

Potential Use of Cardunculus Biomass on Pleurotus eryngii Production: Heteroglycans Content and Nutritional Properties (Preliminary Results)

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
- (i)
- To evaluate the use of cardunculus biomass on P. eryngii fruiting bodies yield.
- (ii)
- To evaluate the possibility to use P. eryngii mycelium, cultivated on media rich in inulin from cardunculus roots, as a source to produce functional heteroglycans.
- (iii)
- To evaluate the influence of cardunculus biomass on heteroglycan content in fruit bodies of P. eryngii.
- (iv)
- To assess the nutraceutical parameters of fruiting bodies grown on cardunculus biomass.
2. Materials and Methods
2.1. Fungal Isolates
2.2. Spawn-Production
2.3. Assessment of the Ability of P. eryngii to Grow on Cardunculus Roots and Stems
2.4. Mycelial Growth
2.5. Evaluation of Fruiting Bodies Production Caractheristics on Different Subtrates
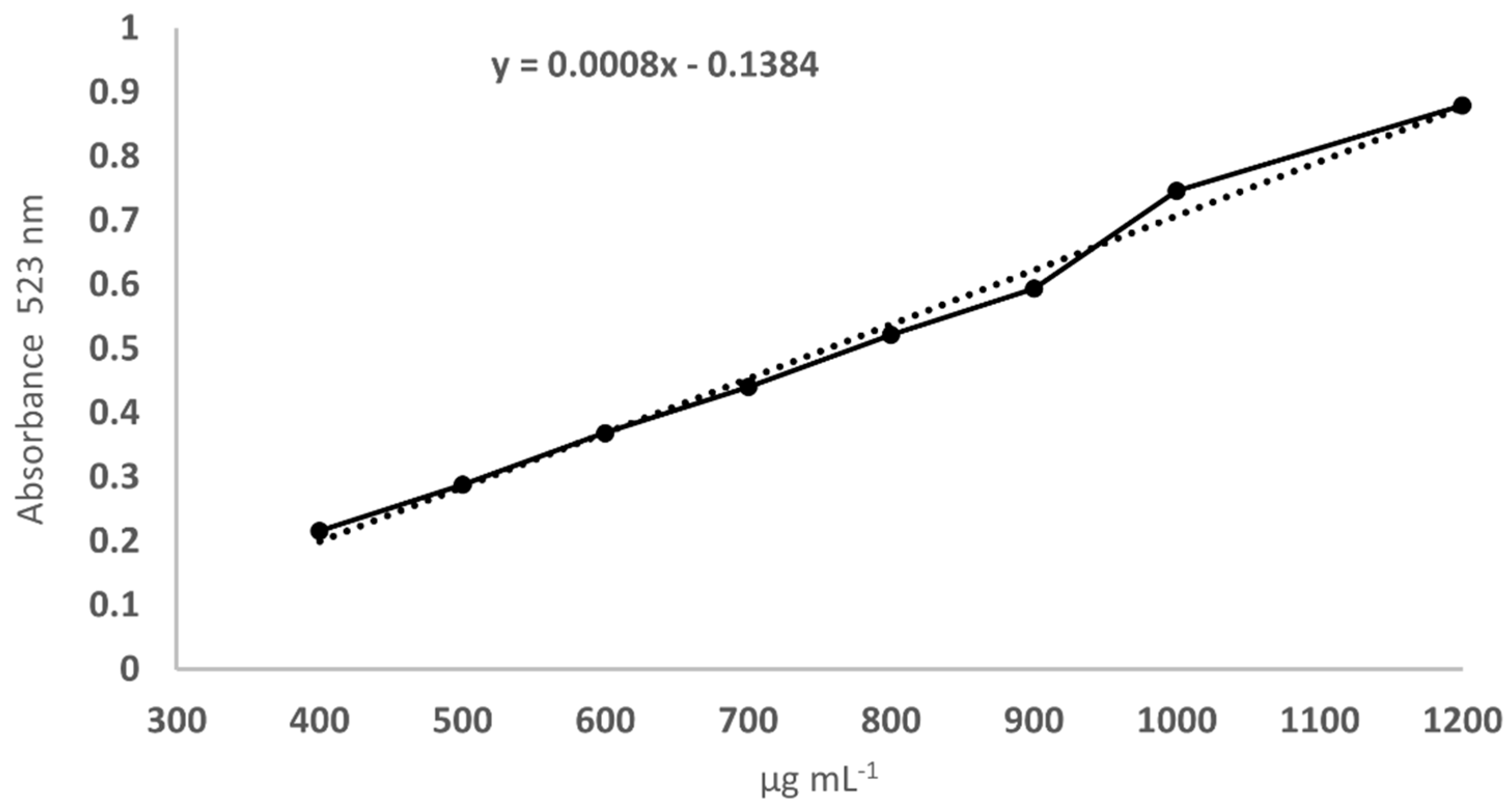
2.6. Heteroglycans Determination in Mycelium and Fruiting Bodies
2.7. Bioactive Compounds, Antioxidant Activity, and Enzymatic Assays
2.8. Statistical Analysis
3. Results
3.1. Assessment of P. eryngii Isolate’s Ability to Colonize Cardunculus Roots and Stems
3.2. Fungal Isolate
3.3. Effects on Mycelial Production by the Addition of Inulin and Sucrose In Vitro
3.4. Fruiting Bodies Yield and Heteroglycans Content on Different Substrates
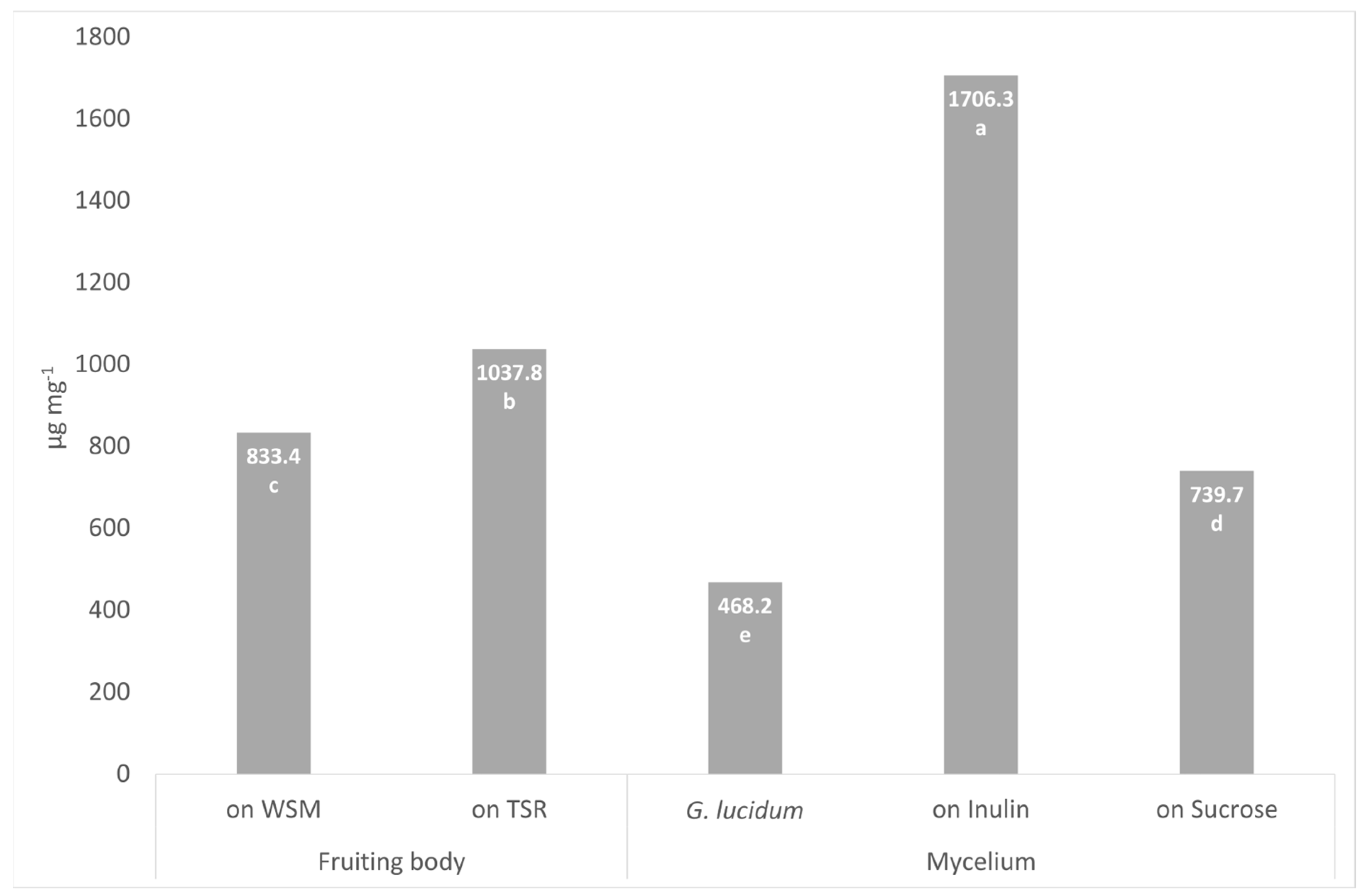
3.5. Heteroglycans Content in Fruiting Bodies and Mycelium of P. eryngii
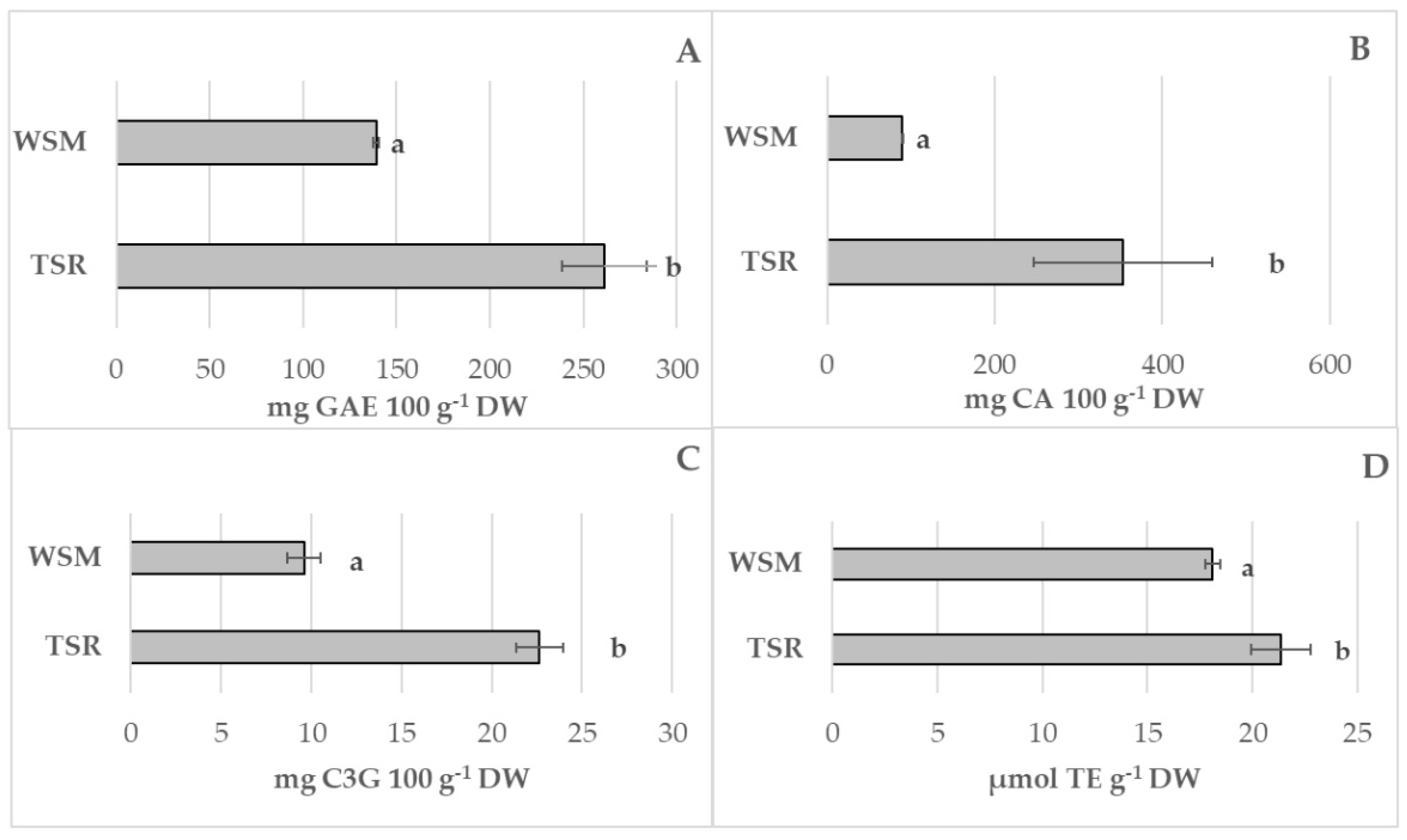
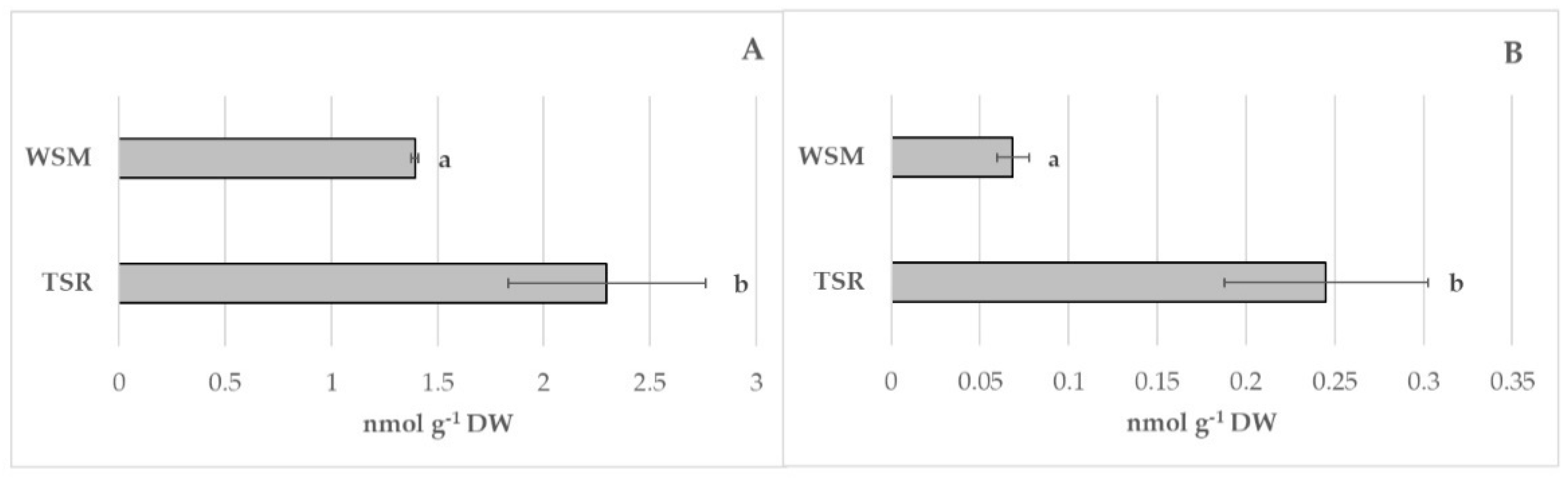
3.6. Evaluation of Different Bioactive Compounds and Antioxidant Activity and Enzymes Involved in Browning Reaction and Membrane Damage
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Takahashi, J.A.; Barbosa, B.V.R.; Martins, B.d.A.; Guirlanda, C.P.; Moura, M.A.F. Use of the Versatility of Fungal Metabolism to Meet Modern Demands for Healthy Aging, Functional Foods, and Sustainability. J. Fungi 2020, 6, 223. [Google Scholar] [CrossRef] [PubMed]
