
Maturation of Moristel in Different Vineyards: Amino Acid and Aroma Composition of Mistelles and Wines with Particular Emphasis in Strecker Aldehydes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents, Solvents and Standards
2.2. Samples
2.2.1. Grape Samples and Ethanolic Musts (Mistelles)
2.2.2. Wine Samples
2.3. Methods
2.3.1. Oenological Parameters
2.3.2. Quantitative Analysis of Major Aroma Compounds
2.3.3. Quantitative Analysis of Minor and Trace Aroma Compounds
2.3.4. Determination of Total Strecker Aldehydes
2.3.5. Quantitative Analysis of Amino Acids
2.4. Data Analysis
3. Results and Discussion
3.1. Chemical Changes in Grapes during Maturation
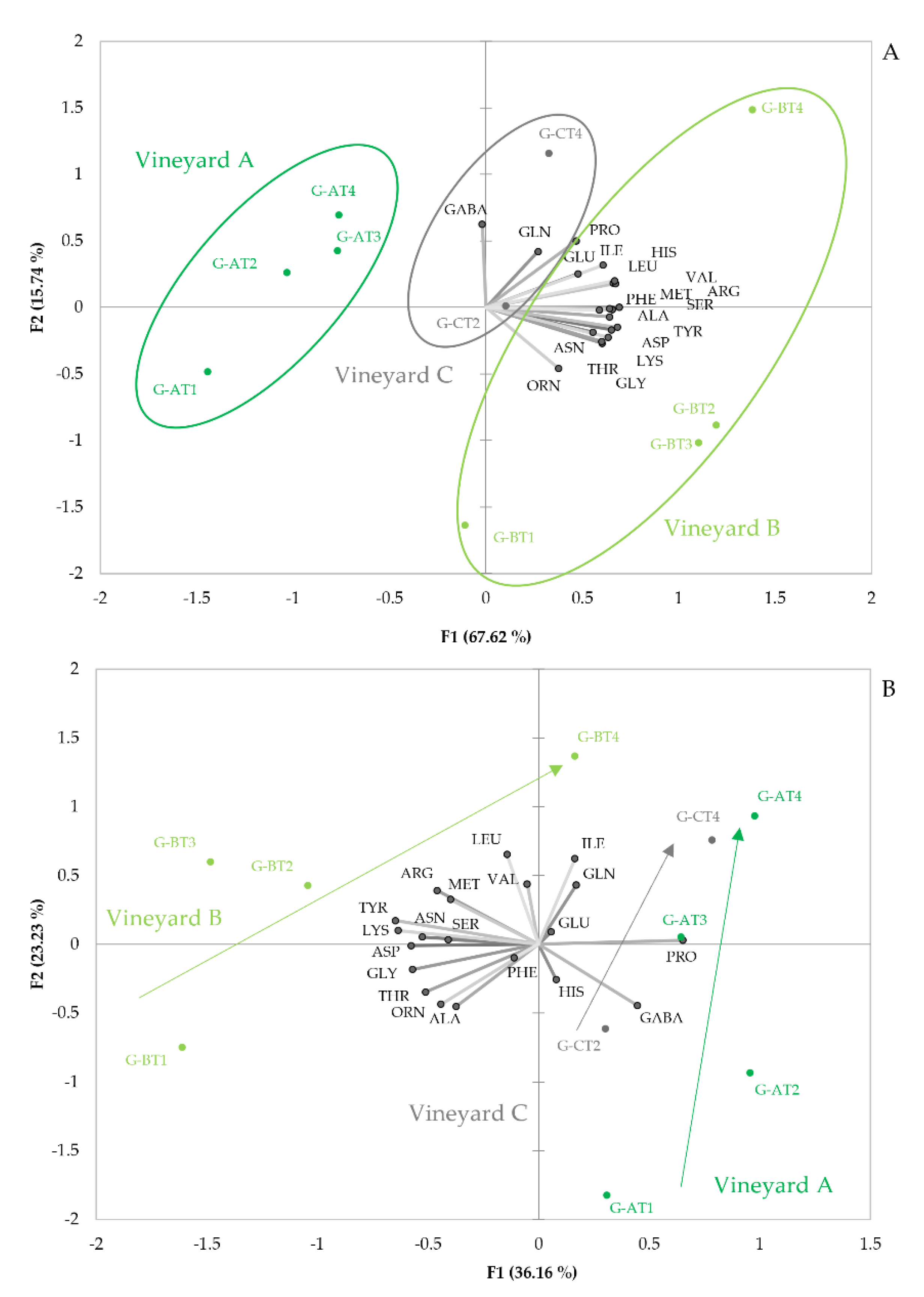
3.1.1. Amino Acid Profiles
3.1.2. Volatile Fraction
3.2. Wines and Their Relationship to Original Mistelles
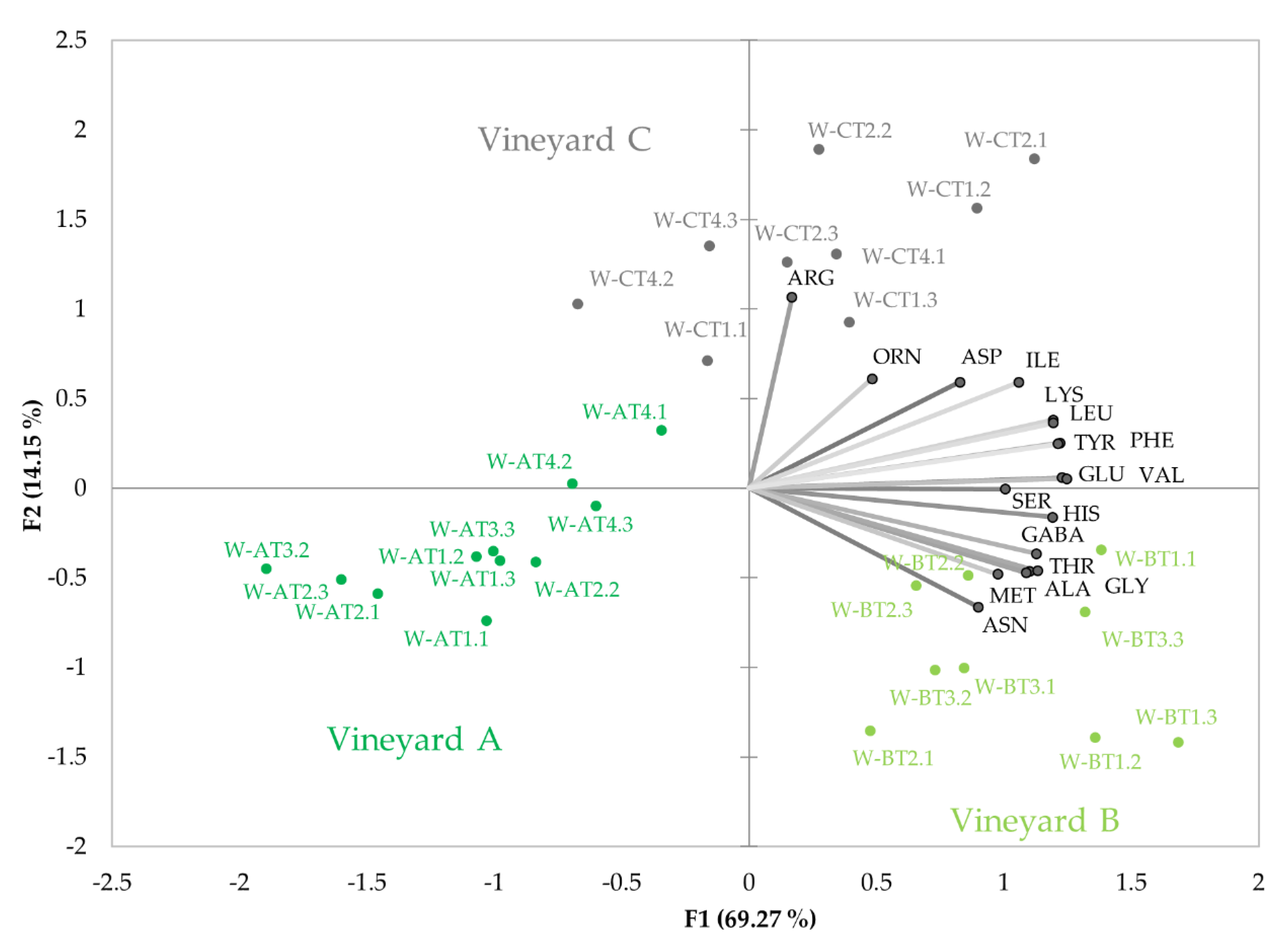
3.2.1. Amino Acid Changes during Fermentation
3.2.2. Amino Acid Consumption and Wine Volatiles
3.2.3. Amino Acid Profiles in Wines
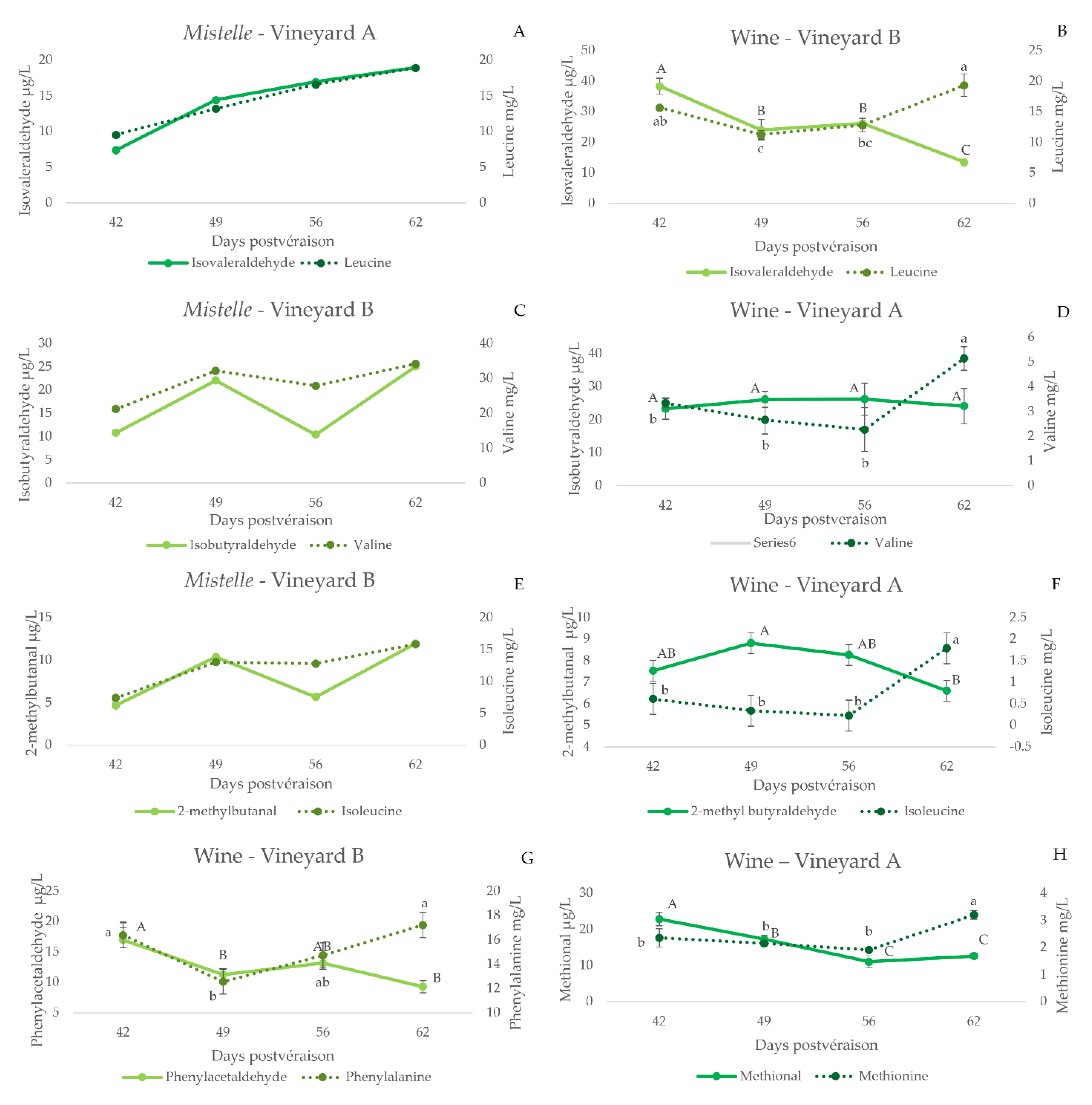
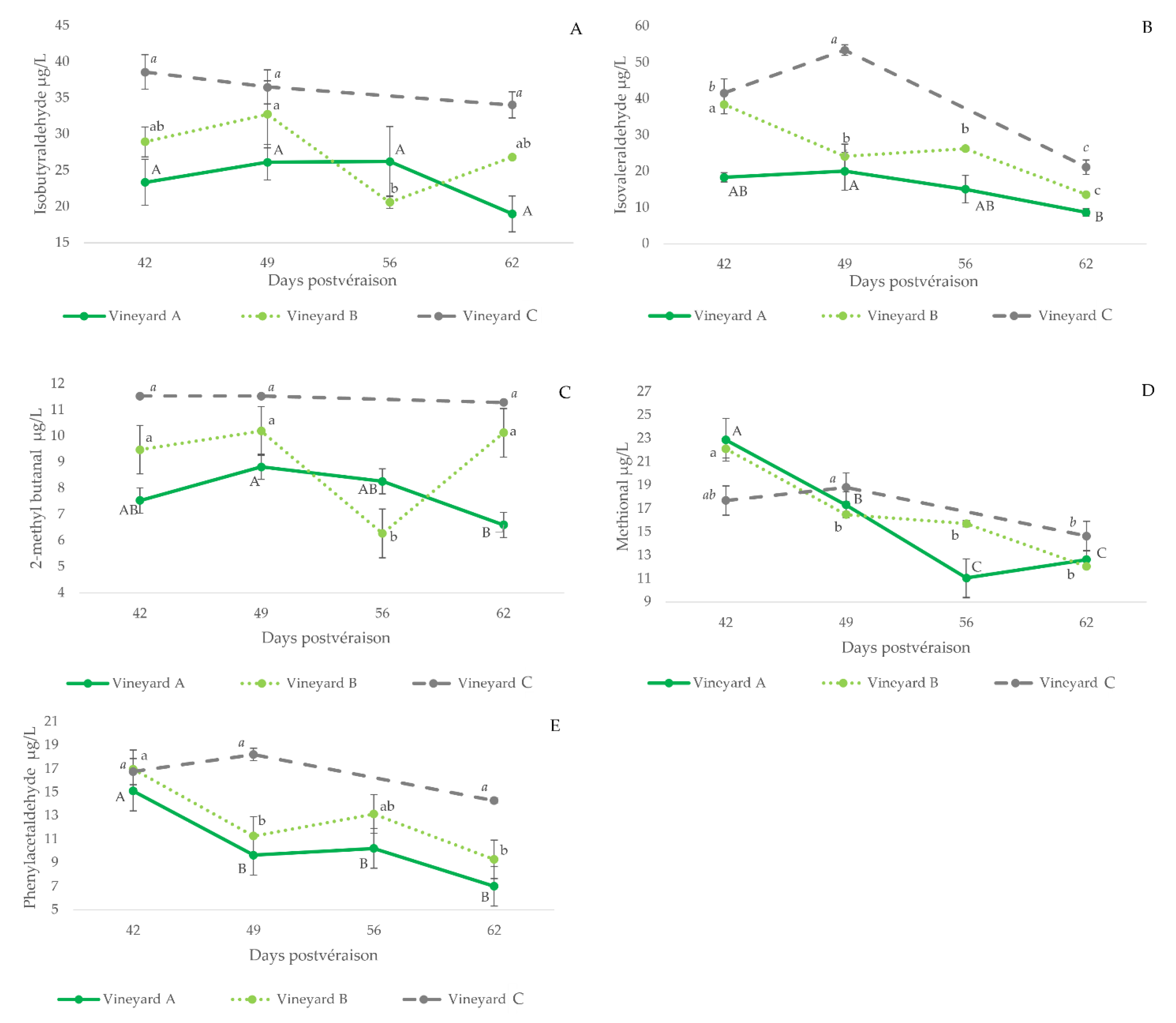
3.2.4. Strecker Aldehydes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernández-Orte, P.; Cacho, J.F.; Ferreira, V. Relationship between varietal amino acid profile of grapes and wine aromatic composition. Experiments with model solutions and chemometric study. J. Agric. Food Chem. 2002, 50, 2891–2899. [Google Scholar] [CrossRef]
- Canoura, C.; Kelly, M.T.; Ojeda, H. Effect of irrigation and timing and type of nitrogen application on the biochemical composition of Vitis vinifera L. cv. Chardonnay and Syrah grapeberries. Food Chem. 2018, 241, 171–181. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma—A Review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Rapp, A.; Versini, G. Influence of nitrogen compounds in grapes on aroma compounds of wines. In Proceedings of the International Symposium on Nitrogen in Grapes and Wine, Seattle, WA, USA, 18–19 June 1991; Rantz, J., Ed.; American Society Enology & Viticulture: Davis, CA, USA; pp. 156–164. [Google Scholar]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. 1991. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Martínez-Gil, A.M.; Lorenzo, C.; Lara, J.F.; Pardo, F.; Salinas, M.R. Implications of nitrogen compounds during alcoholic fermentation from some grape varieties at different maturation stages and cultivation systems. Food Chem. 2011, 124, 106–116. [Google Scholar] [CrossRef]
- Rollero, S.; Mouret, J.R.; Bloem, A.; Sanchez, I.; Ortiz-Julien, A.; Sablayrolles, J.M.; Dequin, S.; Camarasa, C. Quantitative C-13-isotope labelling-based analysis to elucidate the influence of environmental parameters on the production of fermentative aromas during wine fermentation. Microb. Biotechnol. 2017, 10, 1649–1662. [Google Scholar] [CrossRef]
- Robinson, A.L.; Boss, P.K.; Solomon, P.S.; Trengove, R.D.; Heymann, H.; Ebeler, S.E. Origins of grape and wine aroma. Part 1. Chemical components and viticultural impacts. Am. J. Enol. Vitic. 2014, 65, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Seeber, R.; Sferlazzo, G.; Leardi, R.; Serra, A.D.; Versini, G. Multivariate data-analysis in classification of musts and wines of the same variety according to vintage year. J. Agric. Food Chem. 1991, 39, 1764–1769. [Google Scholar] [CrossRef]
