
Proteomic Response of Bacillus subtilis Spores under High Pressure Combined with Moderate Temperature and Random Peptide Mixture LK Treatment


Abstract
:1. Introduction
2. Materials and Methods
2.1. B. subtilis Strain and Sporulation
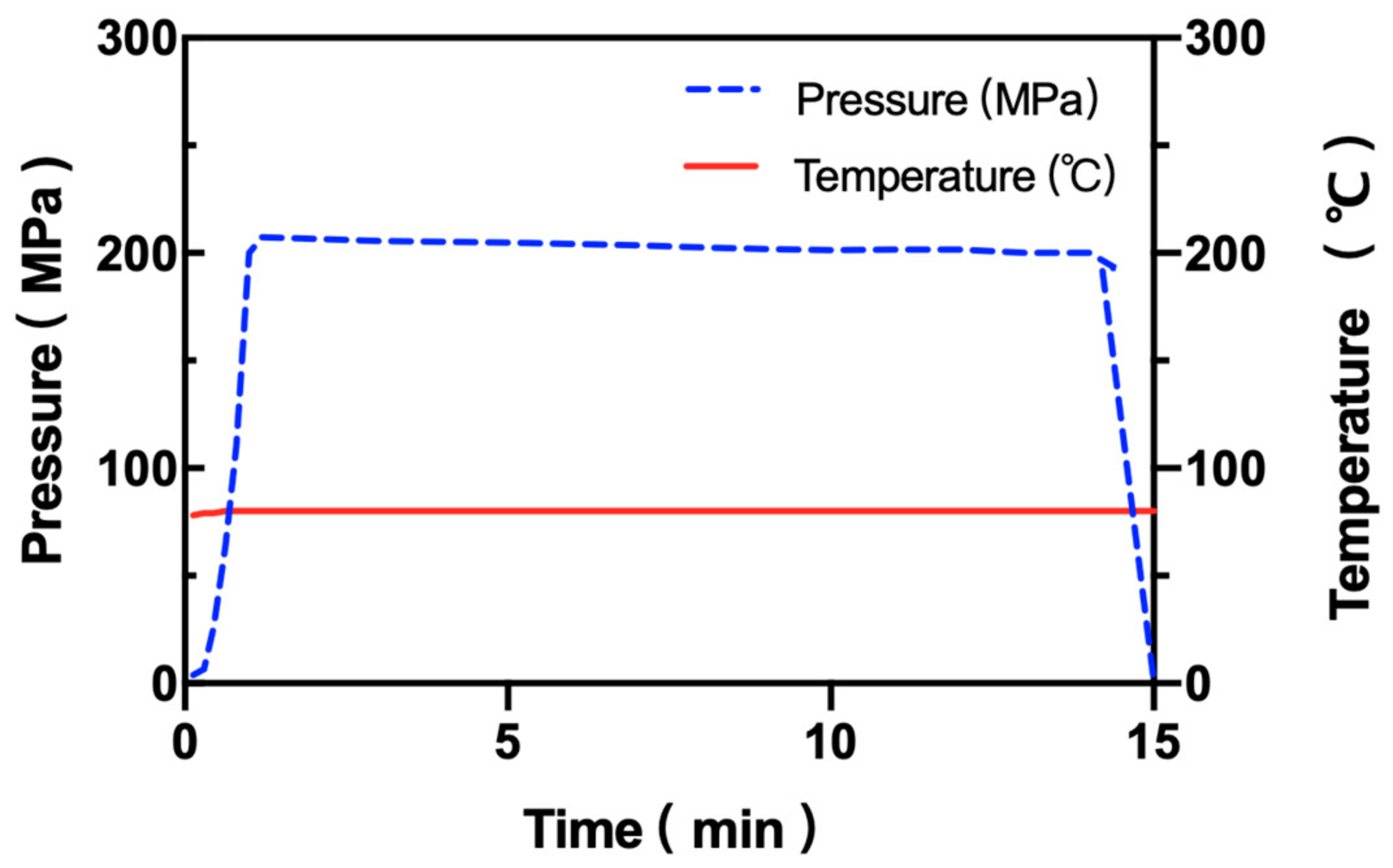
2.2. HPP Equipment and Treatment
2.3. RPM Synthesis
2.4. Spore Enumeration
2.5. Microscope Analysis
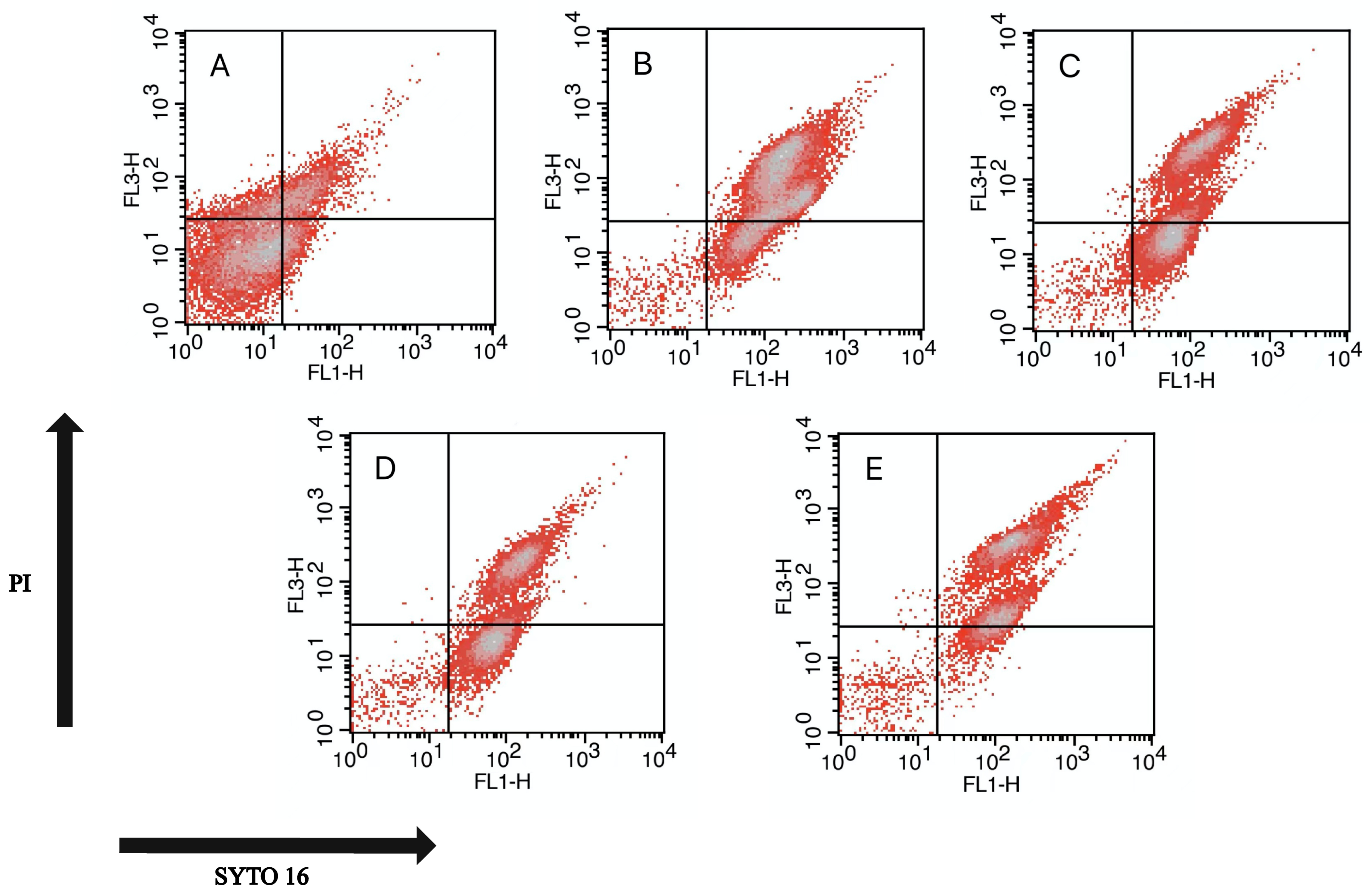
2.6. Flow Cytometry (FCM) Analysis
2.7. Protein Extraction, Digestion, and TMT Quantitation
2.8. Peptide Fractionation
2.9. LC–MS/MS Analysis
2.10. Data Analysis
3. Results
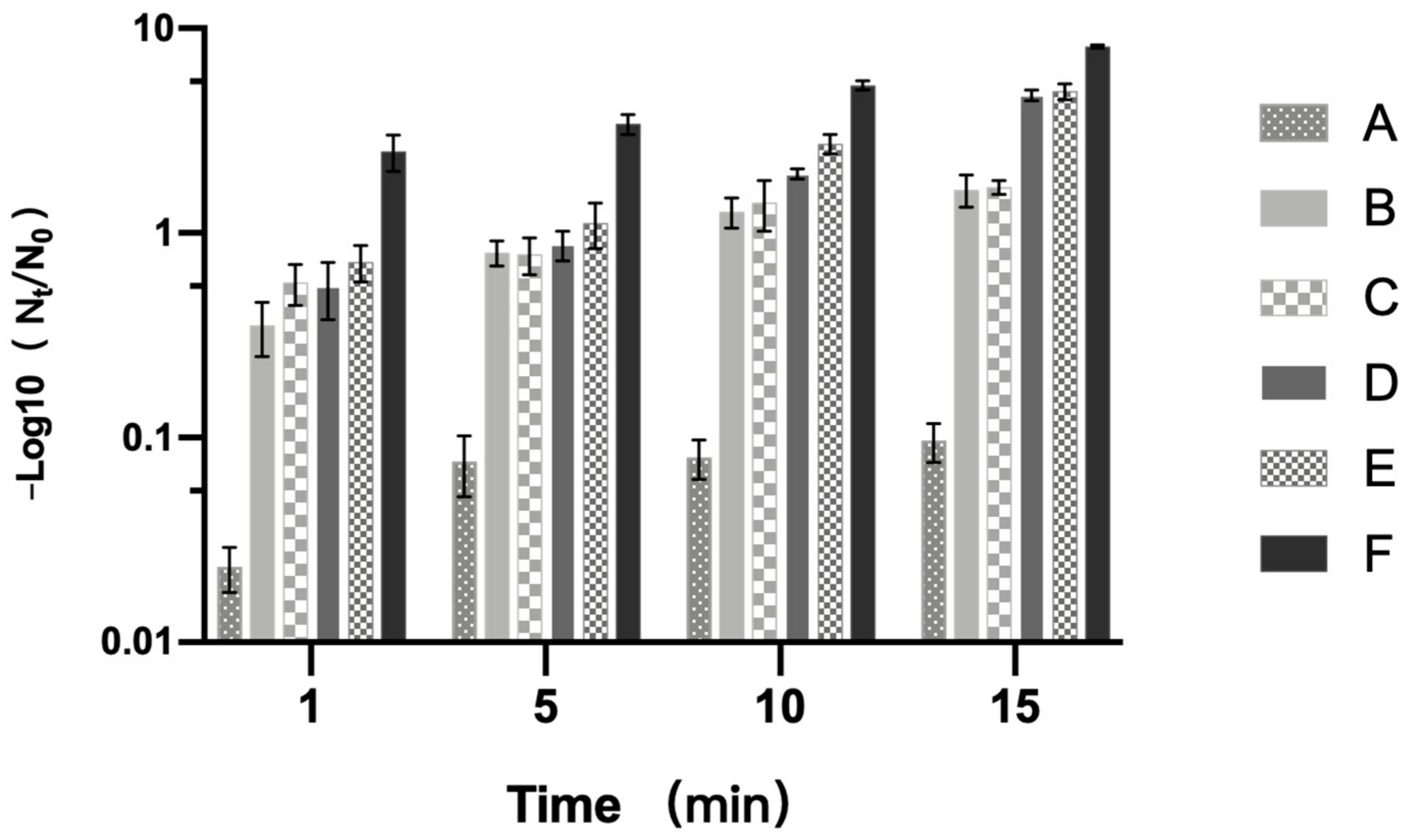
3.1. Inactivation Effect of PT-LK on B. subtilis Spores
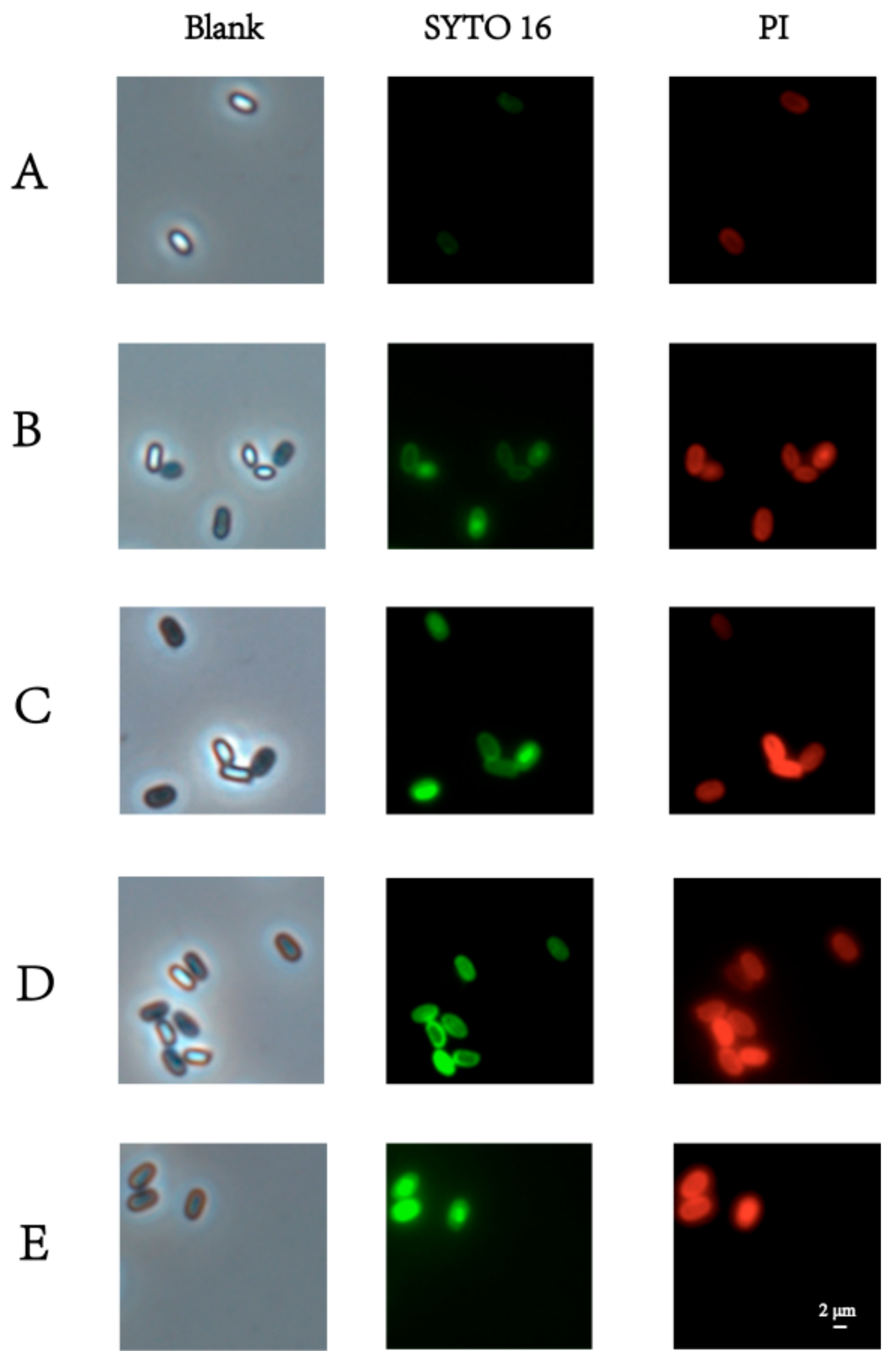
3.2. Morphological Changes in Spores in Response to PT-LK
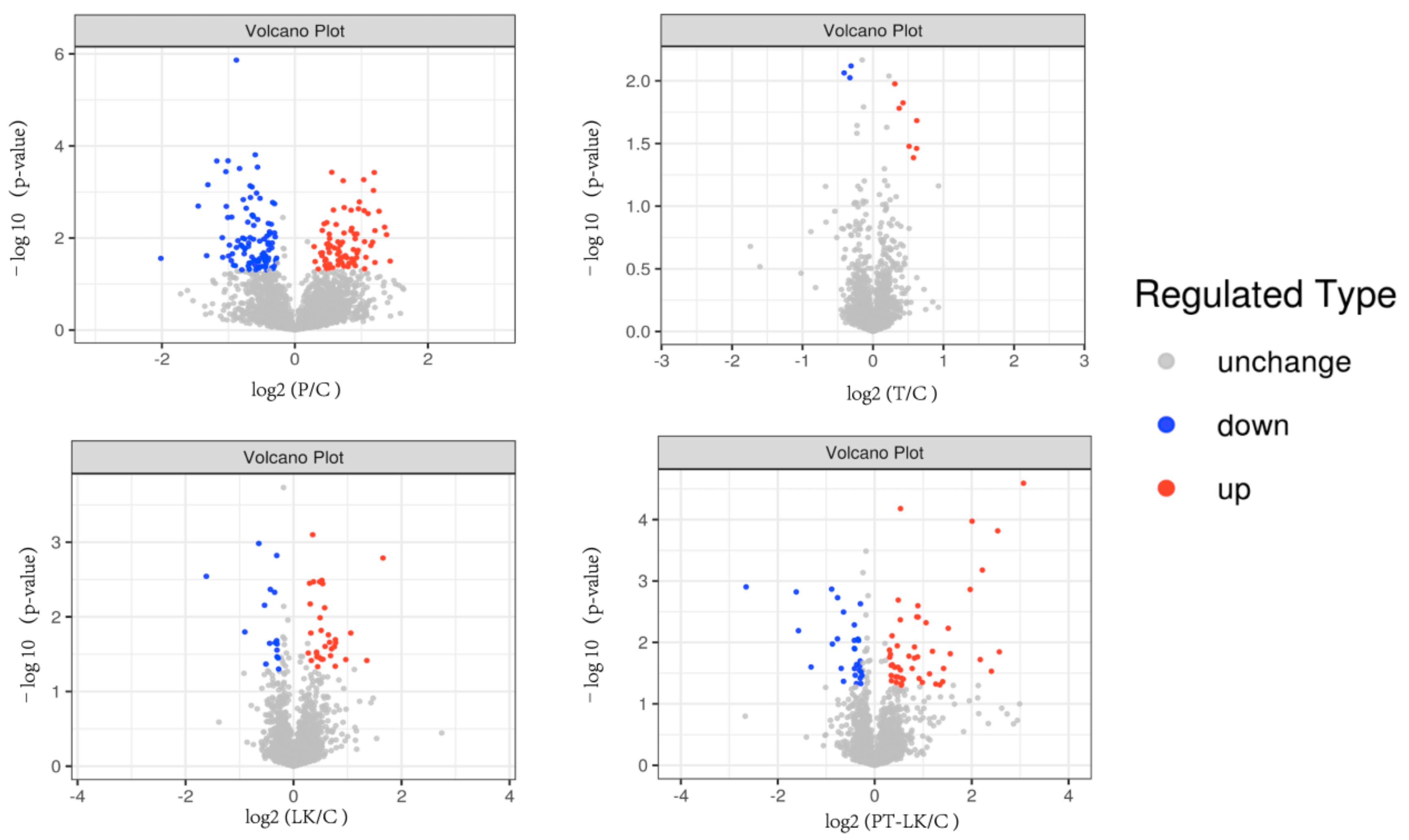
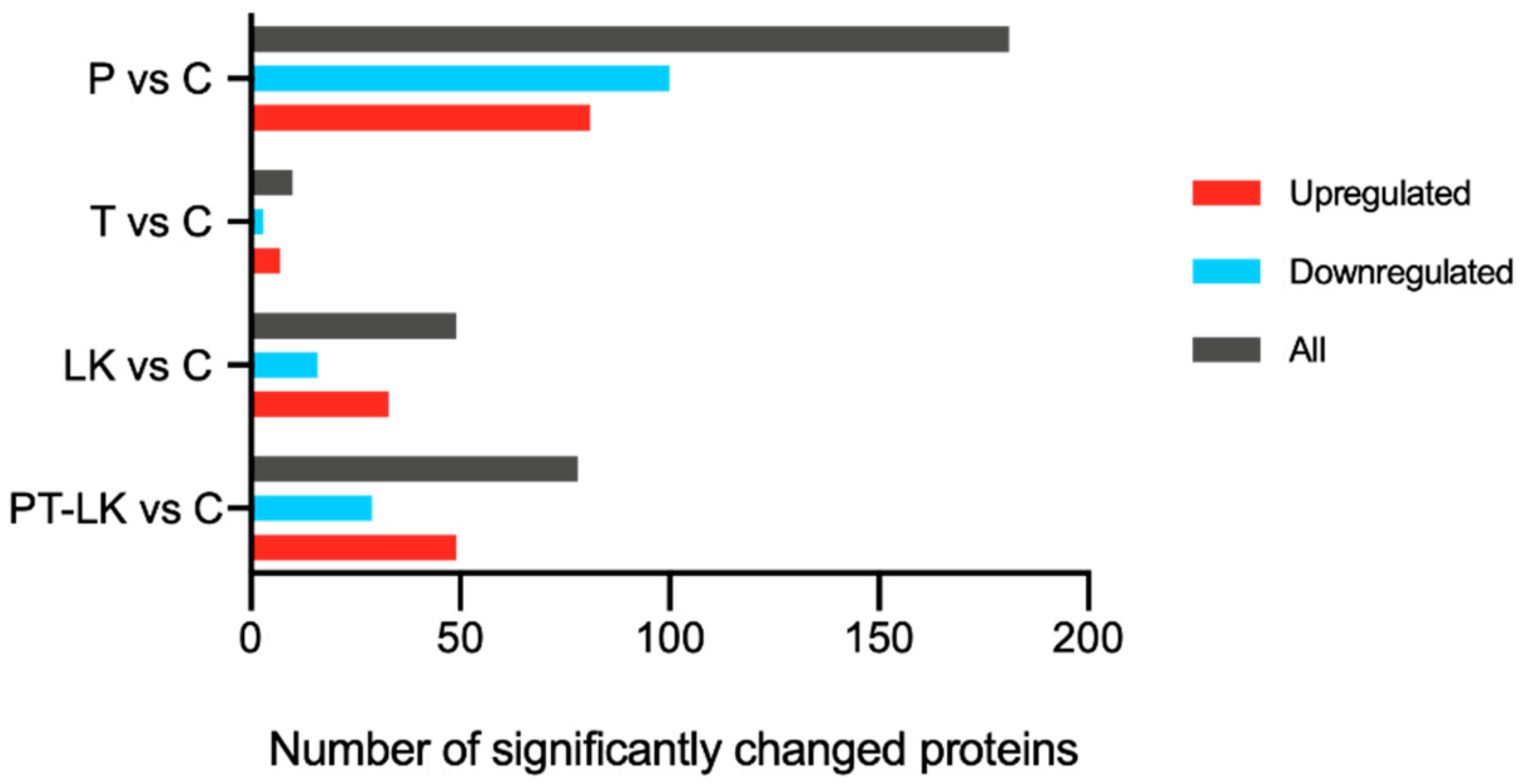
3.3. TMT Proteomic Analysis of B. subtilis Spores in Response to PT-LK
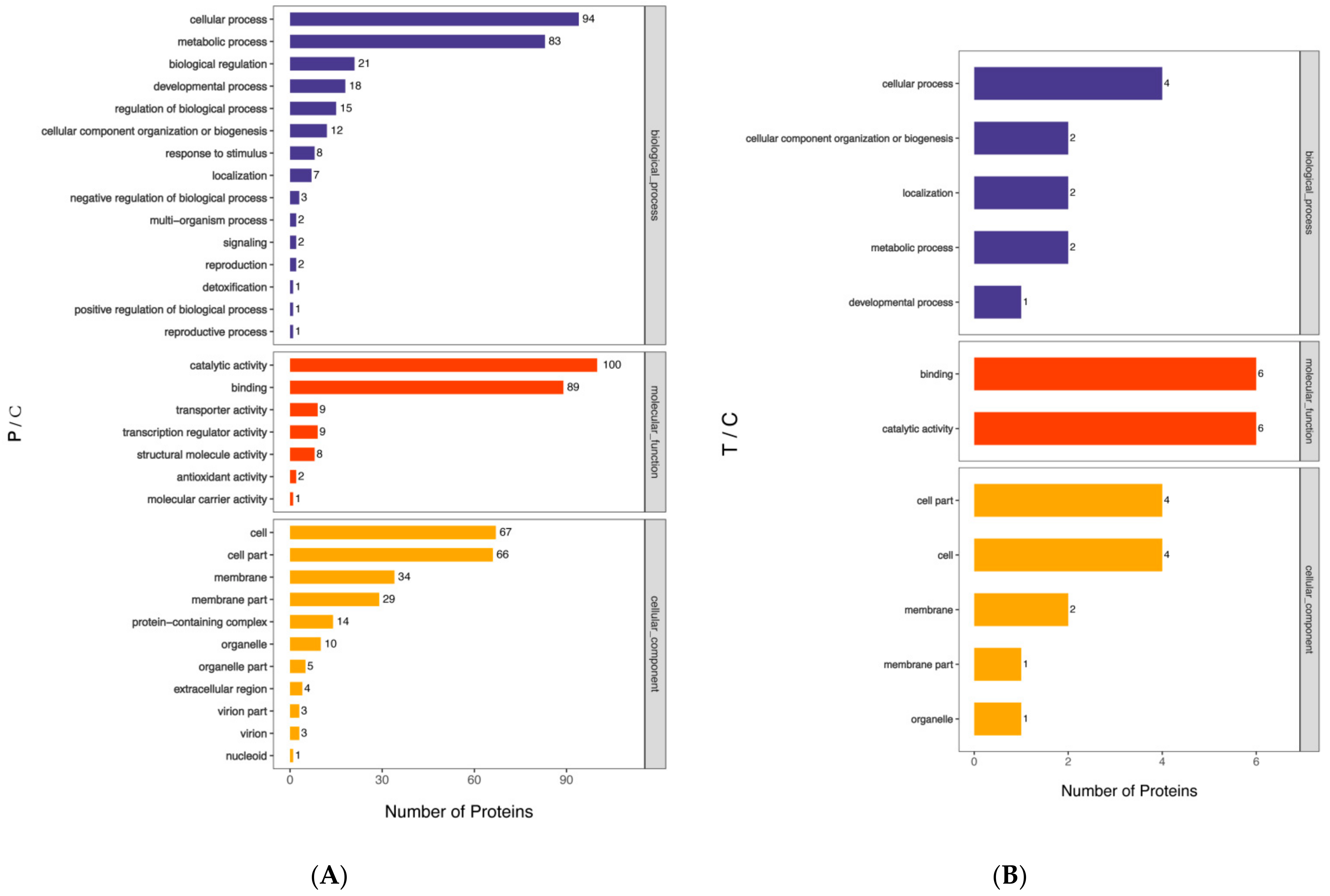
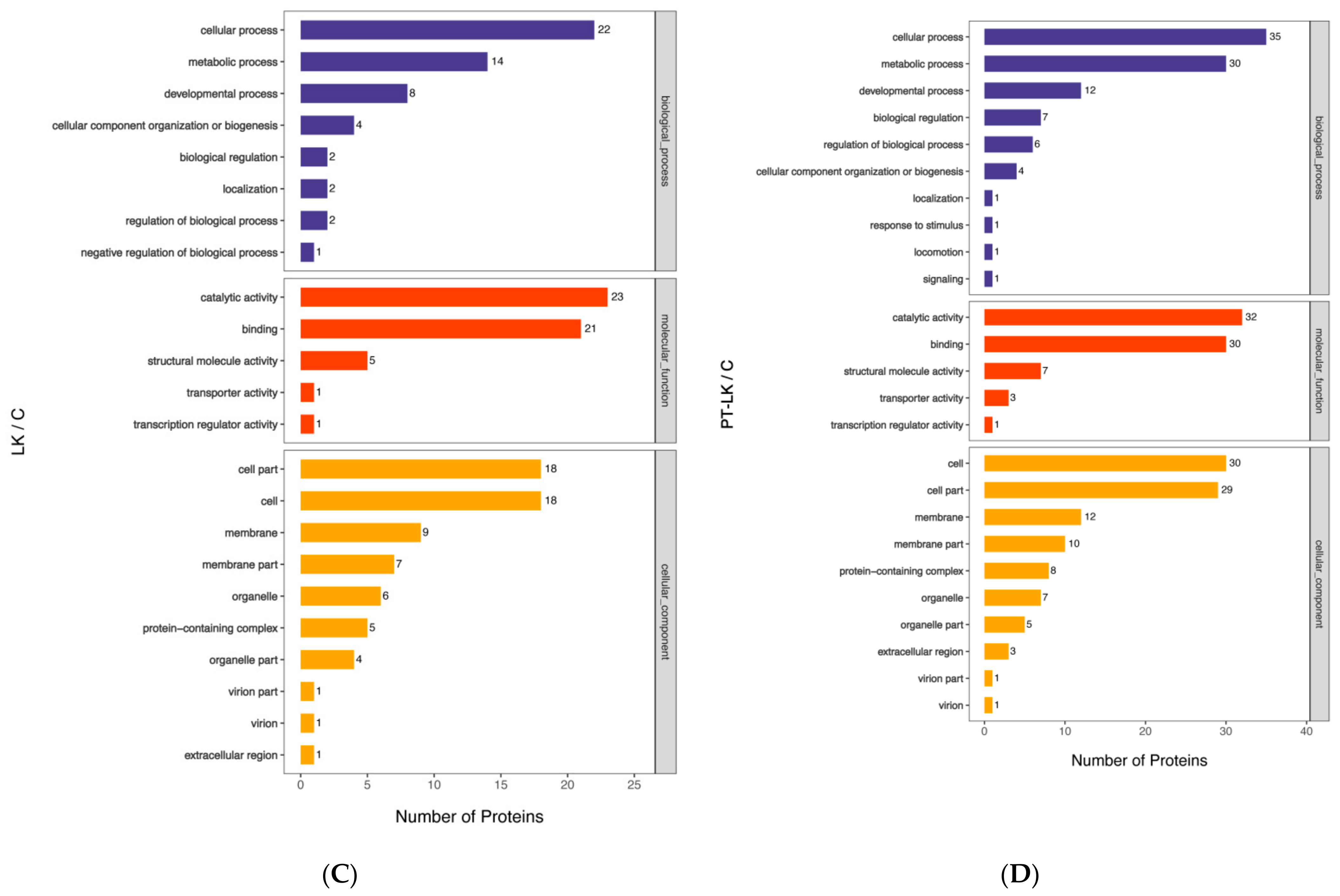
3.4. Gene Ontology (GO) Analysis of B. subtilis Spore Proteins Induced by PT-LK
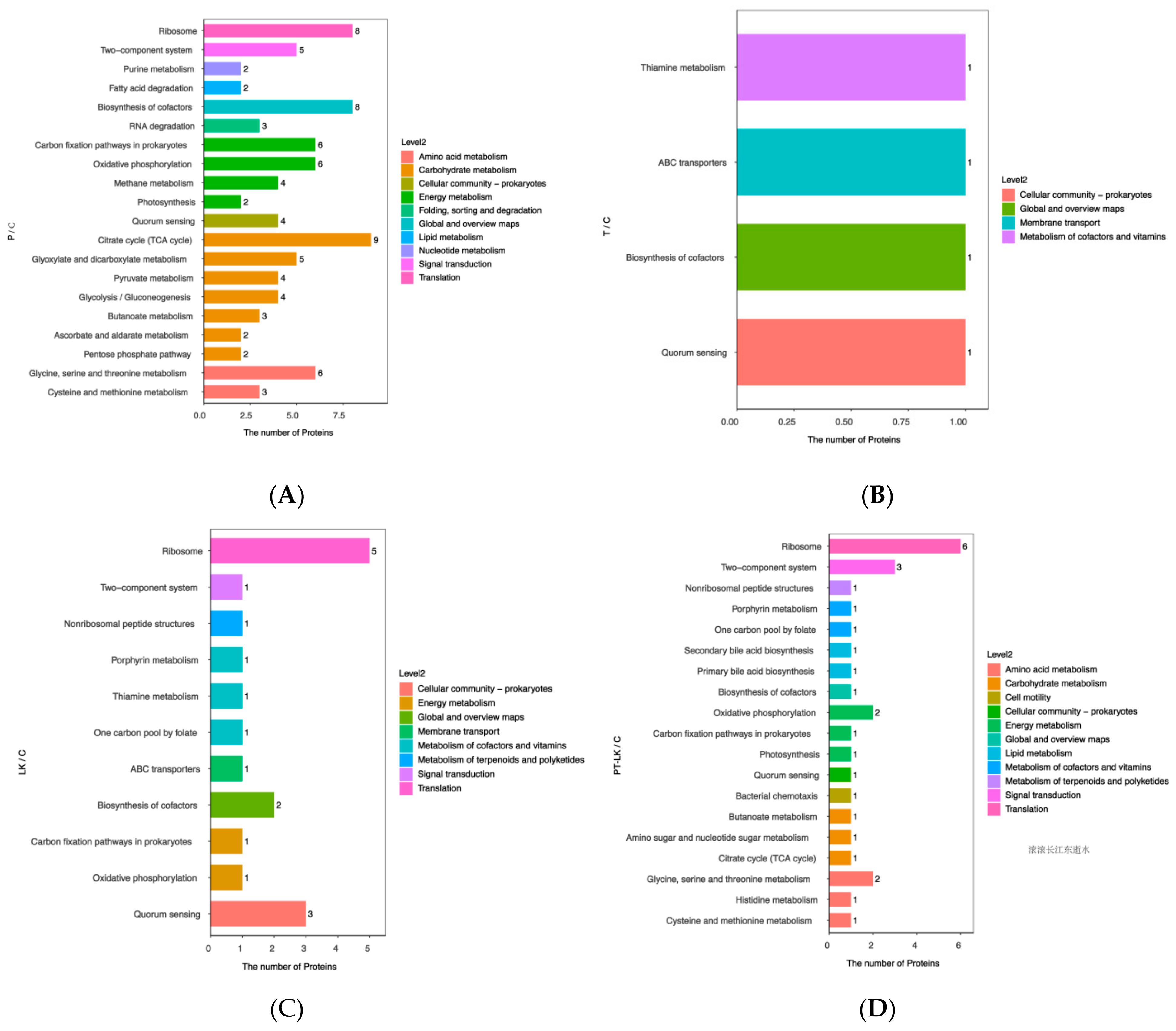
3.5. KEGG Pathway Analysis of DEPs after Different Treatments
3.5.1. Roles of DEPs in P/C Regulation
3.5.2. Roles of DEPs in T/C Regulation
3.5.3. Roles of DEPs in LK/C Regulation
3.5.4. Roles of DEPs in PT-LK/C Regulation
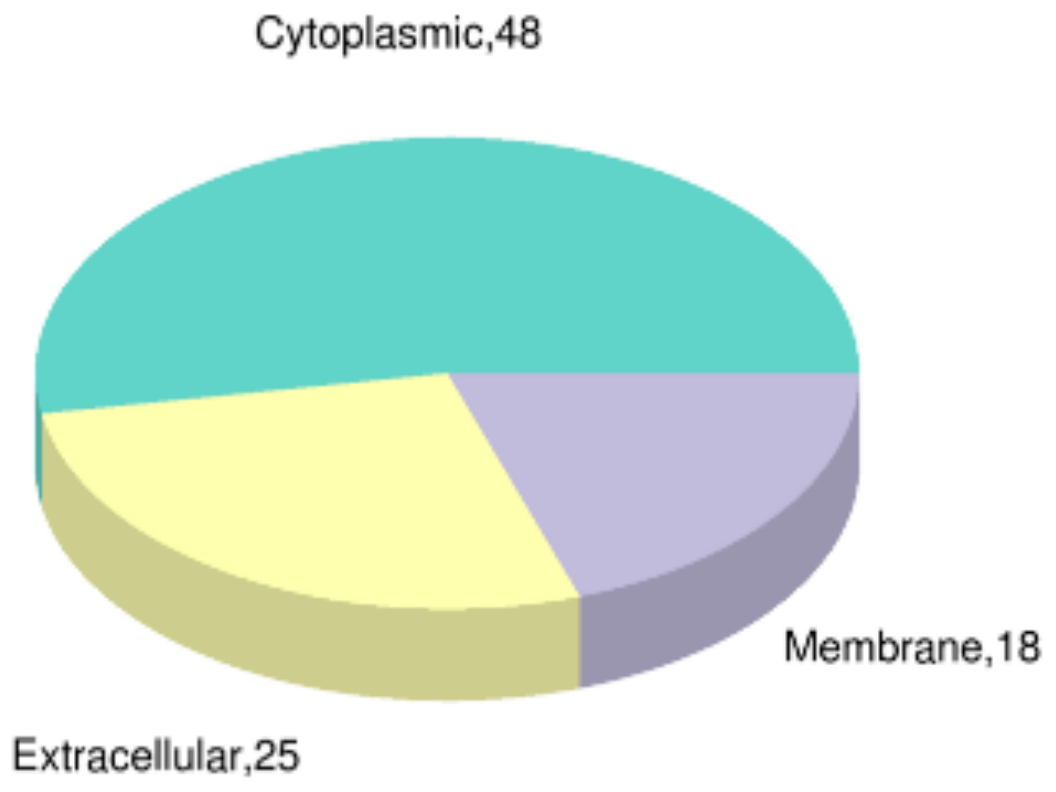
3.6. Subcellular Localization of DEPs after PT-LK Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sevenich, R.; Rauh, C.; Belkova, B.; Hajslova, J. Effect of high-pressure thermal sterilization (HPTS) on the reduction of food processing contaminants (e.g., furan, acrylamide, 3-MCPD-esters, HMF). In Present and Future of High Pressure Processing; Elsevier: Amsterdam, The Netherlands, 2020; pp. 139–172. [Google Scholar] [CrossRef]