- Gominho, J.; Curt, M.D.; Lourenco, A.; Fernández, J.; Pereira, H. Cynara cardunculus L. as a biomass and multi-purpose crop: A review of 30 years of research. Biomass Bioenergy 2018, 109, 257–275. [Google Scholar] [CrossRef]
- Ciancolini, A.; Alignan, M.; Pagnotta, M.A.; Vilarem, G.; Crinò, P. Selection of Italian cardoon genotypes as industrial crop for biomass and polyphenol production. Ind. Crops Prod. 2013, 51, 145–151. [Google Scholar] [CrossRef]
- Carrasco, J.; Zied, D.C.; Pardo, J.E.; Preston, G.M.; Pardo-Giménez, A. Supplementation in mushroom crops and its impact on yield and quality. AMB Express 2018, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Pari, L.; Alfano, V.; Stefanoni, W.; Latterini, F.; Liuzzi, F.; De Bari, I.; Valerio, V.; Ciancolini, A. Inulin Content in Chipped and Whole Roots of Cardoon after Six Months Storage under Natural Conditions. Sustainability 2021, 13, 3902. [Google Scholar] [CrossRef]
- Melilli, M.; Branca, F.; Sillitti, C.; Scandurra, S.; Calderaro, P.; Di Stefano, V. Germplasm evaluation to obtain inulin with high degree of polymerization in Mediterranean environment. Nat. Prod. Res. 2020, 34, 187–191. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, Y.; Xu, Z.; Yang, W.; Mariga, A.M.; Pang, G.; Geng, C.; Hu, Q. Immunoregulatory role of Pleurotus eryngii superfine powder through intercellular communication of cytokines. Food Agric. Immunol. 2014, 25, 586–599. [Google Scholar] [CrossRef] [Green Version]
- Mubasshira, M.; Aminuzzaman, F.M.; Sultana, N.; Tanni, F. Impact of Different Substrates and Mother Cultures on Yield and Yield Attributes of Oyster Mushroom (Pleurotus ostreatus). Asian Food Sci. J. 2020, 19, 25–38. [Google Scholar] [CrossRef]
- Kirbag, S.; Akyuz, M. Effect of various agro-residues on growing periods, yield and biological efficiency of Pleurotus eryngii. J. Food Agric. Environ. 2008, 6, 402–405. [Google Scholar]
- Panjikkaran, S.T.; Mathew, D. An environmentally friendly and cost effective technique for the commercial cultivation of oyster mushroom [Pleurotus florida (Mont.) Singer]. J. Sci. Food Agric. 2013, 93, 973–976. [Google Scholar] [CrossRef]