- Ortega-Heras, M.; Pérez-Magariño, S.; Del-Villar-Garrachón, V.; González-Huerta, C.; Moro González, L.C.; Guadarrama Rodríguez, A.; Villanueva Sánchez, S.; Gallo González, R.; Martín de la Helguera, S. Study of the effect of vintage, maturity degree, and irrigation on the amino acid and biogenic amine content of a white wine from the Verdejo variety. J. Sci. Food Agric. 2014, 94, 2073–2082. [Google Scholar] [CrossRef]
- Bouloumpasi, E.; Soufleros, E.H.; Tsarchopoulos, C.; Biliaderis, C.G. Primary amino acid composition and its use in discrimination of Greek red wines with regard to variety and cultivation region. Vitis 2002, 41, 195–202. [Google Scholar] [CrossRef]
- Etiévant, P.; Schlich, P.; Bouvier, J.C.; Symonds, P.; Bertrand, A. Varietal and geographic classification of french red wines in terms of elements, amino-acids and aromatic alcohols. J. Sci. Food Agric. 1988, 45, 25–41. [Google Scholar] [CrossRef]
- Bouzas-Cid, Y.; Falqué, E.; Orriols, I.; Mirás-Avalos, J.M. Effects of irrigation over three years on the amino acid composition of Treixadura (Vitis vinifera L.) musts and wines, and on the aromatic composition and sensory profiles of its wines. Food Chem. 2018, 240, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Portu, J.; Santamaría, P.; López, R.; Garde-Cerdán, T. Effects on grape amino acid concentration through foliar application of three different elicitors. Int. Food Res. J. 2017, 99, 688–692. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Guitart, A.; Cacho, J. Changes in the concentration of amino acids during the ripening of Vitis vinifera Tempranillo variety from the Denomination d’Origine Somontano (Spain). Am. J. Enol. Vitic. 1999, 50, 144–154. [Google Scholar]
- Kliewer, W.M. Free amino acids and other nitrogenous fractions in wine grapes. J. Food Sci. 1970, 35, 17–21. [Google Scholar] [CrossRef]
- Fang, Y.; Qian, M.C. Quantification of selected aroma-active compounds in Pinot noir wines from different grape maturities. J. Agric. Food Chem. 2006, 54, 8567–8573. [Google Scholar] [CrossRef]
- Arias-Pérez, I.; Ferrero-Del-Teso, S.; Sáenz-Navajas, M.P.; Fernández-Zurbano, P.; Lacau, B.; Astraín, J.; Barón, C.; Ferreira, V.; Escudero, A. Some clues about the changes in wine aroma composition associated to the maturation of “neutral” grapes. Food Chem. 2020, 320, 126610. [Google Scholar] [CrossRef]
- Wilson, B.; Strauss, C.; Williams, P. Changes in free and glycosidically bound monoterpenes in developing muscat grapes. J. Agric. Food Chem. 1984, 32, 919–924. [Google Scholar] [CrossRef]
- Yuan, F.; Qian, M.C. Development of C13-norisoprenoids, carotenoids and other volatile compounds in Vitis vinifera L. Cv. Pinot noir grapes. Food Chem. 2016, 192, 633–641. [Google Scholar] [CrossRef]
- Carrau, F.M.; Medina, K.; Farina, L.; Boido, E.; Henschke, P.A.; Dellacassa, E. Production of fermentation aroma compounds by Saccharomyces cerevisiae wine yeasts: Effects of yeast assimilable nitrogen on two model strains. FEMS Yeast Res. 2008, 8, 1196–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairbairn, S.; McKinnon, A.; Musarurwa, H.T.; Ferreira, A.C.; Bauer, F.F. The Impact of Single Amino Acids on Growth and Volatile Aroma Production by Saccharomyces cerevisiae Strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crépin, L.; Truong, N.M.; Bloem, A.; Sánchez, I.; Dequin, S.; Camarasa, C. Management of Multiple Nitrogen Sources during Wine Fermentation by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2017, 83, e02617-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, C.; Falco, V.; Mendes-Faia, A.; Mendes-Ferreira, A. Nitrogen addition influences formation of aroma compounds, volatile acidity and ethanol in nitrogen deficient media fermented by Saccharomyces cerevisiae wine strains. J. Biosci. Bioeng. 2009, 108, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, G.P. Formation of Strecker aldehydes from polyphenol-derived quinones and alpha-amino acids in a nonenzymic model system. J. Agric. Food Chem. 2006, 54, 1893–1897. [Google Scholar] [CrossRef] [PubMed]