- Alejandro Aldrete-Tapia, J.; Antonio Torres, J. Enhancing the Inactivation of Bacterial Spores during Pressure-Assisted Thermal Processing. Food Eng. Rev. 2021, 13, 431–441. [Google Scholar] [CrossRef]
- Lenz, C.; Vogel, R. Pressure-based strategy for the inactivation of spores. Sub-Cell. Biochem. 2015, 72, 469–537. [Google Scholar] [CrossRef]
- Sevenich, R.; Bark, F.; Crews, C.; Anderson, W.; Pye, C.; Riddellova, K.; Hradecky, J.; Moravcova, E.; Reineke, K.; Knorr, D. Effect of high pressure thermal sterilization on the formation of food processing contaminants. Innov. Food Sci. Emerg. Technol. 2013, 20, 42–50. [Google Scholar] [CrossRef]
- Reineke, K.; Doehner, I.; Schlumbach, K.; Baier, D.; Mathys, A.; Knorr, D. The different pathways of spore germination and inactivation in dependence of pressure and temperature. Innov. Food Sci. Emerg. Technol. 2012, 13, 31–41. [Google Scholar] [CrossRef]
- Roberts, C.; Hoover, D. Sensitivity of Bacillus coagulans Spores to Combinations of High Hydrostatic Pressure, Heat, Acidity and Nisin. J. Appl. Bacteriol. 1996, 81, 363–368. [Google Scholar] [CrossRef]
- López-Pedemonte, T.; Roig-Sagués, A.; Trujillo, A.; Capellas, M.; Guamis, B. Inactivation of spores of Bacillus cereus in cheese by high hydrostatic pressure with the addition of nisin or lysozyme. J. Dairy Sci. 2003, 86, 3075–3081. [Google Scholar] [CrossRef] [Green Version]
- Hofstetter, S.; Gebhardt, D.; Ho, L.; Gänzle, M.; McMullen, L.M. Effects of nisin and reutericyclin on resistance of endospores of Clostridium spp. to heat and high pressure. Food Microbiol. 2013, 34, 46–51. [Google Scholar] [CrossRef]
