





Lactoferrin—The Health-Promoting Properties and Contemporary Application with Genetic Aspects



Abstract
:
1. Introduction
2. Materials and Methods
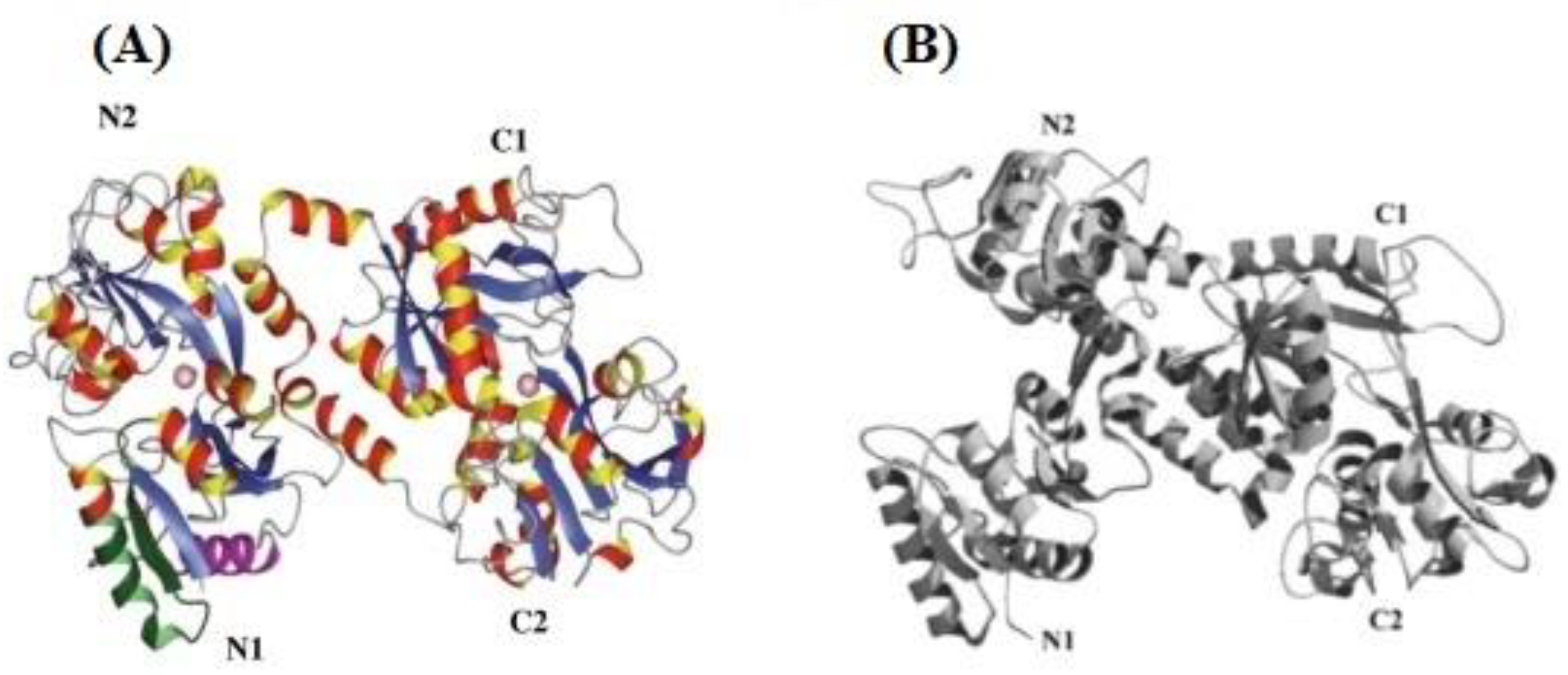
3. Occurrence and Structure of Lactoferrin
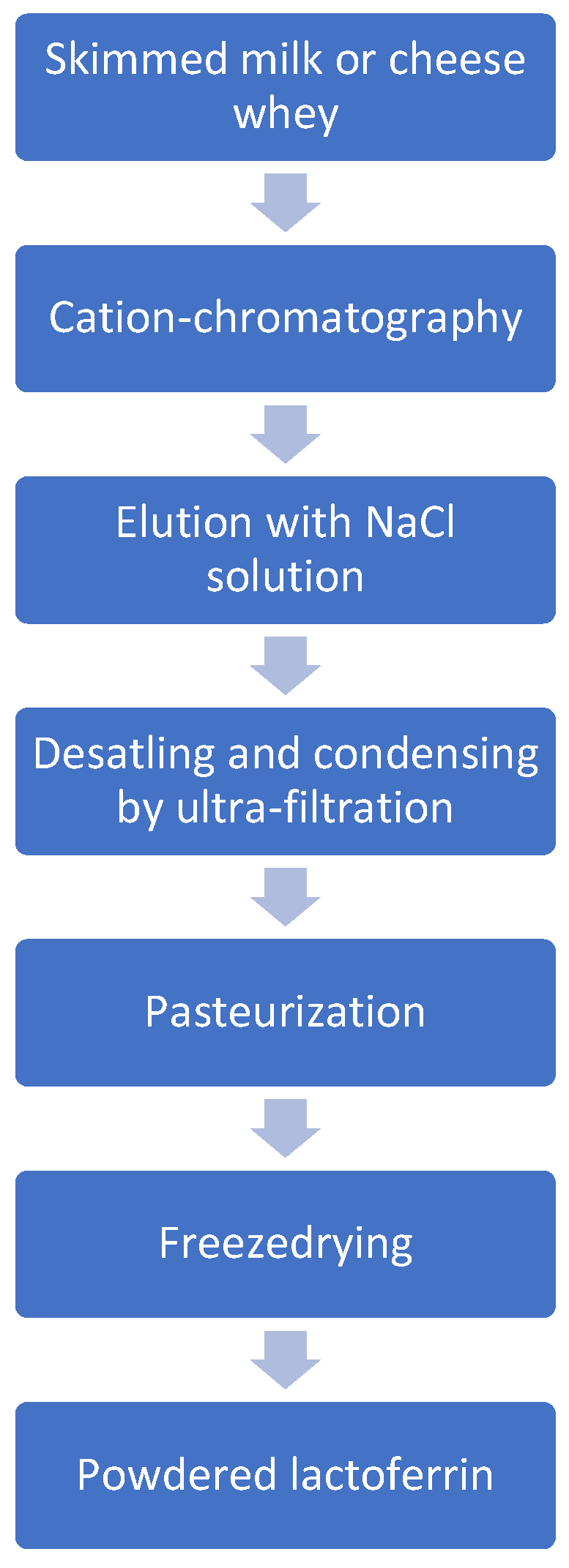
4. Acquisition of Lactoferrin
5. Effect of Temperature on Lactoferrin Content
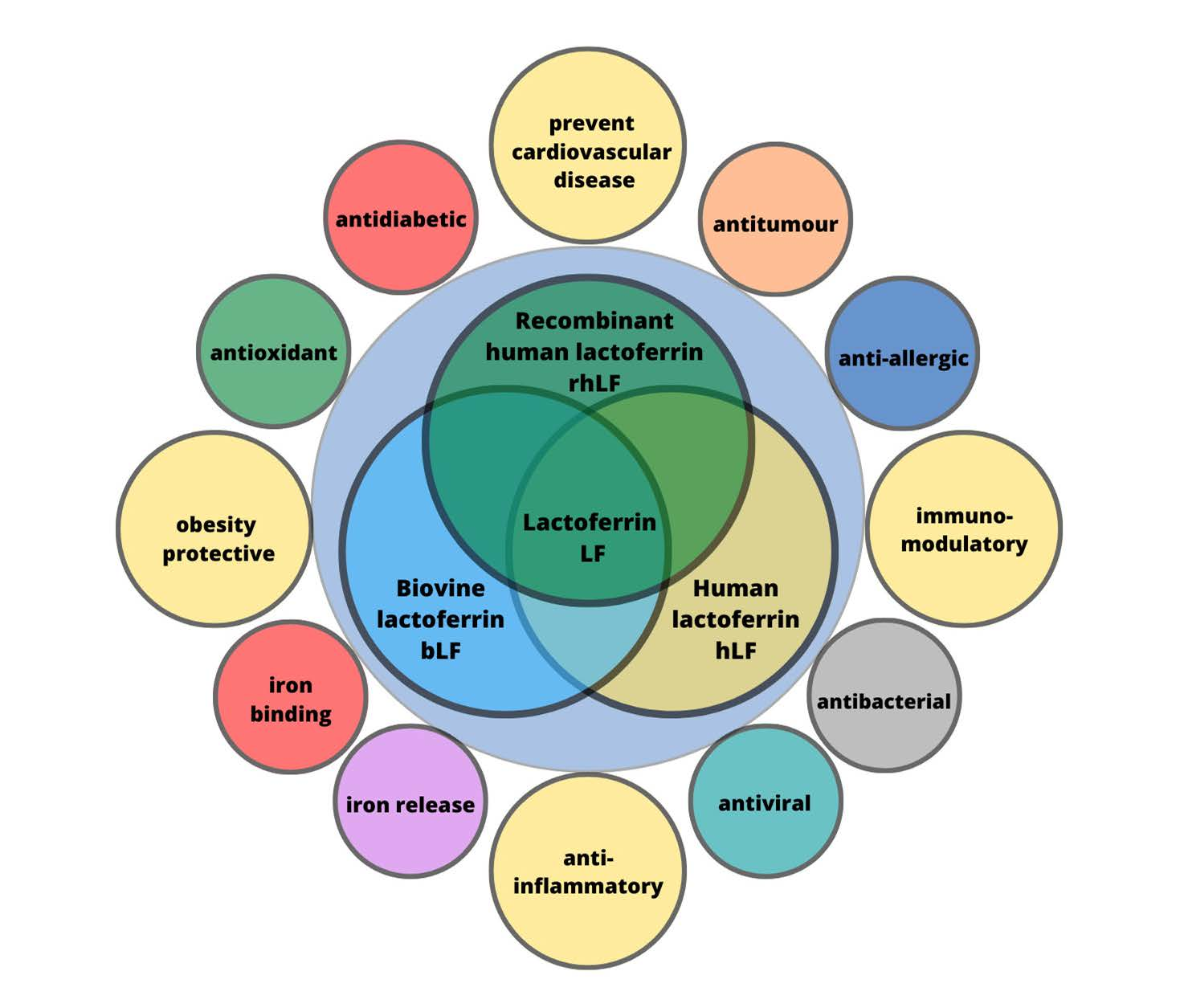
6. Biological Properties of Lactoferrin
6.1. LF and Obesity
6.2. LF and Type 2 Diabetes and Anti-Inflammatory Effects
6.3. LF and Cardiovascular Diseases
6.4. LF and Immune System
7. Scale of Lactoferrin Production
8. Applications in Industry
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Maciela, K.; Santos, L.S.; Ferreira Bonomo, R.C.; Alcântara Verissimoc, L.A.; Rodrigues Minim, V.P.; Minim, L.A. Purification of lactoferrin from sweet whey using ultrafiltration followed by expanded bed chromatography. Sep. Purif. Technol. 2020, 251, 117324. [Google Scholar] [CrossRef]
- Jamka, M. Historical overview on knowledge about lactoferrin. Acta Med. Pol. 2017, 7, 44–52. [Google Scholar] [CrossRef]
- Francoa, I.; Pérezb, M.D.; Conesab, C.; Calvob, M.; Sánchezb, L. Effect of technological treatments on bovine lactoferrin: An overview. Food Res. 2018, 106, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Wakabayashi, H.; Shin, K.; Yamauchi, K.; Yaeshima, T.; Iwatsuki, K. Twenty-five years of research on bovine lactoferrin applications. Biochimie 2009, 91, 52–57. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, N.; Kane, M.; Joshi, L.; Hickey, R.M. Structural and functional characteristics of bovine milk protein glycosylation. Glycobiology 2014, 24, 220–236. [Google Scholar] [CrossRef] [Green Version]