- Ajith, T.A.; Janardhanan, K.K. Indian medicinal mushrooms as a source of antioxidant and antitumor agents. J. Clin. Biochem. Nutr. 2007, 40, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Mariga, A.M.; Pei, F.; Yang, W.-J.; Zhao, L.-Y.; Shao, Y.-N.; Mugambi, D.K.; Hu, Q.-H. Immunopotentiation of Pleurotus eryngii (DC. ex Fr.) Quel. J. Ethnopharmacol. 2014, 153, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, H.W. Innate immunity induced by fungal β-glucans via dectin-1 signaling pathway. Int. J. Med. Mushrooms 2014, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, R.; Manco, R.; Sapio, D.; Iannaccone, M.; Fulgione, A.; Papaianni, M.; de Falco, B.; Grauso, L.; Tarantino, P.; Ianniello, F. Structural data and immunomodulatory properties of a water-soluble heteroglycan extracted from the mycelium of an Italian isolate of Ganoderma lucidum. Nat. Prod. Res. 2017, 31, 2119–2125. [Google Scholar] [CrossRef]
- Kim, Y.H.; Jung, E.-G.; Han, K.-I.; Patnaik, B.B.; Kwon, H.-J.; Lee, H.-S.; Kim, W.J.; Han, M.-D. Immunomodulatory Effects of Extracellular β-Glucan Isolated from the King Oyster Mushroom Pleurotus eryngii (Agaricomycetes) and Its Sulfated Form on Signaling Molecules Involved in Innate Immunity. Int. J. Med. Mushrooms 2017, 19, 521–533. [Google Scholar] [CrossRef]
- Vetvicka, V.; Gover, O.; Karpovsky, M.; Hayby, H.; Danay, O.; Ezov, N.; Hadar, Y.; Schwartz, B. Immune-modulating activities of glucans extracted from Pleurotus ostreatus and Pleurotus eryngii. J. Funct. Foods 2019, 54, 81–91. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Y.; Zhang, F.; Linhardt, R.J.; Zeng, G.; Zhang, A. Extraction, structure and bioactivities of the polysaccharides from Pleurotus eryngii: A review. Int. J. Biol. Macromol. 2020, 150, 1342–1347. [Google Scholar] [CrossRef]
- Murphy, E.J.; Masterson, C.; Rezoagli, E.; O’Toole, D.; Major, I.; Stack, G.D.; Lynch, M.; Laffey, J.G.; Rowan, N.J. β-Glucan extracts from the same edible shiitake mushroom Lentinus edodes produce differential in-vitro immunomodulatory and pulmonary cytoprotective effects-Implications for coronavirus disease (COVID-19) immunotherapies. Sci. Total Environ. 2020, 732, 139330. [Google Scholar] [CrossRef]
