
Isolation, Characterization and Antioxidant Activity of Yam Polysaccharides



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction and Purification of Polysaccharides
2.3. Structure Characterization
2.3.1. Determination of the Content of Total Carbohydrate, Uronic Acid and Protein
2.3.2. Determination of Molecular Weight (Mw)
2.3.3. Monosaccharide Composition
2.3.4. Scanning Electron Microscope (SEM) Analysis
2.3.5. Spectra Analysis
2.4. Cytoprotective Activity
2.4.1. Cell Viability Assay
2.4.2. Measurement of SOD and MDA Levels
2.4.3. Measurement of Intracellular ROS Level
2.4.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Physicochemical Properties and Molecular Weight
3.2. Monosaccharide Compositions
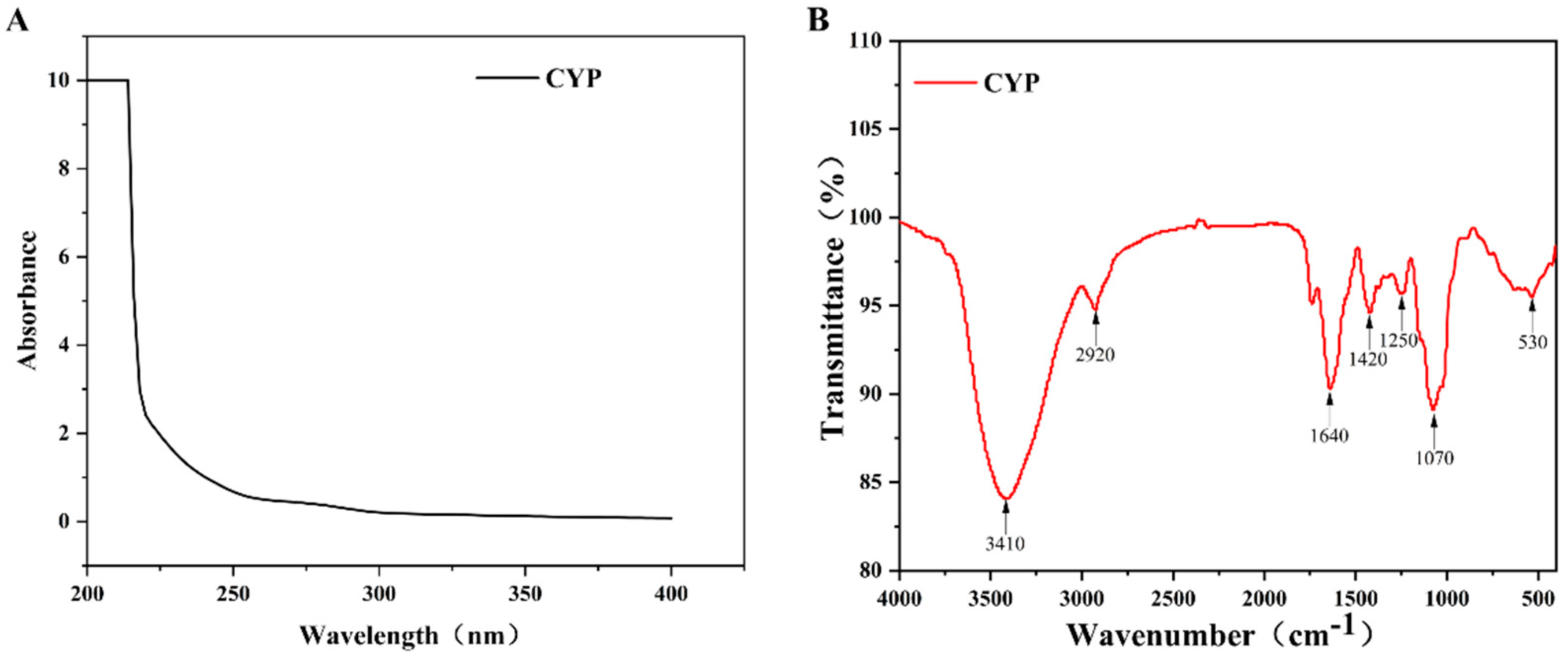
3.3. UV and FT-IR Spectrum Analysis
3.4. SEM Analysis
3.5. Effect of CYP on IEC-6 Cells Viability
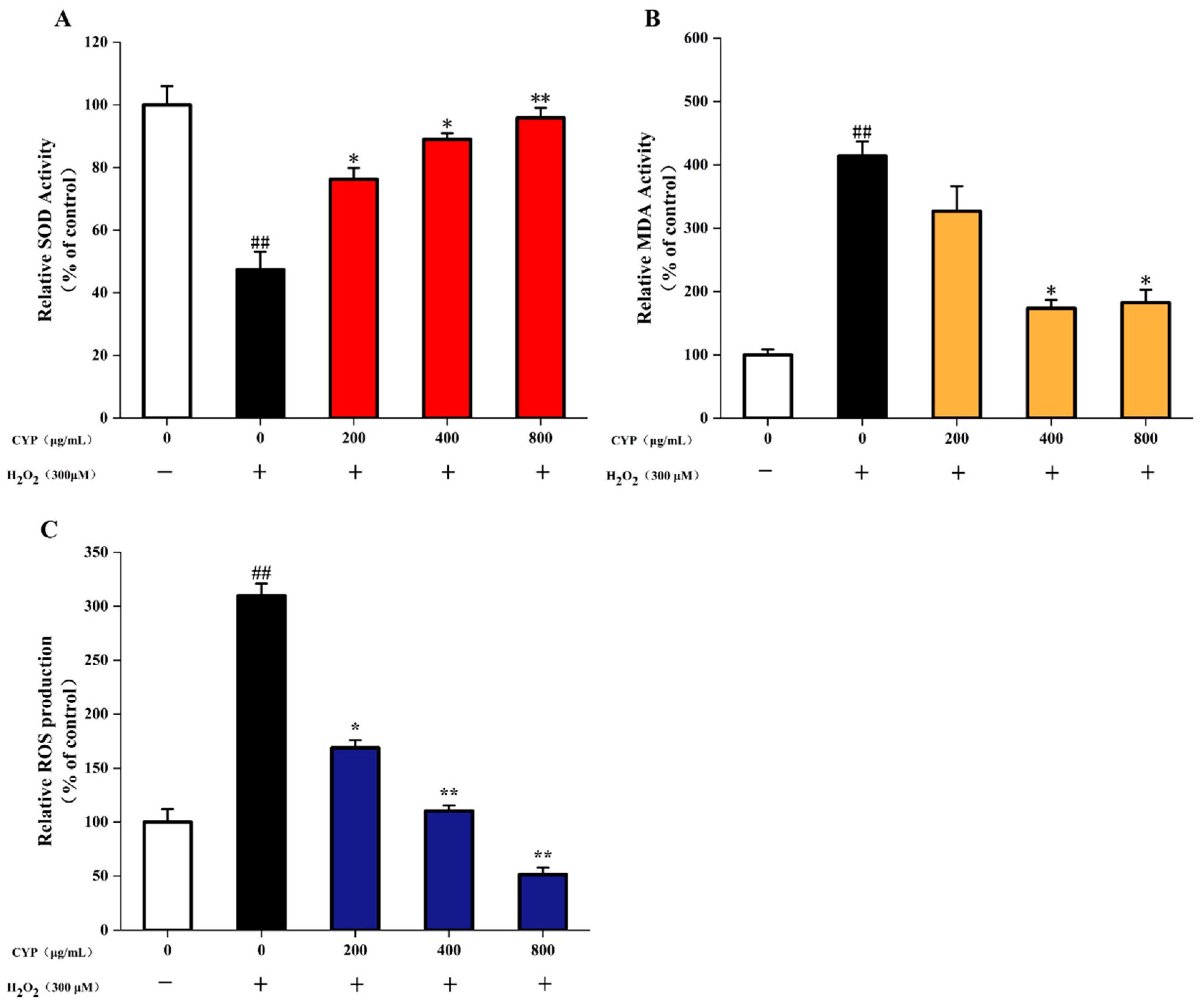
3.6. Effects of CYP on MDA and SOD in H2O2-Stimulated IEC-6 Cells
3.7. ROS Production in H2O2-Induced IEC-6 Cells
3.8. Effect of CYP on MAPK Pathway in Oxidative Stressed IEC-6 Cells
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Medini, F.; Bourgou, S.; Lalancette, K.G.; Snoussi, M.; Mkadmini, K.; Coté, I.; Abdelly, C.; Legault, J.; Ksouri, R. Phytochemical analysis, antioxidant, anti-inflammatory, and anticancer activities of the halophyte Limonium densiflorum extracts on human cell lines and murine macrophages. S. Afr. J. Bot. 2015, 99, 158–164. [Google Scholar] [CrossRef]
- Yun, B.; King, M.; Draz, M.S.; Kline, T.; Alex, R.P. Oxidative reactivity across kingdoms in the gut: Host immunity, stressed microbiota and oxidized foods. Free Radic. Biol. Med. 2021, 178, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Mark, T.D.C.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Salinas, F.L.S.; Gonzalez, A.P.; Casique, A.A.; Ballote, A.I.; Diaz, A.; Treviño, S.; Murrieta, N.H.R.; Peña, L.M.P.; Maycotte, P. Reactive oxygen species: Role in carcinogenesis, cancer cell signaling and tumor progression. Life Sci. 2021, 284, 119942. [Google Scholar] [CrossRef] [PubMed]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Xie, L.M.; Shen, M.Y.; Wen, P.W.; Hong, Y.Z.; Liu, X.; Xie, J.H. Preparation, characterization, antioxidant activity and protective effect against cellular oxidative stress of phosphorylated polysaccharide from Cyclocarya paliurus. Food Chem. Toxicol. 