- Aouadhi, C.; Simonin, H.; Mejri, S.; Maaroufi, A. The combined effect of nisin, moderate heating and high hydrostatic pressure on the inactivation of Bacillus sporothermodurans spores. J. Appl. Microbiol. 2013, 115, 147–155. [Google Scholar] [CrossRef]
- Black, E.; Linton, M.; McCall, R.; Curran, W.; Fitzgerald, F.; Kelly, A.; Patterson, M. The combined effects of high pressure and nisin on germination and inactivation of Bacillus spores in milk. J. Appl. Microbiol. 2008, 105, 78–87. [Google Scholar] [CrossRef]
- Palman, Y.; Leo, R.; Pulvirenti, A.; Green, S.J.; Hayouka, Z. Antimicrobial peptide cocktail activity in minced turkey meat. Food Microbiol. 2020, 92, 103580. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R. Cationic peptides: Effectors in innate immunity and novel antimicrobials. Lancet Infect. Dis. 2001, 1, 156–164. [Google Scholar] [CrossRef]
- Axelsen, P. A chaotic pore model of polypeptide antibiotic action. Biophys. J. 2008, 94, 1549–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathinakumar, R.; Walkenhorst, W.; Wimley, W. Broad-spectrum antimicrobial peptides by rational combinatorial design and high-throughput screening: The importance of interfacial activity. J. Am. Chem. Soc. 2009, 131, 7609–7617. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuji, T.; Gallo, R. Antimicrobial peptides: Old molecules with new ideas. J. Investig. Dermatol. 2012, 132, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.; Daroit, D.; Brandelli, A. Food applications of liposome-encapsulated antimicrobial peptides. Trends Food Sci. Technol. 2010, 21, 284–292. [Google Scholar] [CrossRef]
- Liu, Y.; Eichler, J.; Pischetsrieder, M. Virtual screening of a milk peptide database for the identification of food-derived antimicrobial peptides. Mol. Nutr. Food Res. 2015, 59, 2243–2254. [Google Scholar] [CrossRef]