- Litwińczuk, Z.; Król, J.; Brodziak, A.; Barłowska, J. Changes of protein content and its fractions in bovine milk from different breeds subject to somatic cell count. J. Dairy Sci. 2011, 94, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Król, J.; Brodziak, A.; Litwińczuk, Z.; Litwińczuk, A. Effect of age and stage of lactation on whey protein content in milk of cows of different breeds. Pol. J. Vet. Sci. 2013, 16, 395–397. [Google Scholar] [CrossRef]
- Garcia-Montoya, I.A.; Siqueiros-Cendon, T.; Arevalo-Gallegos, S.; Rascon-Cruz, Q. Lactoferrin multiple bioactive protein: An overview. Biochim. Biophys. Acta 2012, 1820, 226–236. [Google Scholar] [CrossRef]
- Cichosz, G.; Czeczot, H. The Nutritional Phenomenon of Milk; KZSM ZR Publishing: Olsztyn/Warszawa, Poland, 2013; pp. 50–75. [Google Scholar]
- Anghel, L.; Radulescu, A.; Erhan, R.V. Structural aspects of human lactoferrin in the iron-binding process studied by molecular dynamics and small-angle neutron scattering. EPJ 2018, 41, 9. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Boggs, I.; Weeks, M.; Li, Q.; Wu, H.; Harris, P.; Ma, Y.; Day, L. Kinetic modelling of the heat stability of bovine lactoferrin in raw whole milk. J. Food Eng. 2020, 280, 109977. [Google Scholar] [CrossRef]
- Conesa, C.; Sánchez, L.; Rota, C.; Pérez, M.D.; Calvo, M.; Farnaud, S.; Evans, R.W. Isolation of lactoferrin from milk of different species: Calorimetric and antimicrobial studies. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Matijašić, B.B.; Oberčkal, J.; Mohar Lorbeg, P.; Paveljšek, D.; Skale, N.; Kolenc, B.; Zupančič, J.M. Characterisation of lactoferrin isolated from acid whey using pilot-scale monolithic ion-exchange chromatography. Processes 2020, 8, 804. [Google Scholar] [CrossRef]
- Commission Implementing Decision of 22 November 2012 Authorising the Placing on the Market of Bovine Lactoferrin as a Novel Food Ingredient under Regulation (EC) No 258/97 of the European Parliament and of the Council (Friesland Campina) (Notified under Document C(2012) 8404). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32012D0727&from=PL (accessed on 16 May 2022).