2020, 145, 111754. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.W.; Zhou, C.H.; Liu, T.T.; Dai, Y.Y.; Huang, H.H. A novel Hericium erinaceus polysaccharide: Structural characterization and prevention of H2O2-induced oxidative damage in GES-1 cells. Int. J. Biol. Macromol. 2019, 154, 1460–1470. [Google Scholar] [CrossRef]
- Ghosh, J.; Das, J.; Manna, P.; Sil, P.C. Taurine prevents arsenic-induced cardiac oxidative stress and apoptotic damage: Role of NF-κB, p38 and JNK MAPK pathway. Toxicol. Appl. Pharmacol. 2009, 240, 73–87. [Google Scholar] [CrossRef]
- Qiu, H.M.; Veeraperumal, S.; Lv, J.H.; Wu, T.C.; Zhang, Z.P.; Zeng, Q.K.; Liu, Y.; Chen, X.Q.; Aweya, J.J.; Cheong, K.L. Physicochemical properties and potential beneficial effects of porphyran from Porphyra haitanensis on intestinal epithelial cells. Carbohydr. Polym. 2020, 246, 116626. [Google Scholar] [CrossRef]
- Yu, Y.; Zhu, H.; Shen, M.; Yu, Q.; Chen, Y.; Xie, J. Sulfation modification enhances the intestinal regulation of Cyclocarya paliurus polysaccharides in cyclophosphamide-treated mice via restoring intestinal mucosal barrier function and modulating gut microbiota. Food Funct. 2021, 12, 12278–12290. [Google Scholar] [CrossRef]
- Li, G.Q.; Chen, P.F.; Zhao, Y.T.; Zeng, Q.H.; Ou, S.Y.; Zhang, Y.H.; Wang, P.C.; Chen, N.H.; Ou, J.Y. Isolation, structural characterization and anti-oxidant activity of a novel polysaccharide from garlic bolt. Carbohydr. Polym. 2021, 267, 118194. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.L.; Tong, M.Y.; Zeng, H.L.; Zheng, B.D.; Hu, X.H. Structural characterization and in vitro fermentation by rat intestinal microbiota of a polysaccharide from Porphyra haitanensis. Food Res. Int. 2021, 147, 110546. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhu, C.P.; Huang, G.Q.; Xiao, J.X. Fractionation and structural characterization of polysaccharides derived from red grape pomace. Process Biochem. 2021, 109, 37–45. [Google Scholar] [CrossRef]
- Yang, W.Y.; Wang, Y.; Li, X.P.; Yu, P. Purification and structural characterization of Chinese yam polysaccharide and its activities. Carbohydr. Polym. 2015, 117, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xie, J.H.; Yu, Y.; Shen, M. Recent progress in the research of yam mucilage polysaccharides: Isolation, structure and bioactivities. Int. J. Biol. Macromol. 2019, 155, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huo, X.Z.; Liu, Z.; Yang, R.; Zeng, H.J. Investigations on the anti-aging activity of polysaccharides from Chinese yam and their regulation on klotho gene expression in mice. J. Mol. Struct. 2020, 1208, 127895. [Google Scholar] [CrossRef]
