

Kimchi and Leuconostoc mesenteroides DRC 1506 Alleviate Dextran Sulfate Sodium (DSS)-Induced Colitis via Attenuating Inflammatory Responses

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Culture of Leu. mesenteroides
2.2. Preparation of Kimchi Samples
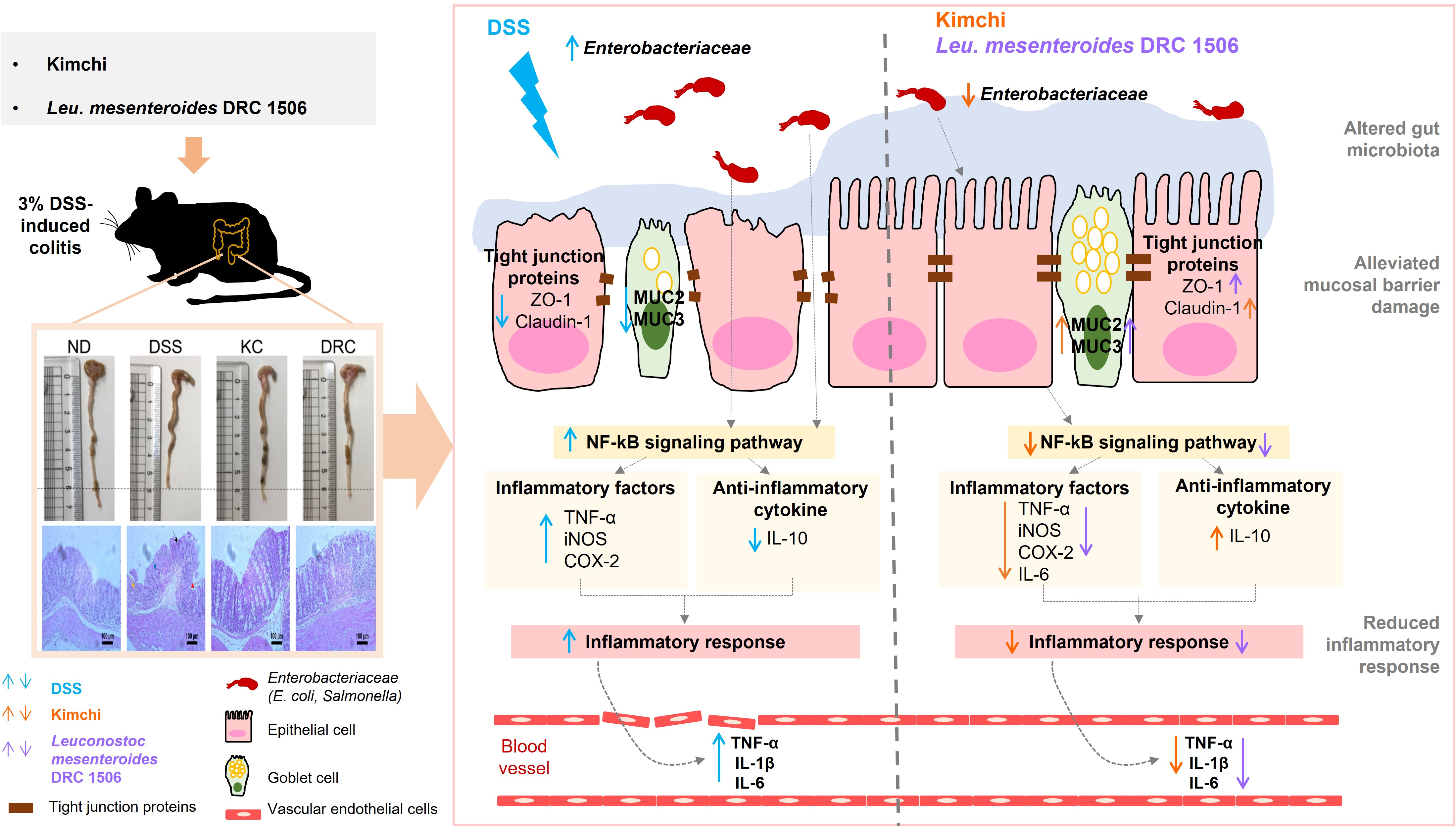
2.3. Experimental Design Inducing Colitis with DSS
2.4. Disease Activity Index (DAI)
2.5. Colonic Histological Damage
2.6. Microbial Count in the Feces
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Quantitative Real-Time Polymerase Chain Reactions (qRT-PCR)
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. Effects of Kimchi and DRC on Body Weight and Colitis Symptoms
3.2. Effects of Kimchi and DRC on Colonic Tissue Damage
3.3. Effects of Kimchi and DRC on Some Gut Microbial Counts
3.4. Effects of Kimchi and DRC on Inflammation-Related Cytokines in Serum and the Colon
3.5. Effects of Kimchi and DRC on the Expression of Genes and Proteins Related to the Inflammatory Response in the Colon
3.6. Effects of Kimchi and DRC on the Expression of Genes and Proteins Related to Mucosal Barrier Function in the Colon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ananthakrishnan, A.N.; Kaplan, G.G.; Ng, S.C. Changing global epidemiology of inflammatory bowel diseases: Sustaining health care delivery into the 21st century. Clin. Gastroenterol. Hepatol. 2020, 18, 1252–1260. [Google Scholar] [CrossRef] [PubMed]
- Mak, W.Y.; Zhao, M.; Ng, S.C.; Burisch, J. The epidemiology of inflammatory bowel disease: East meets west. J. Gastroenterol. Hepatol. 2020, 35, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Ramos, G.P.; Papadakis, K.A. Mechanisms of disease: Inflammatory bowel diseases. Mayo Clinic Proc. 2019, 94, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddavide, R.; Rotolo, O.; Caruso, M.G.; Stasi, E.; Notarnicola, M.; Miraglia, C.; Nouvenne, A.; Meschi, T.; De’Angelis, G.L.; Di Mario, F.; et al. The role of diet in the prevention and treatment of inflammatory bowel diseases. Acta Biomed. 2018, 89, 60–75. [Google Scholar] [PubMed]
- Park, S.H.; Kim, Y.J.; Rhee, K.H.; Kim, Y.H.; Hong, S.N.; Kim, K.H.; Seo, S.I.; Cha, J.M.; Park, S.Y.; Jeong, S.K.; et al. A 30-year trend analysis in the epidemiology of inflammatory bowel disease in the songpa-kangdong district of seoul, korea in 1986–2015. J. Crohns. Colitis 2019, 13, 1410–1417. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, K.; Feuerstein, J.D. New developments in ulcerative colitis: Latest evidence on management, treatment, and maintenance. Drugs. Context 2019, 8, 212572. [Google Scholar] [CrossRef]
- Wehkamp, J.; Stange, E.F. Recent advances and emerging therapies in the non-surgical management of ulcerative colitis. F1000Res. 2018, 7, 1207. [Google Scholar] [CrossRef]