- Cleveland, J.; Montville, T.J.; Nes, I.; Chikindas, M. Bacteriocins: Safe, Natural Antimicrobials for Food Preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef]
- Hayouka, Z.; Chakraborty, S.; Liu, R.; Boersma, M.; Weisblum, B.; Gellman, S.H. Interplay among subunit identity, subunit proportion, chain length, and stereochemistry in the activity profile of sequence-random peptide mixtures. J. Am. Chem. Soc. 2013, 135, 11748–11751. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Role of Ger Proteins in Nutrient and Nonnutrient Triggering of Spore Germination in Bacillus subtilis. J. Bacteriol. 2000, 182, 2513–2519. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Observations on research with spores of Bacillales and Clostridiales species. J. Appl. Microbiol. 2019, 126, 348–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, D.; Wang, X.; Wu, X.; Liao, X.; Chen, F.; Hu, X. The effect of high pressure combined with moderate temperature and peptidoglycan fragments on spore inactivation. Food Res. Int. 2021, 148, 110615. [Google Scholar] [CrossRef] [PubMed]
- Black, E.; Koziol-Dube, K.; Guan, D.; Wei, J.; Setlow, B.; Cortezzo, D.; Hoover, D.; Setlow, P. Factors influencing germination of Bacillus subtilis spores via activation of nutrient receptors by high pressure. Appl. Environ. Microbiol. 2005, 71, 5879–5887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagler, K.; Setlow, P.; Li, Y.; Moeller, R. High salinity alters the germination behavior of Bacillus subtilis spores with nutrient and nonnutrient germinants. Appl. Environ. Microbiol. 2014, 80, 1314–1321. [Google Scholar] [CrossRef] [Green Version]