- Cerletti, C.; Esposito, S.; Iacoviello, L. Edible mushrooms and beta-glucans: Impact on human health. Nutrients 2021, 13, 2195. [Google Scholar] [CrossRef]
- Avni, S.; Ezove, N.; Hanani, H.; Yadid, I.; Karpovsky, M.; Hayby, H.; Gover, O.; Hadar, Y.; Schwartz, B.; Danay, O. Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii. Int. J. Mol. Sci. 2017, 18, 1564. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-T.; Miles, P.G. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Zayed, A.; Serag, A.; Farag, M.A. Cynara cardunculus L.: Outgoing and potential trends of phytochemical, industrial, nutritive and medicinal merits. J. Funct. Foods 2020, 69, 103937. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Ahmed, M.; Abdullah, N.; Ahmed, K.U.; Bhuyan, M.H.M. Yield and nutritional composition of oyster mushroom strains newly introduced in Bangladesh. Pesqui. Agropecuária Bras. 2013, 48, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Nitschke, J.; Modick, H.; Busch, E.; von Rekowski, R.W.; Altenbach, H.-J.; Mölleken, H. A new colorimetric method to quantify β-1,3-1,6-glucans in comparison with total β-1,3-glucans in edible mushrooms. Food Chem. 2011, 127, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Familoni, T.V.; Ogidi, C.O.; Akinyele, B.J.; Onifade, A.K. Evaluation of yield, biological efficiency and proximate composition of Pleurotus species cultivated on different wood dusts. Czech Mycol. 2018, 70, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Kaur, C.; Walia, S.; Nagal, S.; Walia, S.; Singh, J.; Singh, B.B.; Saha, S.; Singh, B.; Kalia, P.; Jaggi, S.; et al. Functional quality and antioxidant composition of selected tomato (Solanum lycopersicon L.) cultivars grown in Northern India. LWT-Food Sci. Technol. 2013, 50, 139–145. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. Curr. Protoc. Food Anal. Chem. 2002, 6, I1.1.1–I1.1.8. [Google Scholar]
- Goffi, V.; Zampella, L.; Forniti, R.; Petriccione, M.; Botondi, R. Effects of ozone postharvest treatment on physicochemical and qualitative traits of Actinidia chinensis ‘Soreli’during cold storage. J. Sci. Food Agric. 2019, 99, 5654–5661. [Google Scholar] [CrossRef]
