]
- Riera, M.; Moreno-Ruiz, E.; Goyard, S.; d’Enfert, C.; Janbon, G. Biofilm Formation Studies in Microtiter Plate Format. In Methods in Molecular Biology; Humana: Totowa, NJ, USA, 2012; Volume 845. [Google Scholar]
- Goswami, S.; Sarkar, R.; Saha, P.; Maity, A.; Sarkar, T.; Das, D.; Chakraborty, P.D.; Bandyopadhyay, S.; Ghosh, C.K.; Karmakar, S.; et al. Effect of human placental extract in the management of biofilm mediated drug resistance-A focus on wound management. Microb. Pathog. 2017, 111, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Luis, A.; Silva, F.; Sousa, S.; Duarte, A.P.; Domingues, F. Antistaphylococcal and biofilm inhibitory activities of gallic, caffeic, and chlorogenic acids. Biofouling 2014, 30, 69–79. [Google Scholar] [CrossRef]
- Meredith, T.C.; Swoboda, J.G.; Walker, S. Late-Stage Polyribitol Phosphate Wall Teichoic Acid Biosynthesis in Staphylococcus aureus. J. Bacteriol. 2008, 190, 3046–3056. [Google Scholar] [CrossRef] [Green Version]
- Pollack, J.H.; Neuhaus, F.C. Changes in wall teichoic acid during the rod-sphere transition of Bacillus subtilis 168. J. Bacteriol. 1994, 176, 7252–7259. [Google Scholar] [CrossRef] [Green Version]
- Wojnicz, D. The influence of subinhibitory concentrations of antibiotics on the bacterial adhesion. Adv. Clin. Exp. Med. 2007, 16, 141–148. [Google Scholar]
- Knidel, C.; Monalessa, F.P.; Barcelos, D.H.F.; Gomes, D.C.O.; Guimares, M.C.C.; Schuenck, R.P. Epigallocatechin gallate has antibacterial and antibiofilm activity in methicillin resistant and susceptible Staphylococcus aureus of different lineages in non-cytotoxic concentrations. Nat. Prod. Res. 2019, 35, 4643–4647. [Google Scholar] [CrossRef]
- Bai, J.R.; Zhong, K.; Wu, Y.P.; Elena, G.; Gao, H. Antibiofilm activity of shikimic acid against Staphylococcus aureus. Food Control 2019, 95, 327–333. [Google Scholar] [CrossRef]
- Raja, A.F.; Ali, F.; Khan, I.A.; Shawl, A.S.; Taneja, S.C. Antistaphylococcal and biofilm inhibitory activities of acetyl-11-keto-β-boswellic acid from Boswellia serrata. BMC Microbiol. 2011, 11, 54. [Google Scholar] [CrossRef]
- Akbas, M.Y.; Kokumer, T. The prevention and removal of biofilm formation of Staphylococcus aureus strains isolated from raw milk samples by citric acid treatments. Int. J. Food Sci. Technol. 2015, 50, 1666–1672. [Google Scholar] [CrossRef]
- Dula, S.; Ajayeoba, T.A.; Ijabadeniyi, O.A. Bacterial biofilm formation on stainless steel in the food processing environment and its health implications. Folia Microbiol. 2021, 66, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Agerer, F.; Lux, S.; Michel, A.; Rohde, M.; Ohlsen, K.; Hauck, C.R. Cellular invasion by Staphylococcus aureus reveals a functional link between focal adhesion kinase and cortactin in integrin-mediated internalization. J. Cell Sci. 2005, 15, 2189–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauck, C.R.; Agerer, F.; Muenzner, P.; Schmitter, T. Cellular adhesion molecules as targets for bacterial infection. Eur. J. Cell Biol. 2006, 85, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Feuillie, C.; Formosa-Dague, C.; Hays, L.M.; Vervaeck, O.; Derclaye, S.; Brennan, M.P.; Foster, T.J.; Geoghegan, J.A.; Dufrêne, Y.F. Molecular interactions and inhibition of the staphylococcal biofilm-forming protein SdrC. Proc. Natl. Acad. Sci. USA 2017, 114, 3738–3743. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, E.; Pozzi, C.; Houston, P.; Humphreys, H.; Robinson, D.A.; Loughman, A.; Foster, T.J.; O’Gara, J.P. A novel Staphylococcus aureus biofilm phenotype mediated by the fibronectin-binding proteins, FnBPA and FnBPB. J. Bacteriol. 2008, 190, 3835–3850. [Google Scholar] [CrossRef] [Green Version]
- Corrigan, R.M.; Miajlovic, H.; Foster, T.J. Surface proteins that promote adherence of Staphylococcus aureus to human desquamated nasal epithelial cells. BMC Microbiol. 2009, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, N.; Jian, Y.; Liu, Y.; Zhao, L.; He, L.; Liu, Q.; Li, M. The pro-inflammatory effect of Staphylokinase contributes to community-associated Staphylococcus aureus pneumonia. Commun. Biol. 2022, 5, 618. [Google Scholar] [CrossRef] [PubMed]