- Grant-Preece, P.; Fang, H.J.; Schmidtke, L.M.; Clark, A.C. Sensorially important aldehyde production from amino acids in model wine systems: Impact of ascorbic acid, erythorbic acid, glutathione and sulphur dioxide. Food Chem. 2013, 141, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.; Carrascón, V.; Ferreira, V. Release and Formation of Oxidation-Related Aldehydes during Wine Oxidation. J. Agric. Food Chem. 2016, 64, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Escudero, A.; Hernández-Orte, P.; Cacho, J.; Ferreira, V. Clues about the role of methional as character impact odorant of some oxidized wines. J. Agric. Food Chem. 2000, 48, 4268–4272. [Google Scholar] [CrossRef]
- Monforte, A.R.; Martins, S.; Ferreira, A.C.S. Strecker Aldehyde Formation in Wine: New Insights into the Role of Gallic Acid, Glucose and Metals in Phenylacetaldehyde Formation. J. Agric. Food Chem. 2018, 66, 2459–2466. [Google Scholar] [CrossRef]
- Culleré, L.; Cacho, J.; Ferreira, V. An assessment of the role played by some oxidation-related aldehydes in wine aroma. J. Agric. Food Chem. 2007, 55, 876–881. [Google Scholar] [CrossRef]
- Vivar-Quintana, A.M.; Santos-Buelga, C.; Francia-Aricha, E.; Rivas-Gonzalo, J.C. Formation of anthocyanin-derived pigments in experimental red wines. Food Sci. Technol. Int. 1999, 5, 347–352. [Google Scholar] [CrossRef]
- de Azevedo, L.C.; Reis, M.M.; Motta, L.F.; da Rocha, G.O.; Silva, L.A.; de Andrade, J.B. Evaluation of the formation and stability of hydroxyalkylsulfonic acids in wines. J. Agric. Food Chem. 2007, 55, 8670–8680. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Bueno, M.; Franco-Luesma, E.; Culleré, L.; Fernández-Zurbano, P. Key changes in wine aroma active compounds during bottle storage of spanish red wines under different oxygen levels. J. Agric. Food Chem. 2014, 62, 10015–10027. [Google Scholar] [CrossRef] [PubMed]
- Denat, M.; Ontañón, I.; Querol, A.; Ferreira, V. The diverse effects of yeast on the aroma of non-sulfite added white wines throughout aging. LWT 2022, 158, 113111. [Google Scholar] [CrossRef]
- Ferrero-del-Teso, S.; Arias, I.; Escudero, A.; Ferreira, V.; Fernández-Zurbano, P.; Sáenz-Navajas, M.-P. Effect of grape maturity on wine sensory and chemical features: The case of Moristel wines. LWT 2020, 118, 108848. [Google Scholar] [CrossRef]
- García, J.; Zheng, W.; Balda, P.; de Toda, F.M. Varietal differences in the sugar content of red grapes at the onset of anthocyanin synthesis. Vitis 2017, 56, 15–18. [Google Scholar] [CrossRef]
- Alegre, Y.; Arias-Pérez, I.; Hernández-Orte, P.; Ferreira, V. Development of a new strategy for studying the aroma potential of winemaking grapes through the accelerated hydrolysis of phenolic and aromatic fractions (PAFs). Int. Food Res. J. 2020, 127, 108728. [Google Scholar] [CrossRef]
- OIV. Compendium of International Methods of Wine and Must Analysis; Office International de la Vigne et du Vin: Paris, France, 2018. [Google Scholar]