- Zhou, S.Y.; Huang, G.L.; Chen, G.Y. Extraction, structural analysis, derivatization and antioxidant activity of polysaccharide from Chinese yam. Food Chem. 2021, 361, 130089. [Google Scholar] [CrossRef]
- Luo, L.; Zheng, S.S.; Huang, Y.F.; Qin, T.; Xing, J.; Niu, Y.L.; Bo, R.N.; Liu, Z.G.; Huang, Y.; Hu, Y.L.; et al. Preparation and characterization of Chinese yam polysaccharide PLGA nanoparticles and their immunological activity. Int. J. Pharm. 2016, 511, 140–150. [Google Scholar] [CrossRef]
- Xue, H.Y.; Li, J.R.; Liu, Y.G.; Gao, Q.; Wang, X.W.; Zhang, J.W.; Tanokura, M.; Xue, Y.L. Optimization of the ultrafiltration-assisted extraction of Chinese yam polysaccharide using response surface methodology and its biological activity. Int. J. Biol. Macromol. 2019, 121, 1186–1193. [Google Scholar] [CrossRef]
- Liu, J.; Stefan, W.; Xu, C.L. A review of bioactive plant polysaccharides: Biological activities, functionalization, and biomedical applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 31–61. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Yang, L.; Zhang, C.N.; Tian, Y.L.; Zhang, F.F.; Li, X.T. Structural and functional analyses of three purified polysaccharides isolated from Chinese Huaishan-yams. Int. J. Biol. Macromol. 2018, 120, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.Y.; Dai, K.Y.; Liu, C.; Yu, J.; Jia, X.Y.; Liu, A.J. Preparation, Antioxidant and Immunoregulatory Activities of a Macromolecular Glycoprotein from Salvia miltiorrhiza. Foods 2020, 11, 705. [Google Scholar] [CrossRef]
- Li, P.; Xiao, N.; Zeng, L.P.; Xiao, J.; Huang, J.Z.; Xu, Y.N.; Chen, Y.L.; Ren, Y.H.; Du, B. Structural characteristics of a mannoglucan isolated from Chinese yam and its treatment effects against gut microbiota dysbiosis and DSS-induced colitis in mice. Carbohydr. Polym. 2020, 250, 116958. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, L.X.; Chen, S.R.; Deng, Y.; Zhao, J.; Wang, Y.; Li, S.P. Non-starch polysaccharide from Chinese yam activated RAW 264.7 macrophages through the Toll-like receptor 4 (TLR4)-NF-κB signaling pathway. J. Funct. Food. 2017, 37, 491–500. [Google Scholar] [CrossRef]