- Zeng, W.; He, D.; Xing, Y.; Liu, J.; Su, N.; Zhang, C.; Wang, Y.; Xing, X. Internal connections between dietary intake and gut microbiota homeostasis in disease progression of ulcerative colitis: A review. Food Sci. Hum. Wellness 2021, 10, 119–130. [Google Scholar] [CrossRef]
- Keshteli, A.H.; Madsen, K.L.; Dieleman, L.A. Diet in the pathogenesis and management of ulcerative colitis; a review of randomized controlled dietary interventions. Nutrients 2019, 11, 1498. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of gut microbiota in inflammatory bowel disease (ibd): Cause or consequence? Ibd treatment targeting the gut microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef]
- Yousefi, B.; Eslami, M.; Ghasemian, A.; Kokhaei, P.; Salek Farrokhi, A.; Darabi, N. Probiotics importance and their immunomodulatory properties. J. Cell Physiol. 2019, 234, 8008–8018. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Kimchi microflora: History, current status, and perspectives for industrial kimchi production. Appl. Microbiol. Biotechnol. 2014, 98, 2385–2393. [Google Scholar] [CrossRef]
- Park, K.Y.; Jeong, J.K.; Lee, Y.E.; Daily, J.W., 3rd. Health benefits of kimchi (korean fermented vegetables) as a probiotic food. J. Med. Food 2014, 17, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Lee, H.J.; Jung, J.Y.; Lee, S.H.; Seo, H.Y.; Park, W.S.; Jeon, C.O. Effects of red pepper powder on microbial communities and metabolites during kimchi fermentation. Int. J. Food Microbiol. 2013, 160, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Kim, J.M.; Park, M.S.; Bae, J.W.; Hahn, Y.; Madsen, E.L.; Jeon, C.O. Metagenomic analysis of kimchi, a traditional korean fermented food. Appl. Environ. Microbiol. 2011, 77, 2264–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Seo, H.Y.; Park, W.S.; Jeon, C.O. Effects of Leuconostoc mesenteroides starter cultures on microbial communities and metabolites during kimchi fermentation. Int. J. Food Microbiol. 2012, 153, 378–387. [Google Scholar] [CrossRef]
- Park, E.J.; Chun, J.; Cha, C.J.; Park, W.S.; Jeon, C.O.; Bae, J.W. Bacterial community analysis during fermentation of ten representative kinds of kimchi with barcoded pyrosequencing. Food Microbiol. 2012, 30, 197–204. [Google Scholar] [CrossRef]
- Park, S.E.; Seo, S.H.; Kim, E.J.; Byun, S.; Na, C.S.; Son, H.S. Changes of microbial community and metabolite in kimchi inoculated with different microbial community starters. Food Chem. 2019, 274, 558–565. [Google Scholar] [CrossRef]
- Song, H.S.; Whon, T.W.; Kim, J.; Lee, S.H.; Kim, J.Y.; Kim, Y.B.; Choi, H.J.; Rhee, J.K.; Roh, S.W. Microbial niches in raw ingredients determine microbial community assembly during kimchi fermentation. Food Chem. 2020, 318, 126481. [Google Scholar] [CrossRef]