- Ortega, C.; López, R.; Cacho, J.; Ferreira, V. Fast analysis of important wine volatile compounds: Development and validation of a new method based on gas chromatographic–flame ionisation detection analysis of dichloromethane microextracts. J. Chromatogr. A 2001, 923, 205–214. [Google Scholar] [CrossRef]
- López, R.; Aznar, M.; Cacho, J.; Ferreira, V. Determination of minor and trace volatile compounds in wine by solid-phase extraction and gas chromatography with mass spectrometric detection. J. Chromatogr. A 2002, 966, 167–177. [Google Scholar] [CrossRef]
- Bueno, M.; Zapata, J.; Ferreira, V. Simultaneous determination of free and bonded forms of odor-active carbonyls in wine using a headspace solid phase microextraction strategy. J. Chromatogr. A 2014, 1369, 33–42. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Ibarz, M.J.; Cacho, J.; Ferreira, V. Amino acid determination in grape juices and wines by HPLC using a modification of the 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate (AQC) method. Chromatographia 2003, 58, 29–35. [Google Scholar] [CrossRef]
- Kliewer, W.M.; Dokoozlian, N.K. Leaf Area/Crop Weight Ratios of Grapevines: Influence on Fruit Composition and Wine Quality. Am. J. Enol. Vitic. 2005, 56, 170–181. [Google Scholar]
- Garde-Cerdán, T.; Lorenzo, C.; Lara, J.F.; Pardo, F.; Ancín-Azpilicueta, C.; Salinas, M.R. Study of the evolution of nitrogen compounds during grape ripening. Application to differentiate grape varieties and cultivated systems. J. Agric. Food Chem. 2009, 57, 2410–2419. [Google Scholar] [CrossRef] [PubMed]
- Bisson, L.F.; Butzke, C.E. Diagnosis and rectification of stuck and sluggish fermentations. Am. J. Enol. Vitic. 2000, 51, 168–177. [Google Scholar]
- Manginot, C.; Sablayrolles, J.M.; Roustan, J.L.; Barre, P. Use of constant rate alcoholic fermentations to compare the effectiveness of different nitrogen sources added during the stationary phase. Enzyme Microb. Technol. 1997, 20, 373–380. [Google Scholar] [CrossRef]
- Bueno, M.; Marrufo-Curtido, A.; Carrascón, V.; Fernández-Zurbano, P.; Escudero, A.; Ferreira, V. Formation and Accumulation of Acetaldehyde and Strecker Aldehydes during Red Wine Oxidation. Front. Chem. 2018, 6, 20. [Google Scholar] [CrossRef]
- Allamy, L.; Darriet, P.; Pons, A. Molecular interpretation of dried-fruit aromas in Merlot and Cabernet Sauvignon musts and young wines: Impact of over-ripening. Food Chem. 2018, 266, 245–253. [Google Scholar] [CrossRef]
- Bloem, A.; Bertrand, A.; Lonvaud-Funel, A.; de Revel, G. Vanillin production from simple phenols by wine-associated lactic acid bacteria. Lett. Appl. Microbiol. 2007, 44, 62–67. [Google Scholar] [CrossRef]
- Sellami, I.; Mall, V.; Schieberle, P. Changes in the Key Odorants and Aroma Profiles of Hamlin and Valencia Orange Juices Not from Concentrate (NFC) during Chilled Storage. J. Agric. Food Chem. 2018, 66, 7428–7440. [Google Scholar] [CrossRef]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Guth, H. Quantitation and Sensory Studies of Character Impact Odorants of Different White Wine Varieties. J. Agric. Food Chem. 1997, 45, 3027–3032. [Google Scholar] [CrossRef]
- Ferreira, V.; Ortín, N.; Escudero, A.; López, R.; Cacho, J. Chemical characterization of the aroma of Grenache rose wines: Aroma extract dilution analysis, quantitative determination, and sensory reconstitution studies. J. Agric. Food Chem. 2002, 50, 4048–4054. [Google Scholar] [CrossRef] [PubMed]