- Liu, X.X.; Yan, Y.Y.; Liu, H.M.; Wang, X.D.; Qin, G.Y. Emulsifying and structural properties of polysaccharides extracted from Chinese yam by an enzyme-assisted method. LWT-Food Sci. Technol. 2019, 111, 242–251. [Google Scholar] [CrossRef]
- Ju, Y.; Xue, Y.; Huang, J.L.; Zhai, Q.Z.; Wang, X.H. Antioxidant Chinese yam polysaccharides and its pro-proliferative effect on endometrial epithelial cells. Int. J. Biol. Macromol. 2014, 66, 81–85. [Google Scholar] [CrossRef]
- Hao, B.H.; Yang, X.; Ma, Y. Study on deproteinization in extraction of polysaccharides from Patentillaunserina by Sevage. Sci. Technol. Food Ind. 2011, 32, 254–258. [Google Scholar]
- Huang, L.X.; Shen, M.Y.; Zhang, X.W.; Jiang, L.; Song, Q.Q.; Xie, J.H. Effect of high-pressure treatment on the physicochemical properties and antioxidant activities of polysaccharide from Mesona chinensis Benth. Carbohydr. Polym. 2018, 200, 191–199. [Google Scholar] [CrossRef]
- Liu, H.; Chen, F.S.; Yang, H.S.; Yao, Y.Z.; Gong, X.Z.; Xin, Y.; Ding, C.H. Effect of calcium treatment on nanostructure of chelate-soluble pectin and physicochemical and textural properties of apricot fruits. Food Res. Int. 2009, 42, 1131–1140. [Google Scholar] [CrossRef]
- Guo, S.Y.; Li, L.; Wang, M.L.; Liang, G.J. The determination of protein content in polysaccharides from Stanuntonia Chinese with coomassie brilliant blue method. Food Res. Dev. 2008, 29, 115–116. [Google Scholar]
- Xu, J.; Chen, Z.Y.; Liu, P.H.; Wei, Y.; Zhang, M.; Huang, X.D.; Peng, L.L.; Wei, X.L. Structural characterization of a pure polysaccharide from Bletilla striata tubers and its protective effect against H2O2-induced injury fibroblast cells. Int. J. Biol. Macromol. 2021, 193, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Xie, J.H.; Kan, L.J.; Wang, J.Q.; Shen, M.Y.; Li, W.J.; Nie, S.P.; Xie, M.Y. Sulfated polysaccharides from Cyclocarya paliurus reduce H2O2-induced oxidative stress in RAW264.7 cells. Int. J. Biol. Macromol. 2015, 80, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.J.; Zhang, J.G.; Sun, Y.H.; Qu, J.; Li, L.; Prasad, C.; Wei, Z.J. Physicochemical properties and antioxidant activities of polysaccharides sequentially extracted from peony seed dreg. Int. J. Biol. Macromol. 