- Adiletta, G.; Magri, A.; Albanese, D.; Liguori, L.; Sodo, M.; Di Matteo, M.; Petriccione, M. Overall quality and oxidative damage in packaged freshly shelled walnut kernels during cold storage. J. Food Meas. Charact. 2020, 14, 3483–3492. [Google Scholar] [CrossRef]
- Magri, A.; Adiletta, G.; Petriccione, M. Evaluation of Antioxidant Systems and Ascorbate-Glutathione Cycle in Feijoa Edible Flowers at Different Flowering Stages. Foods 2020, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Caracciolo, G.; Magri, A.; Petriccione, M.; Maltoni, M.L.; Baruzzi, G. Influence of cold storage on pear physico-chemical traits and antioxidant systems in relation to superficial scald development. Foods 2020, 9, 1175. [Google Scholar] [CrossRef] [PubMed]
- Adiletta, G.; Di Matteo, M.; Albanese, D.; Farina, V.; Cinquanta, L.; Corona, O.; Magri, A.; Petriccione, M. Changes in physico-chemical traits and enzymes oxidative system during cold storage of ‘Formosa’papaya fresh cut fruits grown in the mediterranean area (Sicily). Ital. J. Food Sci. 2020, 32. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- De Gioia, T.; Sisto, D.; Rana, G.L.; Figliuolo, G. Genetic structure of the Pleurotus eryngii species-complex. Mycol. Res. 2005, 109, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Lan, J. Advances in the research of Pleurotus eryngi. Shi Yong Jun Xue Bao 2004, 11, 52–58. [Google Scholar]
- Fernández, J.; Curt, M.D.; Aguado, P.L. Industrial applications of Cynara cardunculus L. for energy and other uses. Ind. Crops Prod. 2006, 24, 222–229. [Google Scholar] [CrossRef]
- Aida, F.M.N.A.; Shuhaimi, M.; Yazid, M.; Maaruf, A.G. Mushroom as a potential source of prebiotics: A review. Trends Food Sci. Technol. 2009, 20, 567–575. [Google Scholar] [CrossRef]
- Pessoni, R.A.; Freshour, G.; Figueiredo-Ribeiro, R.d.C.L.; Hahn, M.G.; Braga, M.R. Cell-wall structure and composition of Penicillium janczewskii as affected by inulin. Mycologia 2005, 97, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Boh, B.; Berovic, M.; Zhang, J.; Zhi-Bin, L. Ganoderma lucidum and its pharmaceutically active compounds. In Biotechnology Annual Review; El-Gewely, M.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 13, pp. 265–301. [Google Scholar]