- Jeon, H.H.; Kim, K.H.; Chun, B.H.; Ryu, B.H.; Han, N.S.; Jeon, C.O. A proposal of Leuconostoc mesenteroides subsp. jonggajibkimchii subsp. Nov. and reclassification of Leuconostoc mesenteroides subsp. suionicum (gu et al., 2012) as Leuconostoc suionicum sp. nov. based on complete genome sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2225–2230. [Google Scholar] [CrossRef]
- Moon, S.H.; Kim, C.R.; Chang, H.C. Heterofermentative lactic acid bacteria as a starter culture to control kimchi fermentation. Lwt 2018, 88, 181–188. [Google Scholar] [CrossRef]
- Heo, W.; Lee, E.S.; Cho, H.T.; Kim, J.H.; Lee, J.H.; Yoon, S.M.; Kwon, H.T.; Yang, S.; Kim, Y.J. Lactobacillus plantarum LRCC 5273 isolated from kimchi ameliorates diet-induced hypercholesterolemia in c57bl/6 mice. Biosci. Biotechnol. Biochem. 2018, 82, 1964–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Moon, J.S.; Lee, M.-G.; Han, N.S. Immunomodulatory effects of Leuconostoc citreum EFELl2061 isolated from kimchi, a traditional korean food, on the th2 type-dominant immune response in vitro and in vivo. J. Funct. Foods 2016, 20, 79–87. [Google Scholar] [CrossRef]
- Kwon, M.-S.; Shin, M.-Y.; Lim, S.K.; Lee, J.; Park, H.K.; Kim, N.; Yun, M.; Jo, H.E.; Oh, Y.J.; Choi, H.-J. Leuconostoc citreum isolated from kimchi suppresses the development of collagen-induced arthritis in dba/1 mice. J. Funct. Foods 2019, 63, 103579. [Google Scholar] [CrossRef]
- Park, J.E.; Oh, S.H.; Cha, Y.S. Lactobacillus brevis OPK-3 from kimchi prevents obesity and modulates the expression of adipogenic and pro-inflammatory genes in adipose tissue of diet-induced obese mice. Nutrients 2020, 12, 604. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.S.; Lee, N.K.; Choi, A.J.; Choe, J.S.; Bae, C.H.; Paik, H.D. Anti-inflammatory potential of probiotic strain Weissella cibaria JW15 isolated from kimchi through regulation of nf-kappab and mapks pathways in lps-induced raw 264.7 cells. J. Microbiol. Biotechnol. 2019, 29, 1022–1032. [Google Scholar] [CrossRef]
- Jang, S.E.; Hyam, S.R.; Han, M.J.; Kim, S.Y.; Lee, B.G.; Kim, D.H. Lactobacillus brevis G-101 ameliorates colitis in mice by inhibiting nf-kappab, mapk and Akt pathways and by polarizing m1 macrophages to m2-like macrophages. J. Appl. Microbiol. 2013, 115, 888–896. [Google Scholar] [CrossRef]
- Jo, S.G.; Noh, E.J.; Lee, J.Y.; Kim, G.; Choi, J.H.; Lee, M.E.; Song, J.H.; Chang, J.Y.; Park, J.H. Lactobacillus curvatus WiKim38 isolated from kimchi induces il-10 production in dendritic cells and alleviates dss-induced colitis in mice. J. Microbiol. 2016, 54, 503–509. [Google Scholar] [CrossRef]