- Gemert, L.J.N.A.H. Compilation of Odour Threshold Values in Air and Water; National Institute for Water Supply: Zeist, The Netherlands, 2003. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year 2016 | Year 2017 | ||||||
|---|---|---|---|---|---|---|---|
| Data | Vineyard | Mistelle | Wine | Data | Vineyard | Mistelle | Wine |
| 12 September 2016 42 days postvéraison | A | G-AT1 | W-AT1.1 | 6 September 2017 41 days postvéraison | C | W-CT1.1 | |
| W-AT1.2 | W-CT1.2 | ||||||
| W-AT1.3 | W-CT1.3 | ||||||
| B | G-BT1 | W-BT1.1 | |||||
| W-BT1.2 | |||||||
| W-BT1.3 | |||||||
| 19 September 2016 49 days postvéraison | A | G-AT2 | W-AT2.1 | 13 September 2017 48 days postvéraison | C | G-CT2 | W-CT2.1 |
| W-AT2.2 | W-CT2.2 | ||||||
| W-AT2.3 | W-CT2.3 | ||||||
| B | G-BT2 | W-BT2.1 | |||||
| W-BT2.2 | |||||||
| W-BT2.3 | |||||||
| 26 September 2016 56 days postvéraison | A | G-AT3 | W-AT3.1 | ||||
| W-AT3.2 | |||||||
| W-AT3.3 | |||||||
| B | G-BT3 | W-BT3.1 | |||||
| W-BT3.2 | |||||||
| W-BT3.3 | |||||||
| 3 October 2016 62 days postvéraison | A | G-AT4 | W-AT4.1 | 26 September 2017 60 days postvéraison | C | G-CT4 | W-CT4.1 |
| W-AT4.2 | W-CT4.2 | ||||||
| W-AT4.3 | W-CT4.3 | ||||||
| B | G-BT4 | W-BT4.1 | |||||
| W-BT4.2 | |||||||
| W-BT4.3 | |||||||
| Same Amino Acid (w) | Isobutanol | Isoamyl Alcohol | Methionol | β-Phenyl Ethanol | Isoamyl Acetate | Butyl Acetate | Isobutyl Acetate | Phenylethyl Acetate | Ethyl Isobutyrate | Ethyl 2- Methyl Butyrate | Ethyl Isovalerate | Isobutyric Acid | Isovalerianic Acid | Isobutyr Aldehyde | Isovaler Aldehyde | 2-Methyl Butanal | Methional | Phenyl Acetaldehyde | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ALA (m) | 0.701 ** | −0.114 | 0.119 | −0.414 | −0.659 ** | 0.379 | 0.499 | 0.283 | −0.134 | −0.660 ** | −0.249 | −0.421 | −0.121 | −0.576 * | 0.496 | 0.517 | 0.472 | −0.009 | 0.382 |
| ARG (m) | 0.468 | −0.185 | −0.064 | −0.290 | −0.436 | 0.413 | 0.294 | 0.466 | 0.152 | −0.718 ** | −0.472 | −0.515 | −0.115 | −0.340 | 0.268 | 0.197 | 0.309 | −0.240 | 0.015 |
| ASN (m) | 0.897 **** | −0.397 | −0.292 | −0.216 | −0.411 | 0.222 | 0.143 | 0.351 | 0.011 | −0.581 * | −0.390 | −0.099 | −0.229 | −0.222 | 0.373 | 0.319 | 0.419 | 0.118 | 0.216 |
| ASP (m) | 0.385 | −0.486 | −0.068 | 0.051 | −0.167 | 0.101 | −0.014 | 0.176 | 0.008 | −0.647 ** | −0.576 * | −0.384 | −0.404 | −0.163 | 0.038 | 0.050 | 0.002 | −0.182 | −0.027 |
| GABA (m) | −0.632 ** | 0.657 ** | 0.291 | −0.580 * | −0.450 | 0.851 *** | 0.678 ** | 0.344 | 0.215 | 0.013 | 0.367 | −0.530 | 0.727 ** | −0.425 | 0.353 | −0.222 | 0.476 | −0.523 | −0.213 |
| GLU (m) | 0.71 3** | 0.171 | 0.049 | −0.831 *** | −0.906 **** | 0.668 ** | 0.861 *** | 0.462 | −0.085 | −0.710 ** | −0.045 | −0.572 * | 0.200 | −0.842 *** | 0.568 * | 0.410 | 0.662 ** | −0.189 | 0.310 |
| GLY (m) | 0.735 ** | −0.380 | 0.188 | 0.219 | −0.047 | 0.030 | −0.108 | 0.095 | 0.016 | −0.573 * | −0.617 * | −0.282 | −0.465 | −0.072 | −0.054 | 0.169 | −0.135 | −0.085 | 0.103 |
| HIS (m) | 0.438 | −0.115 | −0.159 | −0.456 | −0.501 | 0.599 * | 0.393 | 0.604 * | 0.265 | −0.766 *** | −0.436 | −0.575 * | 0.047 | −0.381 | 0.320 | −0.028 | 0.423 | −0.633 ** | −0.135 |
| ILE (m) | 0.687 ** | 0.006 | −0.069 | −0.520 | −0.509 | 0.625 * | 0.489 | 0.529 | 0.198 | −0.807 *** | −0.449 | −0.791 *** | 0.020 | −0.547 * | 0.260 | −0.154 | 0.365 | −0.789 *** | −0.248 |
| LEU (m) | 0.682 ** | −0.096 | −0.074 | −0.473 | −0.536 * | 0.560 * | 0.459 | 0.499 | 0.137 | −0.832 *** | −0.472 | −0.720 ** | −0.060 | −0.548 * | 0.291 | −0.006 | 0.372 | −0.719 ** | −0.123 |