- Wakabayashi, H.; Yamauchi, K.; Abe, F. Quality control of commercial bovine lactoferrin. BioMetals 2018, 31, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Conesa, C.; Calvo, M.; Sánchez, L. Recombinant human lactoferrin: A valuable protein for pharmaceutical products and functional foods. Biotechnol. Adv. 2010, 28, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Yemets, A.I.; Tanasienko, I.V.; Krasylenko, Y.A.; Blume, Y.B. Plant-based biopharming of recombinant human lactoferrin. Cell Biol. Int. 2014, 38, 989–1002. [Google Scholar] [CrossRef]
- Berkel, P.H.C.; Welling, M.M.; Geerts, M.; Veen, H.A.; Ravensbergen, B.; Salaheddine, M.; Pauwels, E.K.J.; Pieper, F.; Nuijens, J.H.; Nibbering, P.H. Large scale production of recombinant human. Nat. Biotechnol. 2002, 20, 484–487. [Google Scholar] [CrossRef]
- Tutykhina, I.L.; Bezborodova, O.A.; Shmarov, M.M.; Logunov, D.Y.; Neugodova, G.L.; Nemtsova, E.R.; Gintsburg, A.L. Production of recombinant human lactoferrin in the allantoic fluid of embryonated chicken eggs and its characteristics. Protein Expr. Purif. 2009, 65, 100–107. [Google Scholar] [CrossRef]
- Semak, I.; Budzevich, A.; Maliushkova, E. Development of dairy herd of transgenic goats as biofactory for large-scale production of biologically active recombinant human lactoferrin. Transgenic Res. 2019, 28, 465–478. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Cai, Y.; Xu, X.; Chen, J.; Wu, Y.; Cheng, G. Expression of active recombinant human lactoferrin in the milk of transgenic goats. Protein Expr. Purif. 2008, 57, 127–135. [Google Scholar] [CrossRef]
- Wajs, J.; Król, K.; Brodziak, A. Milk and dairy products as a source of antiviral compound. In Health and Life Style Economic, Social and Health Effects of a Pandemic; Nowak, W., Szalonka, K., Eds.; Wroclaw University Publishing: Wroclaw, Poland, 2021; pp. 413–425. [Google Scholar]
- Potì, F.; Pozzoli, C.; Adami, M.; Poli, E.; Costa, L.G. Treatments for COVID-19: Emerging drugs against the coronavirus. Acta Biomed. 2020, 91, 118–136. [Google Scholar]
- Gopinathan, J.; Hao, T.N.; Cha, E.; Lee, C.; Das, D.; Noh, I. 3D printable and injectable lactoferrin-loaded carboxymethyl cellulose-glycol chitosan hydrogels for tissue engineering applications. Mater. Sci. Eng. 2020, 113, 111008. [Google Scholar]
- Zimecki, M.; Artym, J.; Kocieba, M.; Kruzel, M.L. Effects of lactoferrin on elicitation of the antigen-specific cellular and humoral cutaneous response in mice. Postepy Hig. Med. Dosw. 2012, 66, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Fujishima, H.; Okada, N.; Matsumoto, K.; Shimizu, E.; Fukuda, S.; Tomita, M. Conjunctival injection reduction in patients with atopic keratoconjunctivitis due to synergic effect of bovine enteric-coated lactoferrin in 0.1% tacrolimus ophthalmic suspension. J. Clin. Med. 2020, 9, 3093. [Google Scholar] [CrossRef] [PubMed]
- Zimecki, M.; Artym, J.; Kocięba, M. Lactoferrin restrains allergen-induced pleurisy in mice. Inflamm. Res. 2012, 61, 1247–1255. [Google Scholar] [CrossRef] [Green Version]
- Negaoui, H.; El Mecherfi, K.E.; Tadjer, S.A.; Grar, H.; Kheroua, O.; Saidi, D. Bovine lactoferrin allergenicity as studied in murine model of allergy. Food Agric. Immunol. 2016, 27, 711–723. [Google Scholar] [CrossRef] [Green Version]
- Teng, C.T.; Gladwell, W. Single nucleotide polymorphisms (SNPs) in human lactoferrin gene. Biochem. Cell Biol. 2006, 84, 381–384. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Serrano, M.; Sabater, M.; Ortega, F.; Serino, M.; Pueyo, N.; Fernández-Real, J.M. Study of lactoferrin gene expression in human and mouse adipose tissue, human preadipocytes and mouse 3T3-L1 fibroblasts. Association with adipogenic and inflammatory markers. J. Nutr. Biochem. 2013, 24, 1266–1275. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Ortega, F.; Moreno, M.; Serrano, M.; Ricart, W.; Fernández-Real, J.M. Lactoferrin gene knockdown leads to similar effects to iron chelation in human adipocytes. J. Cell. Mol. Med. 2014, 18, 391–395. [Google Scholar] [CrossRef]
- Mohamed, J.A.; DuPont, H.L.; Jiang, Z.D.; Belkind-Gerson, J.; Figueroa, J.F.; Armitige, L.Y.; Okhuysen, P.C. A novel single-nucleotide polymorphism in the lactoferrin gene is associated with susceptibility to diarrhea in North American travelers to Mexico. Clin. Infect. Dis. 2007, 44, 945–952. [Google Scholar] [CrossRef]
- WHO 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 21 September 2021).