- Park, J.S.; Joe, I.; Rhee, P.D.; Jeong, C.S.; Jeong, G. A lactic acid bacterium isolated from kimchi ameliorates intestinal inflammation in dss-induced colitis. J. Microbiol. 2017, 55, 304–310. [Google Scholar] [CrossRef]
- Choi, H.Y.; Ryu, B.H.; Kim, D.H. Leuconostoc mesenteroides DRC1506 and Use Thereof. Korean Patent 101809447, 11 December 2017. [Google Scholar]
- Mennigen, R.; Nolte, K.; Rijcken, E.; Utech, M.; Loeffler, B.; Senninger, N.; Bruewer, M. Probiotic mixture vsl#3 protects the epithelial barrier by maintaining tight junction protein expression and preventing apoptosis in a murine model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1140–G1149. [Google Scholar]
- Kim, K.A.; Gu, W.; Lee, I.A.; Joh, E.H.; Kim, D.H. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the tlr4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef] [PubMed]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented foods: Definitions and characteristics, impact on the gut microbiota and effects on gastrointestinal health and disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, J.; Koseki, T.; Watanabe, K.; Ardiansyah; Budijanto, S.; Oikawa, A.; Alauddin, M.; Goto, T.; Aso, H.; Komai, M.; et al. Dietary supplementation of fermented rice bran effectively alleviates dextran sodium sulfate-induced colitis in mice. Nutrients 2017, 9, 747. [Google Scholar] [CrossRef]
- Jang, S.-H.; Park, J.; Kim, S.-H.; Choi, K.-M.; Ko, E.-S.; Cha, J.-D.; Lee, Y.-R.; Jang, H.; Jang, Y.-S. Oral administration of red ginseng powder fermented with probiotic alleviates the severity of dextran-sulfate sodium-induced colitis in a mouse model. Chin. J. Nat. Med. 2017, 15, 192–201. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, H.; Liu, H.; Cheng, H.; Pan, L.; Hu, M.; Li, X. Goji berry juice fermented by probiotics attenuates dextran sodium sulfate-induced ulcerative colitis in mice. J. Funct. Foods 2021, 83, 104491. [Google Scholar] [CrossRef]
- Okada, Y.; Tsuzuki, Y.; Takeshi, T.; Furuhashi, H.; Higashiyama, M.; Watanabe, C.; Shirakabe, K.; Kurihara, C.; Komoto, S.; Tomita, K.; et al. Novel probiotics isolated from a japanese traditional fermented food, funazushi, attenuates dss-induced colitis by increasing the induction of high integrin alphav/beta8-expressing dendritic cells. J. Gastroenterol. 2018, 53, 407–418. [Google Scholar] [CrossRef]
- Lee, H.; Yoon, H.; Ji, Y.; Kim, H.; Park, H.; Lee, J.; Shin, H.; Holzapfel, W. Functional properties of Lactobacillus strains isolated from kimchi. Int. J. Food Microbiol. 2011, 145, 155–161. [Google Scholar] [CrossRef]
- Lee, K.W.; Shim, J.M.; Park, S.-K.; Heo, H.-J.; Kim, H.-J.; Ham, K.-S.; Kim, J.H. Isolation of lactic acid bacteria with probiotic potentials from kimchi, traditional korean fermented vegetable. LWT-Food Sci. Technol. 2016, 71, 130–137. [Google Scholar] [CrossRef]