- Raccuia, S.A.; Melilli, M. Cynara cardunculus L., a potential source of inulin in the Mediterranean environment: Screening of genetic variability. Aust. J. Agric. Res. 2004, 55, 693–698. [Google Scholar] [CrossRef]
- Tarko, D.B.; Sirna, A.M. Substrate optimization for cultivation of Pleurotus ostreatus on lignocellulosic wastes (coffee, sawdust, and sugarcane bagasse) in Mizan–Tepi University, Tepi Campus, Tepi Town. J. Appl. Biol. Biotechnol. 2018, 6, 14–20. [Google Scholar]
- Bonatti, M.; Karnopp, P.; Soares, H.M.; Furlan, S.A. Evaluation of Pleurotus ostreatus and Pleurotus sajor-caju nutritional characteristics when cultivated in different lignocellulosic wastes. Food Chem. 2004, 88, 425–428. [Google Scholar] [CrossRef]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Sardar, H.; Ali, M.A.; Anjum, M.A.; Nawaz, F.; Hussain, S.; Naz, S.; Karimi, S.M. Agro-industrial residues influence mineral elements accumulation and nutritional composition of king oyster mushroom (Pleurotus eryngii). Sci. Hortic. 2017, 225, 327–334. [Google Scholar] [CrossRef]
- da Paz, M.F.; Breyer, C.A.; Longhi, R.F.; Oviedo, M.S.V.P. Determining the basic composition and total phenolic compounds of Pleurotus sajor-caju cultivated in three different substrates by solid state bioprocess. J. Biotechnol. Biodivers. 2012, 3, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Amić, D.; Davidović-Amić, D.; Bešlo, D.; Trinajstić, N. Structure-radical scavenging activity relationships of flavonoids. Croat. Chem. Acta 2003, 76, 55–61. [Google Scholar]
- Islam, T.; Yu, X.; Xu, B. Phenolic profiles, antioxidant capacities and metal chelating ability of edible mushrooms commonly consumed in China. LWT-Food Sci. Technol. 2016, 72, 423–431. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2016, 242, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Ng, Z.X.; Than, M.J.Y.; Yong, P.H. Peperomia pellucida (L.) Kunth herbal tea: Effect of fermentation and drying methods on the consumer acceptance, antioxidant and anti-inflammatory activities. Food Chem. 2021, 344, 128738. [Google Scholar] [CrossRef]