| LYS (m) | 0.780 *** | −0.322 | 0.185 | −0.005 | −0.277 | 0.101 | 0.124 | 0.099 | −0.130 | −0.715 ** | −0.575 * | −0.406 | −0.561 * | −0.345 | 0.087 | 0.329 | 0.018 | −0.092 | 0.230 |
| MET (m) | 0.535 | −0.219 | −0.054 | −0.018 | −0.057 | 0.295 | −0.034 | 0.454 | 0.340 | −0.757 ** | −0.779 *** | −0.521 | −0.305 | −0.071 | −0.091 | −0.139 | −0.051 | −0.468 | −0.272 |
| ORN (m) | 0.218 | −0.411 | 0.168 | −0.051 | −0.457 | −0.229 | 0.200 | −0.358 | −0.797 *** | −0.306 | −0.038 | 0.070 | −0.516 | −0.465 | 0.469 | 0.764 ** | 0.277 | 0.500 | 0.765 *** |
| PHE (m) | 0.586 * | −0.062 | 0.295 | −0.050 | −0.211 | 0.187 | 0.192 | 0.125 | −0.025 | −0.661 ** | −0.547 | −0.696 ** | −0.354 | −0.376 | 0.015 | 0.139 | −0.048 | −0.420 | −0.035 |
| PRO (m) | 0.676 ** | 0.294 | 0.078 | −0.658 ** | −0.584 * | 0.798 *** | 0.665 ** | 0.567 * | 0.258 | −0.587 * | −0.160 | −0.866 *** | 0.359 | −0.566 * | 0.353 | −0.154 | 0.491 | −0.791 *** | −0.280 |
| SER (m) | 0.733 ** | −0.272 | −0.148 | −0.046 | −0.125 | 0.334 | −0.006 | 0.513 | 0.357 | −0.707 ** | −0.697 ** | −0.459 | −0.210 | −0.050 | −0.008 | −0.085 | 0.062 | −0.365 | −0.243 |
| THR + NH4 (m) | 0.697 ** | −0.307 | −0.074 | −0.033 | −0.324 | 0.043 | 0.091 | 0.113 | −0.141 | −0.451 | −0.368 | −0.302 | −0.365 | −0.244 | 0.403 | 0.443 | 0.344 | 0.043 | 0.188 |
| TYR (m) | 0.820 *** | −0.290 | 0.094 | −0.107 | −0.353 | 0.189 | 0.195 | 0.189 | −0.077 | −0.712 ** | −0.526 | −0.490 | −0.379 | −0.374 | 0.198 | 0.294 | 0.149 | −0.165 | 0.147 |
| VAL (m) | 0.691 ** | −0.106 | −0.123 | −0.440 | −0.512 | 0.537 | 0.413 | 0.502 | 0.154 | −0.779 *** | −0.465 | −0.714 ** | −0.061 | −0.470 | 0.343 | −0.001 | 0.422 | −0.694 ** | −0.150 |
| Same amino acid (m) | isobutanol | isoamyl alcohol | methionol | β-phenyl ethanol | isoamyl acetate | butyl acetate | isobutyl acetate | phenylethyl acetate | ethyl isobutyrate | ethyl 2- methyl butyrate | ethyl isovalerate | isobutyric acid | isovalerianic acid | isobutyr aldehyde | isovaler aldehyde | 2-methyl butanal | methional | phenyl acetaldehyde | |
| ALA (m-w) | 0.954 **** | 0.042 | 0.252 | −0.481 | −0.806 *** | 0.369 | 0.685 ** | 0.141 | −0.274 | −0.544 | −0.088 | −0.514 | −0.084 | −0.705 ** | 0.884 *** | 0.559 * | 0.905 **** | −0.072 | 0.415 |
| ARG (m-w) | 0.999 **** | −0.197 | −0.067 | −0.260 | −0.403 | 0.403 | 0.260 | 0.472 | 0.173 | −0.722 ** | −0.501 | −0.503 | −0.130 | −0.308 | 0.236 | 0.180 | 0.650 * | −0.243 | −0.001 |
| ASN (m-w) | 0.264 | 0.052 | −0.397 | −0.671 ** | −0.614 * | 0.089 | 0.540 | −0.040 | −0.428 | −0.213 | 0.232 | −0.425 | −0.044 | −0.664 ** | 0.693 ** | 0.238 | 0.681 ** | −0.182 | 0.050 |
| ASP (m-w) | 0.817 *** | −0.551 | 0.072 | 0.451 | 0.188 | −0.312 | −0.343 | −0.293 | −0.201 | −0.209 | −0.396 | −0.146 | −0.560 * | 0.095 | −0.097 | −0.061 | −0.265 | −0.061 | −0.045 |
| GABA (m-w) | 0.944 **** | 0.659 * | 0.202 | −0.447 | −0.209 | 0.551 * | 0.483 | 0.254 | 0.261 | 0.187 | 0.400 | −0.406 | 0.704 ** | −0.227 | 0.154 | −0.399 | 0.275 | −0.602 * | −0.362 |
| GLU (m-w) | 0.966 **** | 0.292 | 0.122 | −0.939 **** | −0.931 **** | 0.666 ** | 0.944 **** | 0.363 | −0.166 | −0.601 * | 0.102 | −0.640 ** | 0.269 | −0.920 **** | 0.723 ** | 0.371 | 0.884 *** | −0.263 | 0.284 |
| GLY (m-w) | −0.339 | 0.191 | 0.449 | 0.352 | 0.274 | −0.511 | −0.158 | −0.808 *** | −0.506 | 0.683 ** | 0.497 | 0.176 | −0.118 | 0.055 | 0.040 | 0.034 | −0.198 | 0.131 | 0.145 |