- Ono, T.; Morishita, S.; Murakoshi, M. Novel function of bovine lactoferrin in lipid metabolism: Visceral fat reduction by enteric-coated lactoferrin. Pharma Nutr. 2013, 1, 32–34. [Google Scholar] [CrossRef]
- Ono, T.; Murakoshi, M.; Uchiyama, A. Anti-obesity effect of lactoferrin; subgroup analysis excluding subjects with obese and/or hyper-LDL cholesterolemia. Immunol. Endocr. Metab. Agents Med. Chem. 2018, 18, 105–109. [Google Scholar] [CrossRef]
- ElSayed, H.L.; Abdelsayed, M.G.R.; Emara, I.S.A. Effect of lactoferrin supplementation on appetite and weight loss in obese school age children. QJM Int. J. Med. 2021, 114, hcab113.029. [Google Scholar] [CrossRef]
- Sun, J.; Ren, F.; Xiong, L.; Zhao, L.; Guo, H. Bovine lactoferrin suppresses high-fat diet induced obesity and modulates gut microbiota in C57BL/6J mice. J. Funct. Foods 2016, 22, 189–200. [Google Scholar] [CrossRef]
- Shi, J.; Finckenberg, P.; Martonen, E.; Ahlroos-Lehmus, A.; Pilvi, T.K.; Korpela, R.; Mervaala, E.M. Metabolic effects of lactoferrin during energy restriction and weight regain in diet-induced obese mice. J. Funct. Foods 2012, 4, 66–78. [Google Scholar] [CrossRef]
- Di Renzo, L.; Cioccoloni, G.; Falco, S.; Abenavoli, L.; Moia, A.; Sinibaldi Salimei, P.; De Lorenzo, A. Influence of FTO rs9939609 and Mediterranean diet on body composition and weight loss: A randomized clinical trial. J. Transl. Med. 2018, 16, 308. [Google Scholar] [CrossRef] [PubMed]
- Iłowiecka, K.; Glibowski, P.; Skrzypek, M.; Styk, W. The long-term dietitian and psychological support of obese patients who have reduced their weight allows them to maintain the effects. Nutrients 2021, 13, 2020. [Google Scholar] [CrossRef]
- Zhao, N.N.; Dong, G.; Wu, W.; Wang, J.L.; Ullah, R.; Fu, J.F. FTO gene polymorphisms and obesity risk in Chinese population: A meta-analysis. World J. Pediatr. 2019, 15, 382–389. [Google Scholar] [CrossRef]
- Aoun, C.; Hajj, A.; Hajj, F.; Papazian, T.; Rabbaa Khabbaz, L. The interaction between genetic polymorphisms in FTO, MC4R and MTHFR genes and adherence to the Mediterranean Diet in relation to obesity. Gene 2021, 809, 146037. [Google Scholar] [CrossRef]
- Holzbach, L.; Silveira, A.; Franco, L.; Horst, M.; Cominetti, C. Polymorphism PLIN1 11482 G>A interacts with dietary intake to modulate anthropometric measures and lipid profile in adults with normal-weight obesity syndrome. Br. J. Nutr. 2021, 128, 1004–1012. [Google Scholar] [CrossRef]
- Smith, C.E.; Tucker, K.L.; Yiannakouris, N.; Garcia-Bailo, B.; Mattei, J.; Lai, C.Q.; Ordovaás, J.M. Perilipin polymorphism interacts with dietary carbohydrates to modulate anthropometric traits in hispanics of Caribbean origin. J. Nutr. 2008, 138, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Yasueda, A.; Ito, T.; Maeda, K. Review: Evidence-based clinical research of anti-obesity supplements in Japan. Immunol. Endocr. Metab. Agents Med. Chem. 2013, 13, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 20 May 2022).
- Mohamed, W.A.; Schaalan, M.F. Antidiabetic efficacy of lactoferrin in type 2 diabetic pediatrics; controlling impact on PPAR-γ, SIRT-1, and TLR4 downstream signaling pathway. Diabetol. Metab. Syndr. 2018, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Alterations in concentration/activity of superoxide dismutases in context of obesity and selected single nucleotide polymorphisms in genes: SOD1, SOD2, SOD3. Int. J. Mol. Sci. 2020, 21, 5069. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, Ł.; Urbanowicz, I.; Kepinska, M.; Milnerowicz, H. Concentration/activity of superoxide dismutase isozymes and the pro-/antioxidative status, in context of type 2 diabetes and selected single nucleotide polymorphisms (genes: INS, SOD1, SOD2, SOD3)—Preliminary findings. Biomed. Parmacother. 2021, 137, 111396. [Google Scholar] [CrossRef]
- Guo, H.; Ding, J.; Liu, Q.; Li, Y.; Liang, J.; Zhang, Y. Vitamin C and metabolic syndrome: A meta-analysis of observational studies. Front. Nutr. 2021, 8, 728880. [Google Scholar] [CrossRef] [PubMed]
- Totan, B.; Baygut, H.; Karadağ, M.G. Vitamin C physiology: The known and the unknown in obesity. J. Food Nutr. Res. 2019, 7, 613–618. [Google Scholar] [CrossRef]
- Thomas-Valdés, S.; Tostes, M.D.G.V.; Anunciação, P.C.; da Silva, B.P.; Sant’Ana, H.M.P. Association between vitamin deficiency and metabolic disorders related to obesity. Crit. Rev. Food Sci. Nutr. 2016, 57, 3332–3343.a. [Google Scholar] [CrossRef]
- Michels, A.J.; Hagen, T.M.; Frei, B. Human genetic variation influences vitamin C homeostasis by altering vitamin C transport and antioxidant enzyme function. Ann. Rev. Nutr. 2013, 33, 45–70. [Google Scholar] [CrossRef] [Green Version]
- Eck, P. Nutrigenomics of vitamin C absorption and transport. Curr. Opin. Food Sci. 2018, 20, 100–104. [Google Scholar] [CrossRef]
- Xia, W.; Chen, N.; Peng, W.; Jia, X.; Yu, Y.; Wu, X.; Gao, H. Systematic meta-analysis revealed an association of PGC-1α rs8192678 polymorphism in type 2 diabetes mellitus. Dis. Markers 2019, 3, 2970401. [Google Scholar] [CrossRef] [Green Version]