- Mishra, K.K.; Pal, R.S.; Arunkumar, R.; Chandrashekara, C.; Jain, S.K.; Bhatt, J.C. Antioxidant properties of different edible mushroom species and increased bioconversion efficiency of Pleurotus eryngii using locally available casing materials. Food Chem. 2013, 138, 1557–1563. [Google Scholar] [CrossRef]
- Belozerskaya, T.A.; Gessler, N.N.; Aver’yanov, A.A. Melanin Pigments of Fungi; Springer: Berlin, Germany, 2016; pp. 1–29. [Google Scholar] [CrossRef]
- Li, D.; Wang, D.; Fang, Y.; Belwal, T.; Li, L.; Lin, X.; Xu, Y.; Chen, H.; Zhu, M.; Luo, Z. Involvement of energy metabolism and amino acid metabolism in quality attributes of postharvest Pleurotus eryngii treated with a novel phase change material. Postharvest Biol. Technol. 2021, 173, 111427. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Chang, C.-K.; Wong, L.-W.; Hu, C.-C.; Lin, J.-A.; Hsieh, C.-W. Alternating current electric field inhibits browning of Pleurotus ostreatus via inactivation of oxidative enzymes during postharvest storage. LWT 2020, 134, 110212. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Media | Sucrose (g L−1) | Inulin (g L−1) |
|---|---|---|
| 1 | 30 | 0 |
| 2 | 27 | 3 |
| 3 | 21 | 9 |
| 4 | 15 | 15 |
| 5 | 9 | 21 |
| 6 | 3 | 27 |
| 7 | 0 | 30 |
| Substrates 1 | Isolates | Mean | ||||
|---|---|---|---|---|---|---|
| Pe1Ce | Pe2CE | Pe3Ce | Pe4CE | Pe5Ce | ||
| WS | 0.6 | 0.4 | 0.5 | 0.4 | 0.4 | 0.48 |
| TR | 0.7 | 0.5 | 0.6 | 0.5 | 0.5 | 0.58 |
| TS | 0.8 | 0.5 | 0.7 | 0.5 | 0.6 | 0.62 |
| Mean | 0.68 | 0.48 | 0.62 | 0.49 | 0.52 | |
| Statistical analysis 2 | ||||||
| ANOVA | LSD 0.09 | |||||
| Isolate | ** | 0.062 | ||||
| Substrate | ** | 0.075 | ||||
| Isolate × Substrate | ** | 0.087 | ||||
| Media | Carbon Source | DW 1 after 12 Days of Growth (g) | ||
|---|---|---|---|---|
| Sucrose | Inulin | |||
| (g L−1) | (g L−1) | |||
| 1 | 30 | 0 | 10.1 ± 1.72 | b |
| 2 | 27 | 3 | 8.9 ± 1.12 | b |
| 3 | 21 | 9 | 9.1 ± 1.24 | b |
