

Fungal Biomarkers in Traditional Starter Determine the Chemical Characteristics of Turbid Rice Wine from the Rim of the Sichuan Basin, China
 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
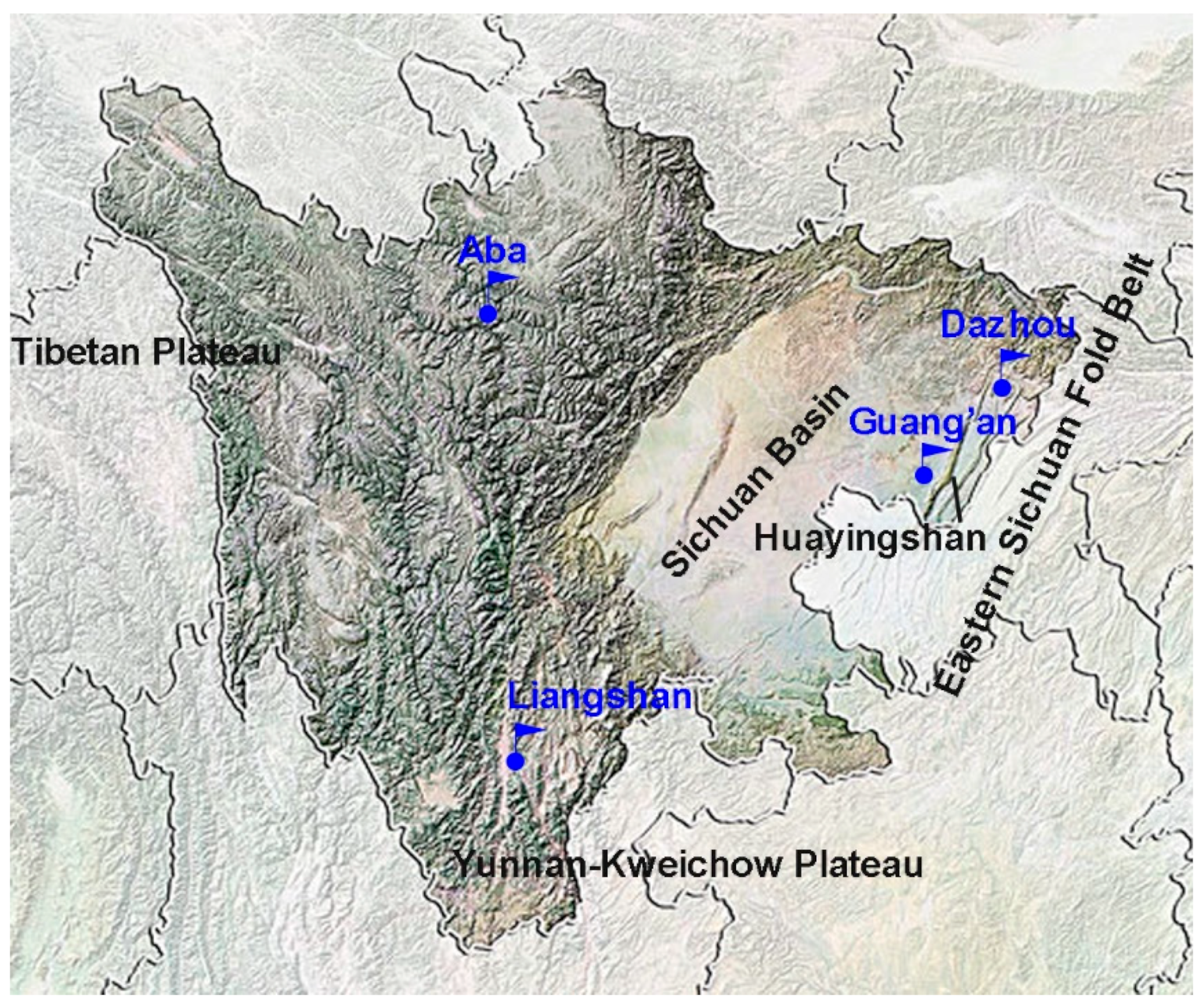
2.1. Sample Collection
2.2. DNA Extraction and Sequencing
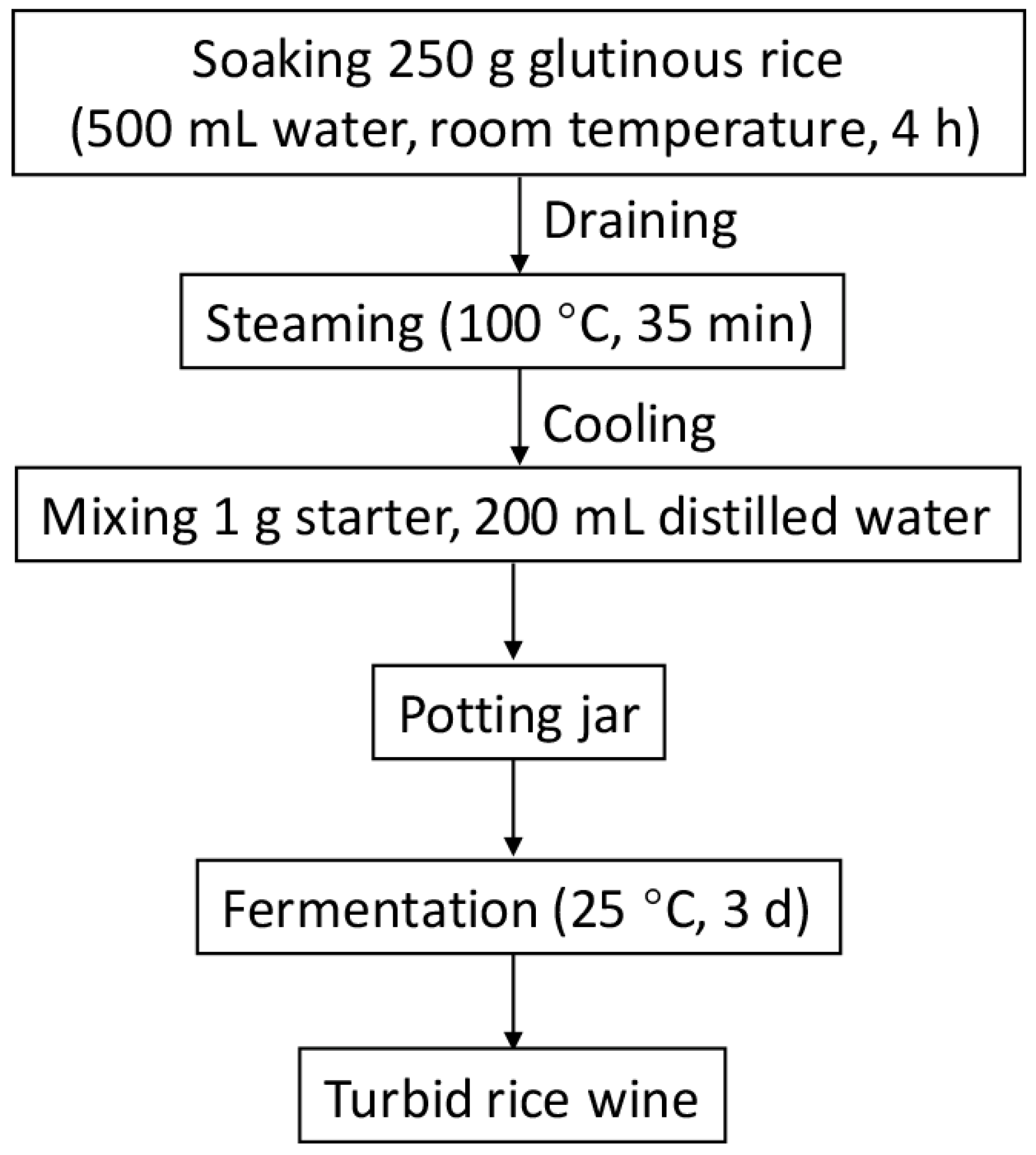