- Mathys, A.; Chapman, B.; Bull, M.; Heinz, V.; Knorr, D. Flow cytometric assessment of Bacillus spore response to high pressure and heat. Innov. Food Sci. Emerg. Technol. 2007, 8, 519–527. [Google Scholar] [CrossRef]
- Zhang, Y.; Delbrück, A.I.; Off, C.; Benke, S.; Mathys, A. Flow Cytometry Combined With Single Cell Sorting to Study Heterogeneous Germination of Bacillus Spores Under High Pressure. Front. Microbiol. 2020, 10, 3118. [Google Scholar] [CrossRef] [Green Version]
- Abhyankar, W.; Wen, J.; Swarge, B.; Tu, Z.; Boer, R.; Smelt, J.; Koning, L.; Manders, E.; Koster, C.; Brul, S. Proteomics and microscopy tools for the study of antimicrobial resistance and germination mechanisms of bacterial spores. Food Microbiol. 2019, 81, 89–96. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.; Blake, J.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spore Resistance Properties. Microbiol. Spectr. 2014, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Henriques, A.; Moran, C. Structure, assembly, and function of the spore surface layers. Annu. Rev. Microbiol. 2007, 61, 555–588. [Google Scholar] [CrossRef]
- Klobutcher, L.; Ragkousi, K.; Setlow, P. The Bacillus subtilis Spore Coat Provides “eat Resistance” during Phagocytic Predation by the Protozoan Tetrahymena Thermophila. Proc. Natl. Acad. Sci. USA 2005, 103, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setlow, P. Summer meeting 2013—When the sleepers wake: The germination of spores of Bacillus species. J. Appl. Microbiol. 2013, 115, 1251–1268. [Google Scholar] [CrossRef]
- Griffiths, K.; Zhang, J.; Cowan, A.; Yu, J.; Setlow, P. Germination proteins in the inner membrane of dormant Bacillus subtilis spores colocalize in a discrete cluster. Mol. Microbiol. 