- Yang, S.J.; Lee, J.E.; Lim, S.M.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Antioxidant and immune-enhancing effects of probiotic Lactobacillus plantarum 200655 isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 491–499. [Google Scholar] [CrossRef]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Rodriguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Utrilla, M.P.; Chueca, N.; Garcia, F.; Rodriguez-Cabezas, M.E.; Galvez, J. Intestinal anti-inflammatory effect of the probiotic Saccharomyces boulardii in dss-induced colitis in mice: Impact on micrornas expression and gut microbiota composition. J. Nutr. Biochem. 2018, 61, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Chen, Y.; Wang, G.; Yang, Y.; Song, X.; Xiong, Z.; Zhang, H.; Lai, P.; Wang, S.; Ai, L. Lactobacillus plantarum AR113 alleviates dss-induced colitis by regulating the tlr4/myd88/nf-κb pathway and gut microbiota composition. J. Funct. Foods 2020, 67, 103854. [Google Scholar] [CrossRef]
- Seo, K.; Seo, J.; Yeun, J.; Choi, H.; Kim, Y.I.; Chang, S.Y. The role of mucosal barriers in human gut health. Arch. Pharm. Res. 2021, 44, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Rabah, H.; do Carmo, F.L.R.; Carvalho, R.D.O.; Cordeiro, B.F.; da Silva, S.H.; Oliveira, E.R.; Lemos, L.; Cara, D.C.; Faria, A.M.C.; Garric, G.; et al. Beneficial propionibacteria within a probiotic emmental cheese: Impact on dextran sodium sulphate-induced colitis in mice. Microorganisms 2020, 8, 380. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xie, Q.; Zhang, Y.; Ma, W.; Ning, K.; Xiang, J.Y.; Cui, J.; Xiang, H. Combination of probiotics with different functions alleviate dss-induced colitis by regulating intestinal microbiota, il-10, and barrier function. Appl. Microbiol. Biotechnol. 2020, 104, 335–349. [Google Scholar] [CrossRef]
- Baldelli, V.; Scaldaferri, F.; Putignani, L.; Del Chierico, F. The role of Enterobacteriaceae in gut microbiota dysbiosis in inflammatory bowel diseases. Microorganisms 2021, 9, 697. [Google Scholar] [CrossRef]
- Zhou, X.; Lu, Q.; Kang, X.; Tian, G.; Ming, D.; Yang, J. Protective role of a new polysaccharide extracted from Lonicera japonica Thunb in mice with ulcerative colitis induced by dextran sulphate sodium. Biomed Res. Int. 2021, 2021, 8878633. [Google Scholar] [CrossRef]
- Peran, L.; Camuesco, D.; Comalada, M.; Bailon, E.; Henriksson, A.; Xaus, J.; Zarzuelo, A.; Galvez, J. A comparative study of the preventative effects exerted by three probiotics, Bifidobacterium lactis, Lactobacillus casei and Lactobacillus acidophilus, in the tnbs model of rat colitis. J. Appl. Microbiol. 2007, 103, 836–844. [Google Scholar] [CrossRef]