| HIS (m-w) | 0.890 **** | 0.169 | −0.117 | −0.526 | −0.412 | 0.835 *** | 0.427 | 0.841 *** | 0.595 * | −0.710 ** | −0.333 | −0.559 * | 0.418 | −0.228 | 0.204 | −0.272 | 0.401 | −0.564 * | −0.344 |
| ILE (m-w) | 0.700 ** | 0.031 | −0.003 | −0.137 | −0.039 | 0.232 | 0.125 | 0.091 | 0.088 | −0.358 | −0.309 | −0.688 ** | −0.099 | −0.235 | −0.004 | −0.526 | 0.005 | −0.778 *** | −0.533 |
| LEU (m-w) | 0.736 ** | 0.081 | −0.018 | −0.246 | −0.145 | 0.451 | 0.220 | 0.349 | 0.284 | −0.465 | −0.357 | −0.754 ** | 0.067 | −0.234 | 0.054 | −0.501 | 0.115 | −0.810 *** | −0.557 * |
| LYS (m-w) | −0.058 | 0.136 | 0.325 | 0.633 ** | 0.749 ** | −0.194 | −0.573 * | −0.116 | 0.375 | 0.216 | −0.337 | −0.107 | −0.130 | 0.614 * | −0.628 * | −0.584 * | −0.690 ** | −0.440 | −0.559 * |
| MET (m-w) | 0.784 *** | 0.203 | 0.206 | −0.100 | 0.028 | 0.666 ** | 0.091 | 0.638* | 0.625 * | −0.537 | −0.555 * | −0.680 ** | 0.057 | −0.050 | −0.207 | −0.398 | −0.154 | −0.806 *** | −0.490 |
| ORN (m-w) | 0.63 3** | −0.373 | 0.207 | 0.403 | 0.088 | −0.792 *** | −0.235 | −0.921 *** | −0.879 *** | 0.279 | 0.183 | 0.212 | −0.685 ** | −0.119 | 0.188 | 0.490 | −0.141 | 0.455 | 0.496 |
| PHE (m-w) | 0.347 | 0.258 | 0.366 | 0.334 | 0.443 | −0.071 | −0.223 | −0.147 | 0.203 | 0.200 | −0.085 | −0.363 | 0.035 | 0.235 | −0.393 | −0.470 | −0.474 | −0.521 | −0.500 |
| PRO (m-w) | −0.317 | 0.120 | 0.364 | 0.267 | 0.092 | −0.532 | −0.048 | −0.856 *** | −0.634 ** | 0.762 *** | 0.729 ** | 0.210 | −0.049 | −0.028 | 0.274 | 0.275 | 0.015 | 0.331 | 0.298 |
| SER (m-w) | 0.984 **** | −0.236 | −0.133 | 0.017 | −0.037 | 0.269 | −0.073 | 0.447 | 0.354 | −0.635 ** | −0.697 ** | −0.496 | −0.244 | 0.001 | −0.022 | −0.152 | 0.021 | −0.440 | −0.331 |
| THR + NH4 (m-w) | 0.982 **** | −0.248 | 0.001 | −0.025 | −0.345 | −0.022 | 0.125 | −0.021 | −0.262 | −0.345 | −0.257 | −0.331 | −0.376 | −0.299 | 0.698 ** | 0.504 | 0.660 * | 0.052 | 0.227 |
| TYR (m-w) | 0.606 * | −0.081 | 0.310 | 0.335 | 0.208 | −0.055 | −0.197 | −0.089 | 0.072 | −0.169 | −0.381 | −0.449 | −0.284 | 0.078 | −0.114 | −0.165 | −0.249 | −0.401 | −0.271 |
| VAL (m-w) | 0.871 **** | 0.009 | −0.043 | −0.362 | −0.396 | 0.422 | 0.370 | 0.265 | 0.047 | −0.507 | −0.271 | −0.757 ** | −0.023 | −0.440 | 0.388 | −0.185 | 0.595 * | −0.636 ** | −0.307 |
| ARG (m) | THR (m) | ALA (m) | GABA (m) | SER (m) | GLU (m) | THR/GABA (m) | ALA/GABA (m) | GLU/GABA (m) | SER/GABA (m) | (SER + GLU)/GABA (m) | (ALA + SER)/GABA (m) | (ALA + GLU)/GABA (m) | Same Aa/GABA (m) | Same Aa (m) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ASN (w) | 0.518 ** | 0.580 *** | 0.448 ** | 0.192 | 0.587 *** | 0.088 | 0.624 *** | 0.632 *** | 0.235 | 0.678 *** | 0.532 ** | 0.702 *** | 0.507 ** | 0.801 **** | 0.787 **** |
| GABA (w) | 0.424 ** | 0.692 *** | 0.542 ** | 0.401 ** | 0.345 * | 0.113 | 0.851 **** | 0.895 **** | 0.364 * | 0.590 *** | 0.615 *** | 0.854 **** | 0.739 **** | - | 0.401 ** |
| GLU (w) | 0.609 *** | 0.469 ** | 0.640 *** | 0.059 | 0.479 ** | 0.507 ** | 0.402 ** | 0.631 *** | 0.727 **** | 0.451 ** | 0.826 **** | 0.617 *** | 0.765 **** | 0.727 *** | 0.507 ** |
| GLY (w) | 0.724 *** | 0.500 ** | 0.518 ** | 0.072 | 0.783 **** | 0.285 | 0.434 ** | 0.547 ** | 0.452 ** | 0.717 *** | 0.755 *** | 0.652 *** | 0.578 *** | 0.429 ** | 0.487 ** |
| HIS (w) | 0.563 ** | 0.709 *** | 0.613 *** | 0.297 | 0.484 ** | 0.223 | 0.812 **** | 0.895 **** | 0.510 ** | 0.701 *** | 0.798 **** | 0.899 **** | 0.825 **** | 0.757 **** | 0.356 * |
| ILE (w) | 0.730 *** | 0.463 ** | 0.768 **** | 0.005 | 0.502 ** | 0.765 **** | 0.268 | 0.518 ** | 0.826 **** | 0.330 * | 0.810 **** | 0.489 ** | 0.726 *** | 0.469 ** | 0.478 ** |