2011, 81, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Loison, P.; Hosny, N.; Gervais, P.; Champion, D.; Kuimova, M.; Perrier-Cornet, J. Direct investigation of viscosity of an atypical inner membrane of Bacillus spores: A molecular rotor/FLIM study. Biochim. Biophys. Acta—Biomembr. 2013, 1828, 2436–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setlow, P. I will survive: DNA protection in bacterial spores. Trends Microbiol. 2007, 15, 172–180. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | % Gated | |||
|---|---|---|---|---|
| UL | UR | LL | LR | |
| Untreated | 12.24 ± 0.54 f | 17.69 ± 1.11 e | 61.83 ± 1.93 c | 8.23 ± 1.1 fg |
| 200 MPa 15 min | 0.06 ± 0.04 i | 76.89 ± 1.42 b | 2.55 ± 0.47 hi | 20.51 ± 1.48 e |
| 80 °C 15 min | 0.41 ± 0.31 i | 50.06 ± 2.56 d | 3.02 ± 1.65 hi | 46.52 ± 3.96 d |
| 102 μg/mL LK 25 °C 15 min | 0.06 ± 0.06 i | 46.59 ± 3.68 d | 3.82 ± 0.46 ghi | 49.53 ± 3.57 d |
| 200 MPa 80 °C 102 μg/mL LK 15 min | 0.35 ± 0.11 i | 82.18 ± 1.44 a | 6.28 ± 1.07 gh | 11.2 ± 2.28 f |
| Protein Description | Test Sequences | p-Value | Rich Factor |
|---|---|---|---|
| Citrate cycle (TCA cycle) | |||
| 2-Oxoglutarate dehydrogenase E1 component | P23129 | 0.00 | 0.18 |
| Aconitate/2-methylaconitate hydratase | P09339 | 0.00 | 0.18 |
| Malate dehydrogenase | P49814 | 0.00 | 0.18 |
| Pyruvate carboxylase | Q9KWU4 | 0.00 | 0.18 |
| Dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex | A0A6A8FKK9 | 0.00 | 0.18 |
| Phosphoenolpyruvate carboxykinase (ATP) | A0A7U5BTG7 | 0.00 | 0.18 |
| Succinate–CoA ligase [ADP-forming] subunit alpha | A0A5F2KMI5 | 0.00 | 0.18 |
| Succinate dehydrogenase flavoprotein subunit | A0A3N6CX89 | 0.00 | 0.18 |
| Succinate–CoA ligase [ADP-forming] subunit beta | A0A3A5I5U1 | 0.00 | 0.18 |
| Ribosome | |||
| 30S ribosomal protein S13 | P20282 | 0.03 | 0.13 |
| 50S ribosomal protein L5 | P12877 | 0.03 | 0.13 |