- Liu, M.; Ding, J.; Zhang, H.; Shen, J.; Hao, Y.; Zhang, X.; Qi, W.; Luo, X.; Zhang, T.; Wang, N. Lactobacillus casei LH23 modulates the immune response and ameliorates dss-induced colitis via suppressing jnk/p-38 signal pathways and enhancing histone h3k9 acetylation. Food Funct. 2020, 11, 5473–5485. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. Nf-kappab: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Hanada, T.; Yoshimura, A. Regulation of cytokine signaling and inflammation. Cytokine. Growth Factor Rev. 2002, 13, 413–421. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.; Shin, J.-S.; Lee, W.-S.; Rhee, Y.K.; Cho, C.-W.; Hong, H.-D.; Lee, K.-T. Anti-colitis effect of Lactobacillus sakei K040706 via suppression of inflammatory responses in the dextran sulfate sodium-induced colitis mice model. J. Funct. Foods 2017, 29, 256–268. [Google Scholar] [CrossRef]
- Yan, S.; Yang, B.; Zhao, J.; Zhao, J.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, W. A ropy exopolysaccharide producing strain Bifidobacterium longum subsp. longum YS108R alleviates dss-induced colitis by maintenance of the mucosal barrier and gut microbiota modulation. Food Funct. 2019, 10, 1595–1608. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bifidobacteria (log CFU/g) | Lactobacillus (log CFU/g) | Enterobacteriaceae (log CFU/g) | |
|---|---|---|---|
| ND | 7.65 ± 0.12 b | 6.60 ± 0.76 b | 2.85 ± 2.47 b |
| DSS | 9.64 ± 0.25 a | 8.77 ± 0.28 a | 7.32 ± 0.19 a |
| KC | 9.83 ± 0.18 a | 8.99 ± 0.32 a | 3.91 ± 1.14 b |
| DRC | 9.50 ± 0.61 a | 8.68 ± 1.52 a | 5.38 ± 0.73 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, H.-J.; Oh, S.-H.; Park, K.-B.; Cha, Y.-S. Kimchi and Leuconostoc mesenteroides DRC 1506 Alleviate Dextran Sulfate Sodium (DSS)-Induced Colitis via Attenuating Inflammatory Responses. Foods 2023, 12, 584. https://doi.org/10.3390/foods12030584
Moon H-J, Oh S-H, Park K-B, Cha Y-S. Kimchi and Leuconostoc mesenteroides DRC 1506 Alleviate Dextran Sulfate Sodium (DSS)-Induced Colitis via Attenuating Inflammatory Responses. Foods. 2023; 12(3):584. https://doi.org/10.3390/foods12030584
Chicago/Turabian StyleMoon, Hye-Jung, Suk-Heung Oh, Ki-Bum Park, and Youn-Soo Cha. 2023. "Kimchi and Leuconostoc mesenteroides DRC 1506 Alleviate Dextran Sulfate Sodium (DSS)-Induced Colitis via Attenuating Inflammatory Responses" Foods 12, no. 3: 584. https://doi.org/10.3390/foods12030584
APA StyleMoon, H.-J., Oh, S.-H., Park, K.-B., & Cha, Y.-S. (2023). Kimchi and Leuconostoc mesenteroides DRC 1506 Alleviate Dextran Sulfate Sodium (DSS)-Induced Colitis via Attenuating Inflammatory Responses. Foods, 12(3), 584. https://doi.org/10.3390/foods12030584