| LEU (w) | 0.597 *** | 0.554 ** | 0.772 **** | 0.058 | 0.367 * | 0.627 *** | 0.479 ** | 0.757 **** | 0.881 **** | 0.381 * | 0.885 **** | 0.671 *** | 0.921 **** | 0.516 ** | 0.419 ** |
| LYS (w) | 0.571 ** | 0.595 *** | 0.807 **** | 0.058 | 0.320 * | 0.614 *** | 0.530 ** | 0.807 **** | 0.866 **** | 0.363 * | 0.862 **** | 0.696 *** | 0.948 **** | 0.566 ** | 0.619 *** |
| MET (w) | 0.317 * | 0.570 ** | 0.314 * | 0.476 ** | 0.340 * | 0.058 | 0.786 **** | 0.667 *** | 0.269 | 0.607 *** | 0.534 ** | 0.702 *** | 0.549 ** | 0.448 ** | 0.255 |
| PHE (w) | 0.624 *** | 0.620 *** | 0.774 **** | 0.075 | 0.435 ** | 0.563 ** | 0.569 ** | 0.812 **** | 0.837 **** | 0.480 ** | 0.925 **** | 0.752 *** | 0.938 **** | 0.415 ** | 0.352 * |
| THR (w) | 0.602 *** | 0.500 ** | 0.421 ** | 0.179 | 0.701 *** | 0.164 | 0.518 ** | 0.564 ** | 0.350 * | 0.745 *** | 0.675 *** | 0.674 *** | 0.536 ** | 0.518 ** | 0.500 ** |
| TYR (w) | 0.662 *** | 0.603 *** | 0.793 **** | 0.063 | 0.460 ** | 0.604 *** | 0.535 ** | 0.800 **** | 0.865 **** | 0.483 ** | 0.947 **** | 0.745 **** | 0.943 **** | 0.643 *** | 0.680 *** |
| VAL (w) | 0.660 *** | 0.474 ** | 0.624 *** | 0.057 | 0.567 ** | 0.482 ** | 0.402 ** | 0.615 *** | 0.694 *** | 0.524 ** | 0.847 **** | 0.634 *** | 0.738 **** | 0.560 ** | 0.470 ** |
| ALA (w) | 0.599 *** | 0.486 ** | 0.507 ** | 0.190 | 0.603 *** | 0.232 | 0.505 ** | 0.652 *** | 0.456 ** | 0.667 *** | 0.735 *** | 0.713 *** | 0.647 *** | 0.652 *** | 0.507 ** |
| ASP (w) | 0.504 ** | 0.231 | 0.470 ** | 0.011 | 0.361 * | 0.677 *** | 0.107 | 0.273 | 0.704 *** | 0.200 | 0.626 *** | 0.269 | 0.486 ** | 0.068 | 0.143 |
| SER (w) | 0.588 *** | 0.383 * | 0.552 ** | 0.031 | 0.543 ** | 0.418 ** | 0.322 * | 0.528 ** | 0.578 *** | 0.480 ** | 0.732 *** | 0.555 ** | 0.626 *** | 0.480 ** | 0.543 ** |
| ARG (w) | 0.055 | 0.032 | 0.200 | 0.299 | 0.008 | 0.447 ** | 0.003 | 0.017 | 0.249 | 0.069 | 0.046 | 0.000 | 0.091 | 0.014 | 0.055 |
| ORN (w) | 0.306 * | 0.091 | 0.276 | 0.052 | 0.249 | 0.458 ** | 0.022 | 0.112 | 0.403 ** | 0.093 | 0.340 * | 0.115 | 0.240 | 0.000 | 0.001 |
| PRO (w) | 0.468 ** | 0.133 | 0.257 | 0.041 | 0.547 ** | 0.376 * | 0.046 | 0.116 | 0.340 * | 0.290 | 0.436 ** | 0.179 | 0.221 | 0.561 ** | 0.469 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arias-Pérez, I.; Ontañón, I.; Ferreira, V.; Escudero, A. Maturation of Moristel in Different Vineyards: Amino Acid and Aroma Composition of Mistelles and Wines with Particular Emphasis in Strecker Aldehydes. Foods 2022, 11, 958. https://doi.org/10.3390/foods11070958
Arias-Pérez I, Ontañón I, Ferreira V, Escudero A. Maturation of Moristel in Different Vineyards: Amino Acid and Aroma Composition of Mistelles and Wines with Particular Emphasis in Strecker Aldehydes. Foods. 2022; 11(7):958. https://doi.org/10.3390/foods11070958
Chicago/Turabian StyleArias-Pérez, Ignacio, Ignacio Ontañón, Vicente Ferreira, and Ana Escudero. 2022. "Maturation of Moristel in Different Vineyards: Amino Acid and Aroma Composition of Mistelles and Wines with Particular Emphasis in Strecker Aldehydes" Foods 11, no. 7: 958. https://doi.org/10.3390/foods11070958
APA StyleArias-Pérez, I., Ontañón, I., Ferreira, V., & Escudero, A. (2022). Maturation of Moristel in Different Vineyards: Amino Acid and Aroma Composition of Mistelles and Wines with Particular Emphasis in Strecker Aldehydes. Foods, 11(7), 958. https://doi.org/10.3390/foods11070958