- Hanan, S.A.; Najla, O.A.; Azza, M.M.; Iman, A.S.; Nouf, M.A. Modulating impacts of quercetin and/or lactoferrin on diabetic nephropathy and cardiomyopathy induced rats. Int. J. Pharm. Res. Allied Sci. 2017, 6, 128–137. [Google Scholar]
- Ling, C.J.; Min, Q.Q.; Yang, J.R.; Zhang, Z.; Yang, H.H.; Xu, J.Y.; Qin, L.Q. Lactoferrin alleviates the progression of atherosclerosis in ApoE−/− mice fed with high-fat/cholesterol diet through cholesterol homeostasis. J. Med. Food 2019, 22, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Zee, R.Y.; Cook, N.R.; Cheng, S.; Erlich, H.A.; Lindpaintner, K.; Ridker, P.M. Polymorphism in the beta2-adrenergic receptor and lipoprotein lipase genes as risk determinants for idiopathic venous thromboembolism: A multilocus, population-based, prospective genetic analysis. Circulation 2006, 113, 2193–2200. [Google Scholar] [CrossRef] [PubMed]
- Kvaløy, K.; Holmen, J.; Hveem, K.; Holmen, T.L. Genetic effects on longitudinal changes from healthy to adverse weight and metabolic status—The HUNT study. PLoS ONE 2015, 10, e0139632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivkar, R.R.; Gawade, G.C.; Padwal, M.K.; Diwan, A.G.; Mahajan, S.A.; Kadam, C.Y. Association of MTHFR C677T (rs1801133) and A1298C (rs1801131) polymorphisms with serum homocysteine, folate and vitamin B12 in patients with young coronary artery disease. Indian J. Clin. Biochem. 2022, 37, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Nozari, S.; Fathi Maroufi, N.; Nouri, M.; Paytakhti Oskouei, M.; Shiralizade, J.; Yekani, F.; Mamipour, M.; Faridvand, Y. Decreasing serum homocysteine and hypocholesterolemic effects of bovine lactoferrin in male rat fed with high-cholesterol diet. J. Cardiovasc. Thorac. Res. 2018, 10, 203–208. [Google Scholar]
- Fu, L.; Li, Y.N.; Luo, D.; Deng, S.; Wu, B.; Hu, Y.Q. Evidence on the causal link between homocysteine and hypertension from a meta-analysis of 40 173 individuals implementing Mendelian randomization. J. Clin. Hypertens. 2019, 21, 1879–1894. [Google Scholar] [CrossRef] [Green Version]
- Mayeur, S.; Spahis, S.; Pouliot, Y.; Levy, E. Lactoferrin, a pleiotropic protein in health and disease. Antioxid. Redox Signal. 2016, 24, 813–836. [Google Scholar] [CrossRef] [Green Version]
- Gkouskou, K.K.; Grammatikopoulou, M.G.; Vlastos, I.; Sanoudou, D.; Eliopoulos, A.G. Genotype-guided dietary supplementation in precision nutrition. Nutr. Rev. 2021, 79, 1225–1235. [Google Scholar] [CrossRef]
- Paesano, R.; Pacifici, E.; Benedetti, S.; Berlutti, F.; Frioni, A.; Polimeni, A.; Valenti, P. Safety and efficacy of lactoferrin versus ferrous sulphate in curing iron deficiency and iron deficiency anaemia in hereditary thrombophilia pregnant women: An interventional study. BioMetals 2014, 27, 999–1006. [Google Scholar] [CrossRef]
- Hashim, H.A.; Foda, O.; Ghayaty, E. Lactoferrin or ferrous salts for iron deficiency anemia in pregnancy: A meta-analysis of randomized trials. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 219, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Paesano, R.; Berlutti, F.; Pietropaoli, M.; Pantanella, F.; Pacifici, E.; Goolsbee, W.; Valenti, P. Lactoferrin efficacy versus ferrous sulfate in curing iron deficiency and iron deficiency anemia in pregnant women. Biol. Met. 2010, 23, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.; Meng, M.; Lan, Z.; Hua, L.; Ying, Z.; Humina, X.; Jia, S.; Weizheng, T.; Ping, Y.; Lingying, C.; et al. Iron metabolism in infants: Influence of bovine lactoferrin from iron-fortified formula. Nutrition 2015, 31, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.; Lepanto, M.L.; Cutone, A.; Siciliano, R.A.; Paesano, R.; Costi, R.; Musci, G.; Valenti, P. Influence of oral administration mode on the efficacy of commercial bovine Lactoferrin against iron and inflammatory homeostasis disorders. BioMetals 2020, 33, 159–168. [Google Scholar] [CrossRef]
- Legrand, D. Overview of lactoferrin as a natural immune modulator. J. Pediatr. 2016, 173S, S10–S15. [Google Scholar] [CrossRef] [Green Version]
- Mulder, A.M.; Morris, C.A. Lactoferrin in immune function, cancer and disease resistance. In Dietary Components and Immune Function. Nutrition and Health; Watson, R.R., Zibadi, S., Preedy, V.R., Eds.; Springer: Totowa, NJ, USA, 2010; p. 297316. [Google Scholar]
- Oda, H.; Wakabayashi, H.; Yamauchi, K.; Abe, F. Lactoferrin and bifidobacterial. BioMetals 2014, 27, 915–922. [Google Scholar] [CrossRef]
- Król, J.; Brodziak, A. Milk proteins with antibacterial properties. Probl. Higieny Epidemiol. 2015, 96, 399–405. [Google Scholar]
- Sekse, C.; Bohlin, J.; Skjerve, E.; Vegarud, G.E. Growth comparison of several Escherichia coli strains exposed to various concentrations of lactoferrin using linear spline regression. Microb. Inform. Exp. 2012, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Flores-Villaseñor, H.; Canizalez-Román, A.; Reyes-Lopez, M.; Zazueta-Beltrán, J.; León-Sicairos, N.; Bolscher, J.G. Bactericidal effect of bovine lactoferrin, LFcin, LFampin and LFchimera on antibiotic-resistant Staphylococcus aureus and Escherichia coli. Biometals 2010, 23, 569–578. [Google Scholar] [CrossRef]