| 4 | 15 | 15 | 10.3 ± 1.45 | b |
| 5 | 9 | 21 | 9.2 ± 1.47 | b |
| 6 | 3 | 27 | 16.1 ± 1.87 | a |
| 7 | 0 | 30 | 15.9 ± 2.11 | a |
| Substrate 1 | MC 2 (days) | PA 3 (days) | FBF 4 (days) | MW 5 (g) | BE 6 (%) |
|---|---|---|---|---|---|
| TSI | 17.4 ± 0.82 b | 22.8 ± 1.12 c | 7.0 ± 0.6 a | 305.7 ± 12.5 b | 50.9 ± 5.7 bc |
| TSS | 15.6 ± 0.67 c | 26.5 ± 0.98 b | 7.0 ± 0.5 a | 310.5 ± 9.4 b | 51.7 ± 7.9 b |
| TSR | 15.6 ± 0.82 c | 26.0 ± 0.89 b | 5.0 ± 0.4 b | 371.7 ± 11.2 a | 61.9 ± 4.7 a |
| WSM | 19.2 ± 1.02 a | 29.3 ± 1.21 a | 8.0 ± 0.6 a | 280.6 ± 9.8 c | 46.7 ± 4.8 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Battaglia, V.; Sorrentino, R.; Verrilli, G.; del Piano, L.; Sorrentino, M.C.; Petriccione, M.; Sicignano, M.; Magri, A.; Cermola, M.; Cerrato, D.; et al. Potential Use of Cardunculus Biomass on Pleurotus eryngii Production: Heteroglycans Content and Nutritional Properties (Preliminary Results). Foods 2023, 12, 58. https://doi.org/10.3390/foods12010058
Battaglia V, Sorrentino R, Verrilli G, del Piano L, Sorrentino MC, Petriccione M, Sicignano M, Magri A, Cermola M, Cerrato D, et al. Potential Use of Cardunculus Biomass on Pleurotus eryngii Production: Heteroglycans Content and Nutritional Properties (Preliminary Results). Foods. 2023; 12(1):58. https://doi.org/10.3390/foods12010058
Chicago/Turabian StyleBattaglia, Valerio, Roberto Sorrentino, Giulia Verrilli, Luisa del Piano, Maria Cristina Sorrentino, Milena Petriccione, Mariarosaria Sicignano, Anna Magri, Michele Cermola, Domenico Cerrato, and et al. 2023. "Potential Use of Cardunculus Biomass on Pleurotus eryngii Production: Heteroglycans Content and Nutritional Properties (Preliminary Results)" Foods 12, no. 1: 58. https://doi.org/10.3390/foods12010058
APA StyleBattaglia, V., Sorrentino, R., Verrilli, G., del Piano, L., Sorrentino, M. C., Petriccione, M., Sicignano, M., Magri, A., Cermola, M., Cerrato, D., & Lahoz, E. (2023). Potential Use of Cardunculus Biomass on Pleurotus eryngii Production: Heteroglycans Content and Nutritional Properties (Preliminary Results). Foods, 12(1), 58. https://doi.org/10.3390/foods12010058