2016, 91, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xie, J.H.; Liu, X.; Shen, M.Y. Sulfated modification enhances the modulatory effect of yam polysaccharide on gut microbiota in cyclophosphamide-treated mice. Food Res. Int. 2021, 145, 110393. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.L.; Zhang, F.; Zhang, R.; Liu, F.; Peng, Q.; Wang, M. An acidic polysaccharide from Ziziphus Jujuba cv. Muzao: Purification and structural characterization. Food Chem. 2019, 274, 494–499. [Google Scholar] [CrossRef]
- Jing, L.; Sheng, J.W.; Jiang, J.R.; Wang, Y.; Shen, X.Y.; Liu, D.M.; Zhang, W.F.; Mao, S.M. Chemical characteristics and cytoprotective activities of polysaccharide fractions from Athyrium Multidentatum (Doll.) Ching. Int. J. Biol. Macromol. 2020, 158, 1227–1238. [Google Scholar] [CrossRef]
- Wang, L.C.; Chen, L.Y.; Li, J.S.; Di, L.Q.; Wu, H. Structural elucidation and immune-enhancing activity of peculiar polysaccharides fractioned from marine clam Meretrix meretrix (Linnaeus). Carbohydr. Polym. 2018, 201, 500–513. [Google Scholar] [CrossRef]
- Chai, Z.; Huang, W.Y.; Zhao, X.; Wu, H.; Zeng, X.X.; Li, C.Y. Preparation, characterization, antioxidant activity and protective effect against cellular oxidative stress of polysaccharide from Cynanchum auriculatum Royle ex Wight. Int. J. Biol. Macromol. 2018, 119, 1068–1076. [Google Scholar] [CrossRef]
- Liao, W.Z.; Luo, Z.; Liu, D.; Ning, Z.X.; Yang, J.G.; Ren, J.Y. Structure Characterization of a Novel Polysaccharide from Dictyophora indusiata and Its Macrophage Immunomodulatory Activities. J. Agric. Food Chem. 2015, 63, 535–544. [Google Scholar] [CrossRef]
- Tang, W.; Liu, C.C.; Liu, J.J.; Hu, L.Y.; Huang, Y.S.; Yuan, L.; Liu, F.W.; Pan, S.J.; Chen, S.P.; Bian, S.G.; et al. Purification of polysaccharide from Lentinus edodes water extract by membrane separation and its chemical composition and structure characterization. Food Hydrocoll. 2020, 105, 105851. [Google Scholar] [CrossRef]
- Zong, P.Z.; Jiao, C.; Yi, C.; Yuntong, M.; Qing, S.Y.; Yun, Q.F.; Chao, M.F.; Boonjai, L.; Rui, L.; Wan, L. Extraction, structural characterization and antioxidant activity of turmeric polysaccharides. LWT-Food Sci. Technol. 2022, 154, 112805. [Google Scholar]