- Ciccaglione, A.F.; Di Giulio, M.; Di Lodovico, S.; Di Campli, E.; Cellini, L.; Marzio, L. Bovine lactoferrin enhances the efficacy of levofloxacin-based triple therapy as first-line treatment of Helicobacter pylori infection: An in vitro and in vivo study. J. Antimicrob. Chemother. 2019, 74, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Välimaa, H.; Tenovuo, J.; Waris, M.; Hukkanen, V. Human lactoferrin but not lysozyme neutralizes HSV-1 and inhibits HSV-1 replication and cell-to-cell spread. Virol. J. 2009, 6, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuccotti, G.; Salvini, F.; Riva, E.; Agostoni, C. Oral lactoferrin in HIV-1 vertically infected children: An observational follow-up of plasma viral load and immune parameters. J. Int. Med. Res. 2006, 34, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EL-Fakharany, E.M.; Sánchez, L.; Al-Mehdar, H.A.; Redwan, E.M. Effectiveness of human, camel, bovine and sheep lactoferrin on the hepatitis C virus cellular infectivity: Comparison study. Virol. J. 2013, 10, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzik, S.; Miciński, B.; Miciński, J.; Mituniewicz, T.; Kowalski, P.M. Therapeutic properties of lactoferrin. Pol. Ann. Med. 2018, 25, 158–161. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, P.; Wang, H.; Luo, Y.; Wan, L.; Jiang, M.; Chu, Y. Lactoferrin for the treatment of COVID-19 (Review). Exp. Ther. Med. 2020, 20, 272. [Google Scholar] [CrossRef]
- Chang, R.; Ng, T.; Sun, W. Lactoferrin as potential preventative and adjunct treatment for COVID-19. Int. J. Antimicrob. Agents 2020, 56, 106118. [Google Scholar] [CrossRef]
- Hussain, F.; Rahim, K.; Rehman, S.U.; Shafqat, F.; Siddique, A.; Ahsan, M.; Shuaib, M.; Khan, I.U.; Shanyuan, C. Application of natural products in SARS-CoV-2. AP 2023, 15, 339–351. [Google Scholar]
- Mattar, E.H.; Elrashdy, F.; Almehdar, H.A.; Uversky, V.N.; Redwan, E.M. Natural resources to control COVID-19: Could lactoferrin amend SARS-CoV-2 infectivity? PeerJ 2021, 9, e11303. [Google Scholar] [CrossRef]
- Quantec 2021. Available online: https://www.quantec.co.nz/ (accessed on 19 June 2021).
- Stobiecka, M.; Król, J.; Brodziak, A. Antioxidant activity of milk and dairy products. Animals 2022, 12, 245. [Google Scholar] [CrossRef]
- Mulder, A.M.; Connellan, P.A.; Oliver, C.J.; Morris, C.A.; Stevenson, L.M. Bovine lactoferrin supplementation supports immune and antioxidant status in healthy human males. Nutr. Res. 2008, 28, 583–589. [Google Scholar] [CrossRef]
- Serrano, G.; Kochergina, I.; Albors, A.; Eva Diaz, E.; Oroval, M.; Hueso, G.; Serrano, J.M. Liposomal lactoferrin as potential prevention and cure for COVID-19. Int. J. Res. Health Sci. 2020, 8, 8–15. [Google Scholar]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Pleiotropic effect of lactoferrin in the prevention and treatment of COVID-19 infection: Randomized clinical trial, in vitro and in silico preliminary evidences. bioRxiv 2020. [CrossRef]
- Chichlowski, M.; Bokulich, N.; Harris, C.L.; Wampler, J.L.; Li, F.; Berseth, C.L.; Rudolph, C.; Wu, S.S. Effect of bovine milk fat globule membrane and lactoferrin in infant formula on gut microbiome and metabolome at 4 months of age. Curr. Dev. Nutr. 2021, 5, nzab027. [Google Scholar] [CrossRef] [PubMed]
- Friesland Campina 2021. Available online: https://www.frieslandcampina.com/news/frieslandcampina-ingredients-significantly-expands-its-lactoferrin-production-capacity/ (accessed on 19 October 2022).
- Dairy Reporter 2020. Available online: https://www.dairyreporter.com/Article/2020/10/29/Morinaga-Milk-expanding-lactoferrin-production-capacity-through-German-subsidiary (accessed on 19 June 2021).
- Grand View Research 2021. Available online: https://www.grandviewresearch.com/industry-analysis/lactoferrin-market (accessed on 19 October 2022).
- Market Data Forecast 2021a. Available online: https://www.marketdataforecast.com/market-reports/europe-lactoferrin-market (accessed on 19 October 2022).
- Market Data Forecast 2021b. Available online: https://www.marketdataforecast.com/market-reports/latin-america-lactoferrin-market (accessed on 30 November 2022).
- Market Research Report 2020. Available online: https://www.fortunebusinessinsights.com/industry-reports/bovine-lactoferrin-market-101656 (accessed on 19 October 2022).
- Bourbon, A.I.; Martins, J.T.; Pinheiro, A.C.; Madalena, D.A.; Marques, A.; Nunes, R.; Vicente, A.A. Nanoparticles of lactoferrin for encapsulation of food ingredients. In Biopolymer Nanostructures for Food Encapsulation Purposes; Jafari, S., Ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2019; pp. 147–168. [Google Scholar]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific opinion on bovine lactoferrin. EFSA J. 2012, 10, 2701–2727. [Google Scholar] [CrossRef] [Green Version]
- Commission Implementing Regulation (EU) 2018/1023 of 23 July 2018 correcting Implementing Regulation (EU) 2017/2470 establishing the Union list of novel foods (L 187). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32018R1023&from=EN (accessed on 10 May 2022).