- Rooijakkers, S.H.; Ruyken, M.; van Roon, J.; van Kessel, K.P.; van Strijp, J.A.; van Wamel, W.J. Early expression of SCIN and CHIPS drives instant immune evasion by Staphylococcus aureus. Cell Microbiol. 2006, 8, 1282–1293. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.R.; Swierstra, J.W.; Lemmens-den, T.N.A.; Snijders, S.V.; Hansenova, M.S.; Verbon, A.; Wamel, W.J.B. Production of Staphylococcal Complement Inhibitor (SCIN) and other immune modulators during the early stages of Staphylococcus aureus biofilm formation in a mammalian cell culture medium. Infect. Immun. 2018, 86, e00352-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resch, A.; Rosenstein, R.; Nerz, C.; Gotz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, N.M.; Jefferson, K.K. Staphylococcus aureus clumping factor B mediates biofilm formation in the absence of calcium. Microbiology 2012, 158, 1504–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, X.; Ma, Q.; Wang, G.; Li, N.; Mao, Y.; Wang, X.; Wang, Y.; Wang, G. Potential mechanisms of quercetin influence the ClfB protein during biofilm formation of Staphylococcus aureus. Front. Pharmacol. 2022, 13, 825489. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Kraus, F.; Popella, P.; Baykal, A.; Guttroff, C.; François, P.; Sass, P.; Plietker, B.; Götz, F. The Polycyclic Polyprenylated Acylphloroglucinol Antibiotic PPAP 23 Targets the Membrane and Iron Metabolism in Staphylococcus aureus. Front. Microbiol. 2019, 10, 14. [Google Scholar] [CrossRef]
- Fan, Q.X.; He, Q.; Zhang, T.; Song, W.; Sheng, Q.L.; Yuan, Y.H.; Yue, T.L. Antibiofilm potential of lactobionic acid against Salmonella Typhimurium. LWT 2022, 162, 113461. [Google Scholar] [CrossRef]
- Fan, Q.X.; Yuan, Y.H.; Zhang, T.; Song, W.; Sheng, Q.L.; Yue, T.L. Inhibitory effects of lactobionic acid on Vibrio parahaemolyticus planktonic cells and biofilms. Food Microbiol. 2022, 103, 103963. [Google Scholar] [CrossRef]
- Kang, S.M.; Kong, F.H.; Shi, X.Y.; Han, H.J.; Li, M.H.; Guan, B.Y.; Yang, M.; Cao, X.Y.; Tao, D.B.; Zheng, Y. Antibacterial activity and mechanism of lactobionic acid against Pseudomonas fluorescens and Methicillin-resistant Staphylococcus aureus and its application on whole milk. Food Control 2020, 108, 106876. [Google Scholar] [CrossRef]
- Cao, J.R.; Fu, H.J.; Gao, L.H.; Zheng, Y. Antibacterial activity and mechanism of lactobionic acid against Staphylococcus aureus. Folia Microbiol. 2019, 64, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gill, C.J.; Lee, S.H.; Mann, P.; Zuck, P.; Meredith, T.C.; Murgolo, N.; She, X.W.; Kales, S.; Liang, L.Z.; et al. Discovery of Wall Teichoic Acid Inhibitors as Potential Anti-MRSA β-Lactam Combination Agents. Chem. Biol. 2013, 20, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, A.; van Dalen, R.; Ali, S.; Gerlach, D.; van der Marel, G.A.; Fuchsberger, F.F.; Aerts, P.C.; de Haas, C.J.C.; Peschel, A.; Rademacher, C.; et al. Impact of glycan linkage to Staphylococcus aureus wall teichoic acid on langerin recognition and langerhans cell activation. ACS Infect. Dis. 2021, 7, 624–635. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) |
|---|---|
| glpQ | F: ATTTATGGCTGCTTCTGCTGT R: GCTTGAAACGTATGCTCTGGT |
| sdrC | F: TGATAAAGATGCCGATGGTGG R: CGCTGTCTGAATCGCTGTCTG |
| saK | F: TGTAGTCCCAGGTTTAATAGG R: CGCGAGTTATTTTGAACC |
| clfB | F: ATAGGCAATCATCAAGCA R: TGTATCATTAGCCGTTGTAT |
| rpoB (housekeeping gene) | F: ATGACTTAGCAAGCGTGGGT R: GCGTTCGATTCAAGTACATCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, W.; Kang, S.; Chang, J.; Tian, X.; Shi, C. Correlation Analysis between GlpQ-Regulated Degradation of Wall Teichoic Acid and Biofilm Formation Triggered by Lactobionic Acid in Staphylococcus aureus. Foods 2022, 11, 3438. https://doi.org/10.3390/foods11213438
Hou W, Kang S, Chang J, Tian X, Shi C. Correlation Analysis between GlpQ-Regulated Degradation of Wall Teichoic Acid and Biofilm Formation Triggered by Lactobionic Acid in Staphylococcus aureus. Foods. 2022; 11(21):3438. https://doi.org/10.3390/foods11213438
Chicago/Turabian StyleHou, Wanwan, Shimo Kang, Jiang Chang, Xiaorong Tian, and Chunlei Shi. 2022. "Correlation Analysis between GlpQ-Regulated Degradation of Wall Teichoic Acid and Biofilm Formation Triggered by Lactobionic Acid in Staphylococcus aureus" Foods 11, no. 21: 3438. https://doi.org/10.3390/foods11213438
APA StyleHou, W., Kang, S., Chang, J., Tian, X., & Shi, C. (2022). Correlation Analysis between GlpQ-Regulated Degradation of Wall Teichoic Acid and Biofilm Formation Triggered by Lactobionic Acid in Staphylococcus aureus. Foods, 11(21), 3438. https://doi.org/10.3390/foods11213438