- Wang, L.F.; Shi, Z.X.; Wang, X.Y.; Mu, S.; Xu, X.Y.; Shen, L.; Li, P. Protective effects of bovine milk exosomes against oxidative stress in IEC-6 cells. Eur. J. Nutr. 2021, 60, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, M.B.; Chen, Z.Y.; Wang, G.; Su, Q.; Liu, Y.J. Preparation, structural characterization and neuroprotective effects of polysaccharides from the pericarp of Zanthoxylum bungeanum Maxim against H2O2-induced oxidative damage in PC12 cells. S. Afr. J. Bot. 2021, 142, 165–174. [Google Scholar] [CrossRef]
- Yu, Y.; Mo, S.R.; Shen, M.Y.; Chen, Y.; Yu, Q.; Li, Z.D.; Xie, J.H. Sulfated modification enhances the immunomodulatory effect of Cyclocarya paliurus polysaccharide on cyclophosphamide-induced immunosuppressed mice through MyD88-dependent MAPK/NF-κB and PI3K-Akt signaling pathways. Food Res. Int. 2021, 150, 110756. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Feng, R.; Yang, Z.; Liu, T.T.; Huo, T.G.; Jiang, H. Oxidative stress induced by realgar in neurons: p38 MAPK and ERK1/2 perturb autophagy and induce the p62-Keap1-Nrf2 feedback loop to activate the Nrf2 signalling pathway. J. Ethnopharmacol. 2022, 282, 114582. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CYP |
|---|---|
| Yield (%) | 0.21 ± 0.01 |
| Carbohydrate (%) | 33.62 ± 0.08 |
| Uronic acid (%) | 34.95 ±0.21 |
| Protein (%) | 5.21 ± 0.26 |
| Mw (kDa) | 20.89 |
| Monosaccharide compositions | Ratio (%) |
| Arabinose | 4.51 |
| Galactose | 28.57 |
| Glucose | 11.28 |
| Mannose | 6.77 |
| Xylose | 2.26 |
| Rhamnose | 4.89 |
| Galacturonic acid | 37.59 |
| Glucuronic acid | 4.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Xiao, W.; Xie, J.; Chen, Y.; Yu, Q.; Zhang, W.; Shen, M. Isolation, Characterization and Antioxidant Activity of Yam Polysaccharides. Foods 2022, 11, 800. https://doi.org/10.3390/foods11060800
Li Z, Xiao W, Xie J, Chen Y, Yu Q, Zhang W, Shen M. Isolation, Characterization and Antioxidant Activity of Yam Polysaccharides. Foods. 2022; 11(6):800. https://doi.org/10.3390/foods11060800
Chicago/Turabian StyleLi, Zhedong, Wenhao Xiao, Jianhua Xie, Yi Chen, Qiang Yu, Weidong Zhang, and Mingyue Shen. 2022. "Isolation, Characterization and Antioxidant Activity of Yam Polysaccharides" Foods 11, no. 6: 800. https://doi.org/10.3390/foods11060800
APA StyleLi, Z., Xiao, W., Xie, J., Chen, Y., Yu, Q., Zhang, W., & Shen, M. (2022). Isolation, Characterization and Antioxidant Activity of Yam Polysaccharides. Foods, 11(6), 800. https://doi.org/10.3390/foods11060800