- Li, F.; Wu, S.S.; Berseth, C.L.; Harris, C.L.; Richards, J.D.; Wampler, J.L.; Colombo, J. Improved neurodevelopmental outcomes associated with bovine milk fat globule membrane and lactoferrin in infant formula: A randomized, controlled trial. J. Pediatr. 2019, 215, 24–31.e8. [Google Scholar] [CrossRef]
- Büyükkiraz, M.E.; Kesmen, Z. Antimicrobial peptides (AMPs): A promising class of antimicrobial compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef]
- Tsukahara, T.; Fujimori, A.; Misawa, Y.; Oda, H.; Yamauchi, K.; Abe, F.; Nomiyama, T. The Preventive effect of lactoferrin-containing yogurt on gastroenteritis in nursery school children—Intervention study for 15 weeks. Int. J. Environ. Res. Public Health 2020, 17, 2534. [Google Scholar] [CrossRef]
- Barbiroli, A.; Bonomi, F.; Capretti, G.; Iametti, S.; Manzoni, M.; Piergiovanni, L.; Rollini, M. Antimicrobial activity of lysozyme and lactoferrin incorporated in cellulose-based food packaging. Food Control 2012, 26, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Soyer, F.; Keman, D.; Eroğlu, E.; Türe, H. Synergistic antimicrobial effects of activated lactoferrin and rosemary extract in vitro and potential application in meat storage. J. Food Sci. Technol. 2020, 57, 4395–4403. [Google Scholar] [CrossRef]
- Padrão, J.; Gonçalves, S.; Silva, J.P.; Sencadas, V.; Lanceros-Méndez, S.; Pinheiro, A.C.; Dourado, F. Bacterial cellulose-lactoferrin as an antimicrobial edible packaging. Food Hydrocoll. 2016, 58, 126–140. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Xiong, X.; Zhou, H.; Xiao, Q. Effect of lactoferrin on physicochemical properties and microstructure of pullulan-based edible films. J. Sci. Food Agric. 2019, 99, 4150–4157. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LF Form | Dosage | Research Assumptions | Effect of LF | References |
|---|---|---|---|---|
| bLF | 300 mg | Effects of bLF on systemic lipid metabolism and visceral fat reduction | A significant reduction in visceral fat area (VFA) was observed (−14.6 cm2), as well as a decrease in body weight (−1.5 kg), BMI (−0.6 kg/m2) and waist circumference (−4.4 cm) | [34] |
| Enteric-coated LF (eLF) | No data | Effect of LF supplementation on weight reduction (excluding subjects with BMI ≥ 30 and/or hyper-LDL cholesterolemia) | eLF caused a significant decrease in adipose tissue (−10.2 cm2, p < 0.01) and reduction in the average visceral fat content (−16.2 cm2, p < 0.01) and a significant decrease in BMI (−0.38 kg/m2, p < 0.05) | [35] |
| LF | 200 mg/day | Effect of LF on weight loss and appetite in obese school-age children | LF was shown to reduce appetite in obese school-aged children. It did not cause weight loss | [36] |
| bLF | 100 mg/kg BW | Effect of LF supplementation on the development of obesity on a high-fat diet (HF) in mice | bLF reduced the increase in body weight gain by 50%, decreased the total cholesterol concentration in the blood serum, and positively influenced the blood glucose level. No differences were noted in triacylglycerol content | [37] |
| LF | 15% of total protein source | LF prevents obesity and diet-induced development formation of hepatic steatosis in C57Bl/6J mice | LF supplementation was shown to improve weight loss and limit its subsequent regain, mitigated fatty liver, improved glucose tolerance, and reduced the inflammation of adipose tissue | [38] |
| LF Form | Dosage | Research Assumptions | Effect of LF | References |
|---|---|---|---|---|
| bLF | 200 mg/day | Effectiveness of bLF and ferrous sulphate in treating iron deficiency and iron-deficiency anaemia in pregnant women | Administration of 30%-iron-saturated bLF to pregnant women increased haemoglobin levels, erythrocyte counts, and levels of ferritin and total iron. bLF caused a decrease in the IL-6 level and an increase in the hepcidin concentration | [67] |
| bLF | 200 mg/day | Safety and efficacy of LF versus ferrous sulphate in pregnant women | Supplementation LF decreased the IL-6 level by 26% but increased by 21% in the group that received ferrous sulphate. No side effects were noted in the LF group, whereas in the ferrous sulphate group, they were reported by 16.5% of women | [65] |
| bLF | 38 mg bLF/100 g infant formula | Efficacy of lactoferrin-enriched infant formula in improving hematologic indices and iron status | The body weight of the infants receiving bLF was significantly higher than that in the group that received iron. Milk enriched with bLF significantly increased the concentration of haemoglobin (Hb) by 12%, ferritin by 76%, and total iron by 29%, significantly decreasing the detection rate of anaemia from 12% to 5% and of iron-deficiency anaemia from 35% to 16% | [68] |
| bLF | 200 mg/day | Evaluation of the effectiveness of LF administration before and after a meal. The possible influence of digestive protease on LF lactoferrin degradation | An increase in serum haematological parameters, i.e., haemoglobin, erythrocytes, ferritin and iron, was observed only in the group in which the bLF supplement was administered before meals | [69] |
| LF Form | Dosage | Research Assumptions | Effect of LF | References |
|---|---|---|---|---|
| bLF | Placebo—first week 100 mg—second week 200 mg—third week | Potential immune-modulating properties and antioxidant activity of oral supplementation of bovine lactoferrin in humans. | After two weeks of bLF supplementation, the total T cell count had significantly increased by 29%, helper T cells by 22%, and cytotoxic T cells by 25%. The number of NK cells was not affected by the supplementation. An important finding was the significant increase in antioxidant capacity after two weeks of LF supplementation | [87] |
| Liposomal bovine LF (LLF) | 128–192 mg/day | LLF as potential prevention and cure for COVID-19. | Administration of LLF led to 100% recovery in 75 SARS-CoV-2-positive patients within 4–5 days | [88] |
| LLF | 1 g/day | Evaluation of the antiviral effect of oral and nasal LLF in asymptomatic and mild to moderate COVID-19 patients. | Faster recovery from COVID-19 in patients taking LLF | [89] |