| 30S ribosomal protein S1 homolog | P38494 | 0.03 | 0.13 |
| 30S ribosomal protein S2 | P21464 | 0.03 | 0.13 |
| 50S ribosomal protein L13 | M4KMS5 | 0.03 | 0.13 |
| 50S ribosomal protein L29 | D4G3L1 | 0.03 | 0.13 |
| 30S ribosomal protein S4 | A0A4R6HVR7 | 0.03 | 0.13 |
| 30S ribosomal protein S14 | A0A5D4N259 | 0.03 | 0.13 |
| Oxidative phosphorylation | |||
| Quinol oxidase subunit 1 | P34956 | 0.02 | 0.15 |
| ATP synthase subunit c | P37815 | 0.02 | 0.15 |
| ATP synthase subunit alpha | A0A0D1KS60 | 0.02 | 0.15 |
| Cytochrome c oxidase subunit IVB | A0A5F2KHE8 | 0.02 | 0.15 |
| Succinate dehydrogenase flavoprotein subunit | A0A3N6CX89 | 0.02 | 0.15 |
| Cytochrome c oxidase subunit 2 | A0A0G2YRU4 | 0.02 | 0.15 |
| Glycine, serine, and threonine metabolism | |||
| Betaine aldehyde dehydrogenase | P71016 | 0.04 | 0.14 |
| Aminomethyltransferase | P54378 | 0.04 | 0.14 |
| l-Serine ammonia-lyase, iron-sulfur-dependent subunit beta | A0A8A7MTS3 | 0.04 | 0.14 |
| Hydroxymethyltransferase | A0A857HNR6 | 0.04 | 0.14 |
| Glycine dehydrogenase (Decarboxylating) subunit 1 | A0A857HK36 | 0.04 | 0.14 |
| NAD-dependent alcohol dehydrogenase | A0A8B5NR48 | 0.04 | 0.14 |
| Carbon fixation pathways in prokaryotes | |||
| Aconitate/2-methylaconitate hydratase | P09339 | 0.04 | 0.14 |
| Malate dehydrogenase | P49814 | 0.04 | 0.14 |
| Pyruvate carboxylase | Q9KWU4 | 0.04 | 0.14 |
| Succinate–CoA ligase (ADP-forming) subunit alpha | A0A5F2KMI5 | 0.04 | 0.14 |
| Succinate dehydrogenase flavoprotein subunit | A0A3N6CX89 | 0.04 | 0.14 |
| Succinate–CoA ligase (ADP-forming) subunit beta | A0A3A5I5U1 | 0.04 | 0.14 |
| Protein Description | Test Sequences | p-Value | Rich Factor |
|---|---|---|---|
| Ribosome | |||
| 30S ribosomal protein S13 | P20282 | 0.00 | 0.078 |
| 50S ribosomal protein L21 | A0A1A0G613 | 0.00 | 0.078 |
| 30S ribosomal protein S2 | P21464 | 0.00 | 0.078 |
| 30S ribosomal protein S7 | A0A7Z9E609 | 0.00 | 0.078 |
| 30S ribosomal protein S4 | A0A4R6HVR7 | 0.00 | 0.078 |
| Protein Description | Test Sequences | p-Value | Rich Factor |
|---|---|---|---|
| Ribosome | |||
| 30S ribosomal protein S13 | P20282 | 0.00 | 0.09 |
| 50S ribosomal protein L2 | P42919 | 0.00 | 0.09 |
| 30S ribosomal protein S2 | P21464 | 0.00 | 0.09 |
| 30S ribosomal protein S7 | A0A7Z9E609 | 0.00 | 0.09 |
| 30S ribosomal protein S4 | A0A4R6HVR7 | 0.00 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, Y.; Wu, R.; Cui, T.; Zhang, Z.; Dong, L.; Chen, F.; Hu, X. Proteomic Response of Bacillus subtilis Spores under High Pressure Combined with Moderate Temperature and Random Peptide Mixture LK Treatment. Foods 2022, 11, 1123. https://doi.org/10.3390/foods11081123
Pang Y, Wu R, Cui T, Zhang Z, Dong L, Chen F, Hu X. Proteomic Response of Bacillus subtilis Spores under High Pressure Combined with Moderate Temperature and Random Peptide Mixture LK Treatment. Foods. 2022; 11(8):1123. https://doi.org/10.3390/foods11081123
Chicago/Turabian StylePang, Yaru, Ruobin Wu, Tianlin Cui, Zequn Zhang, Li Dong, Fang Chen, and Xiaosong Hu. 2022. "Proteomic Response of Bacillus subtilis Spores under High Pressure Combined with Moderate Temperature and Random Peptide Mixture LK Treatment" Foods 11, no. 8: 1123. https://doi.org/10.3390/foods11081123
APA StylePang, Y., Wu, R., Cui, T., Zhang, Z., Dong, L., Chen, F., & Hu, X. (2022). Proteomic Response of Bacillus subtilis Spores under High Pressure Combined with Moderate Temperature and Random Peptide Mixture LK Treatment. Foods, 11(8), 1123. https://doi.org/10.3390/foods11081123