| bLF | 0.6 g/L | Comparison of the gut microbiome profile and its metabolites in infants up to 12 months of age who received formula containing bLF and MMGF (milk fat globules) or regular formula without these additives. | Infants fed the formula with bLF and MMGF showed an increase in numbers of Bacteroides uniformis and Bacteroides plebeius and of bifidobacteria in the faeces, accompanied by a decrease in numbers of E. coli, in comparison to infants receiving the standard formula up to 4 months of age | [90] |
| Producer | Country |
|---|---|
| MILEI GmbH | Leutkirch im Allgäu, Germany |
| FrieslandCampina DOMO | Amersfoort, The Netherlands |
| Synlait Ltd. | Dunsandel, New Zealand |
| Glanbia Plc. | Kilkenny, Ireland |
| Bega Bionutrients | Port Melbourne, VIC, Australia |
| Saputo Dairy Australia Pty Ltd. | Allansford, VIC, Australia |
| Fonterra Co-operative Group | Auckland, New Zealand |
| Armor Protéines SAS | Loudéac, France |
| Hilmar Cheese Co. | Hilmar, CA, USA |
| Murray Goulburn Co-operative Co. Ltd. | Southbank, VIC, Australia |
| Ingredia | Arras, France |
| Product Type | Product Name | Lactoferrin Content | Use | Web Page Link |
|---|---|---|---|---|
| Cream | Endvir Simplex (Vitis Pharma) | 6% |
| http://vitispharma.pl/katalog/produkt/produkty/endvir/endvir-simplex (accessed on 16 May 2022) |
| Cream | Acnex (Farmina Ltd.) | No data |
| https://farmina.pl/product/acnex-krem/ (accessed on 16 May 2022) |
| Capsules | Lactoferrin (Pharmabest) | 100 mg/capsule |
| https://pharmabest.pl/sklep/produkt/laktoferyna-kapsulki/ (accessed on 16 May 2022) |
| Capsules | Lactoferrin (Jarrow Formulas) | 250 mg/capsule |
| https://jarrow.com/products/lactoferrin-250-mg-60-capsules (accessed on 16 May 2022) |
| Capsules | IronSorb + Lactoferrin (Jarrow Formulas) | 200 mg/capsule |
| https://jarrow.com/products/ironsorb-lactoferrin-60-capsules (accessed on 16 May 2022) |
| Capsules | Lactoferrin LFS 90% 100 mg (Aliness—MedicaLine) | 100 mg/capsule |
| https://aliness.pl/pl/p/Lactoferrin-LFS-90-100-mg-x-30-kapsulek/228 (accessed on 16 May 2022) |
| Oral drops | Lactoferrin (Pharmabest) | 100 mg/12 drops |
| https://pharmabest.pl/laktoferyna-krople-doustne/ (accessed on 16 May 2022) |
| Sachets | Lactoferrin (Pharmabest) | 100 mg/sachet |
| https://pharmabest.pl/sklep/produkt/laktoferyna-saszetki/ (accessed on 16 May 2022) |
| Sachets | Fibraxine (Alfasigma) | 50 mg/sachet |
| https://www.alfasigma.com/ (accessed on 16 May 2022) |
| Dragées | Fiorda Junior (PhytoPharm) | 6 mg/dragée |
| http://phytopharm.pl/en/fiorda (accessed on 16 May 2022) |
| Toothpaste | Lactoferrin Toothpaste (DENTE91) | No data |
| https://dente91.com/dente91 (accessed on 16 May 2022) |
| Mouthwash | Lactoferrin Mouthwash (DENTE91) | No data |
| https://dente91.com/dente-91 (accessed on 16 May 2022) |
| Food Category | Maximum Level of bLF |
|---|---|
| Infant formulae and follow-on formulae within the meaning of Regulation (EU) No 609/2013 (ready to drink) | 100 mg/100 mL |
| Dairy-based foods for small children (ready-to-eat) | 200 mg/100 g |
| Processed cereal products (solid) | 670 mg/100 g |
| Food for special medical purposes within the meaning of Regulation (EU) No 609/2013 | Up to 3 g/day |
| Milk-based beverages | 200 mg/100 g |
| Powdered milk-based drink mixes (ready-to-drink) | 330 mg/100 g |
| Beverages based on fermented milk (including yoghurt drinks) | 50 mg/100 g |
| Non-alcoholic drinks | 120 mg/100 g |
| Yoghurt-based products | 80 mg/100 g |
| Cheese-based products | 2000 mg/100 g |
| Ice cream | 130 mg/100 g |
| Cakes and pastries | 1000 mg/100 g |
| Candies | 750 mg/100 g |
| Chewing gum | 3000 mg/100 g |
| Product Type | Product Name | Lactoferrin Content | Country of Manufacture | Use | Web Page Link |
|---|---|---|---|---|---|
| Modified powdered milk (up to 1 year of age) | Morinaga Hagukumi (Morinaga Milk Industry Co., Ltd) | 80 mg/100 g powder | Tokio, Japan |
| https://www.morinagamilk.co.jp/english/products/jp/infantformula.php (accessed on 17 May 2022) |
| Modified powdered milk (up to 1 year of age) | Enfamil Enspire Infant Formula (Mead Johnson & Company) | <2% | Chicago, IL, USA |
| https://www.enfamil.com/products/enfamil-enspire-infant-formula/ (accessed on 17 May 2022) |
| Mleko Modified powdered milk (from 12 months to 3 years of age) | Morinaga Chil-mil (Morinaga Milk Industry Co., Ltd) | 55 mg/100 g powder | Tokio, Japan |
| https://www.morinagamilk.co.jp/english/products/jp/infantformula.php (accessed on 17 May 2022) |
| Yoghurt | Lactoferrin Yogurt (Morinaga Milk Industry Co., Ltd) | 100 mg | Tokio, Japan |
| https://www.morinagamilk.co.jp/english/products/jp/yogurt.php (accessed on 17 May 2022) |
| Chewing gum with lactoferrin and manuka honey | Blue®m dental chewing gum (Blue®m) | No data | VH Wijhe, Netherlands |
| https://bluemcare.com/product/dental-chewing-gum/ (accessed on 17 May 2022) |
| Carbonated drink with lactoferrin | Immune + (Beston Global Foods) | No data | Adelaide, SA, Australia |
| https://immuneplus.com.au/ (accessed on 17 May 2022) |
| Fruit gums for children | C + Zinc Jelly (NANA and TAKA) | No data | Taiwan |
| https://www.hktvmall.com/hktv/en/main/Hong-Kong-Chien-Cao-Tong-Medical-Limited/s/H6449002/Personal-Care-%26-Health/Personal-Care-%26-Health/Health/Immunity/CZinc-Jelly/p/H6449002_S_CCT10867?scrollTo=recommendationTab (accessed on 17 May 2022) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jańczuk, A.; Brodziak, A.; Czernecki, T.; Król, J. Lactoferrin—The Health-Promoting Properties and Contemporary Application with Genetic Aspects. Foods 2023, 12, 70. https://doi.org/10.3390/foods12010070
Jańczuk A, Brodziak A, Czernecki T, Król J. Lactoferrin—The Health-Promoting Properties and Contemporary Application with Genetic Aspects. Foods. 2023; 12(1):70. https://doi.org/10.3390/foods12010070
Chicago/Turabian StyleJańczuk, Anna, Aneta Brodziak, Tomasz Czernecki, and Jolanta Król. 2023. "Lactoferrin—The Health-Promoting Properties and Contemporary Application with Genetic Aspects" Foods 12, no. 1: 70. https://doi.org/10.3390/foods12010070
APA StyleJańczuk, A., Brodziak, A., Czernecki, T., & Król, J. (2023). Lactoferrin—The Health-Promoting Properties and Contemporary Application with Genetic Aspects. Foods, 12(1), 70. https://doi.org/10.3390/foods12010070