2.3. Rice Wine Fermentation on a Lab-Scale
2.4. Reducing Sugar and Ethanol Analysis
2.5. Non-Volatile Organic Acids Analysis
2.6. Volatile Compounds Analysis
2.7. Statistical Analyses
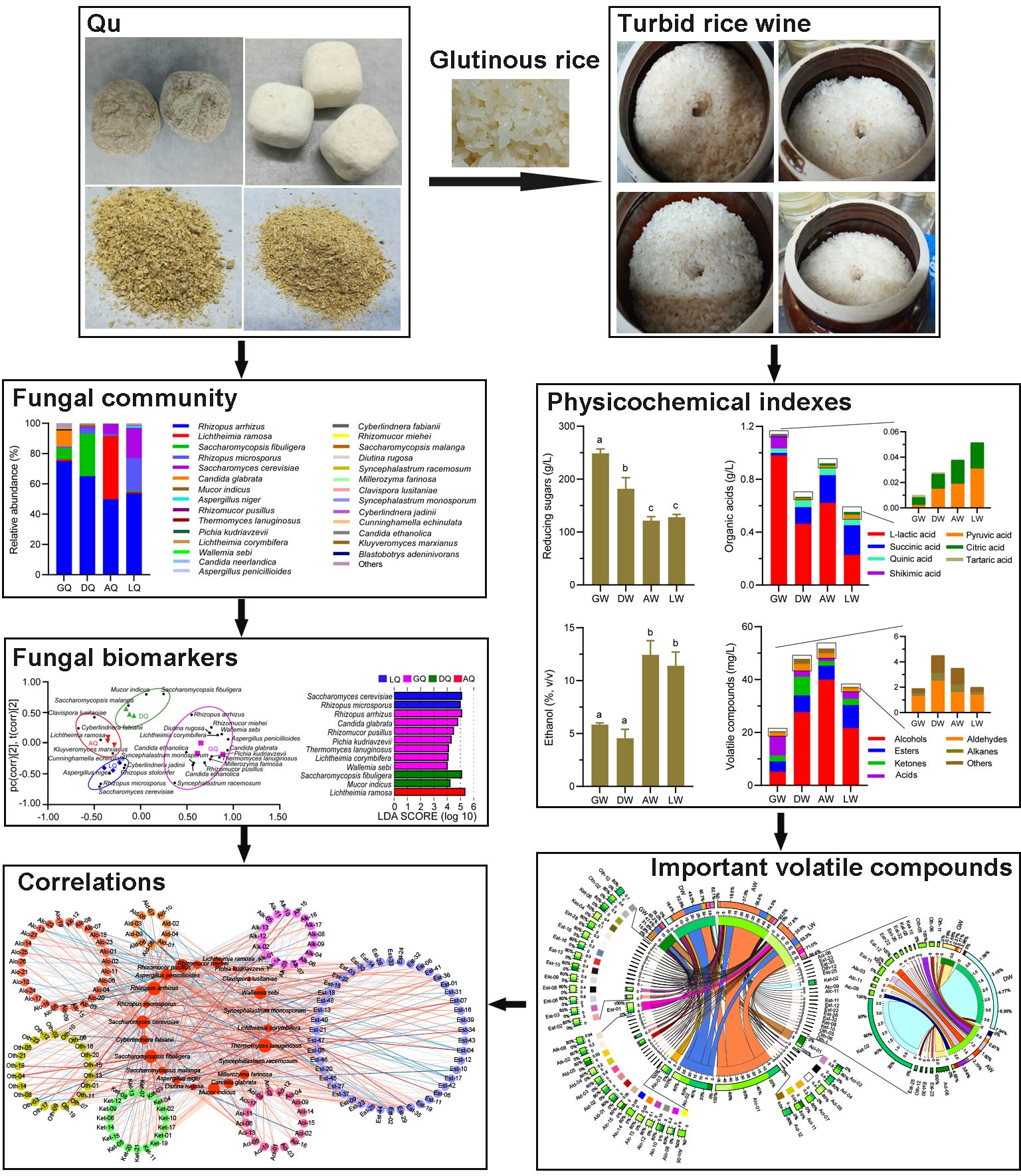
3. Results
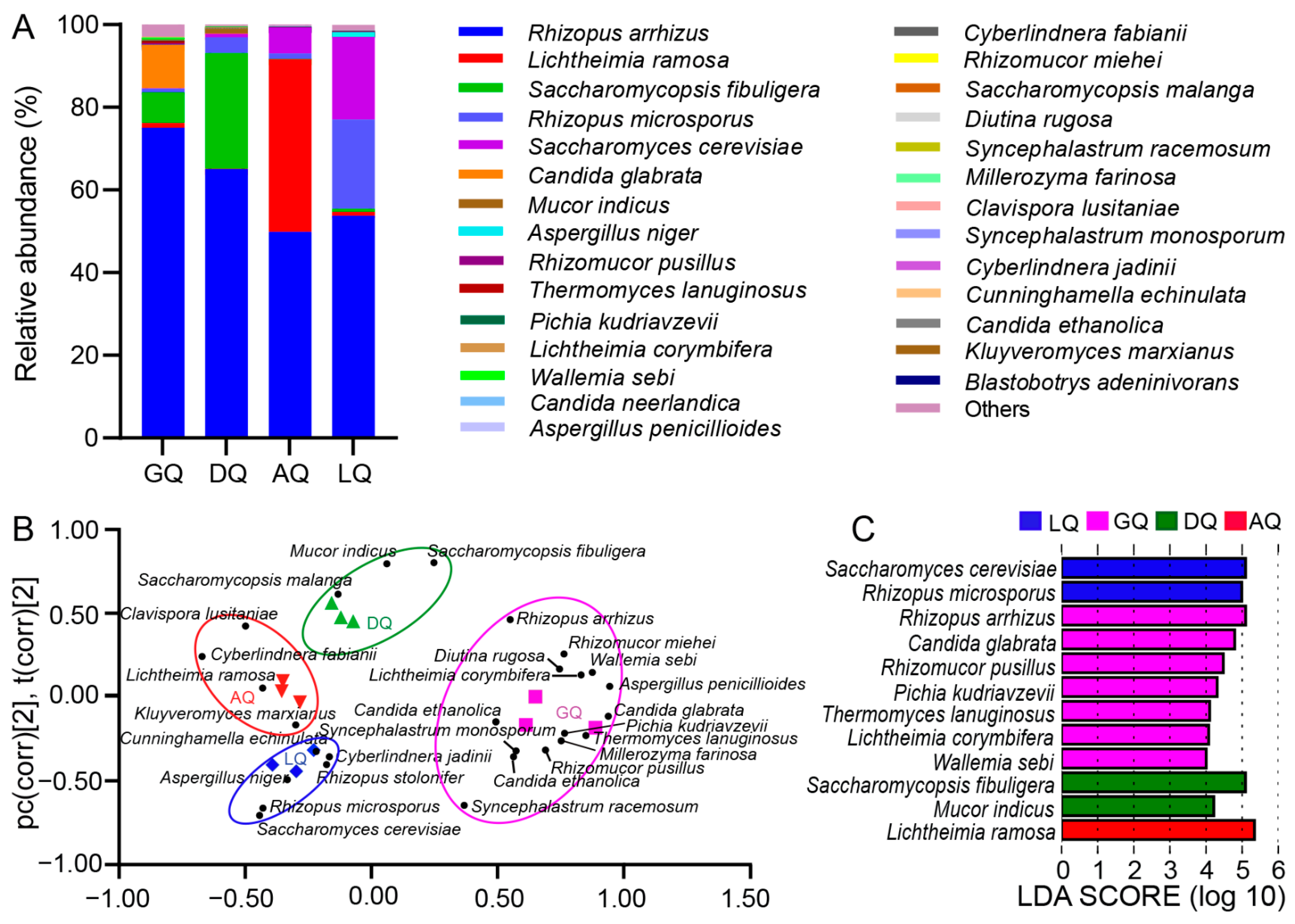
3.1. Fungal Profiles of Different Qu
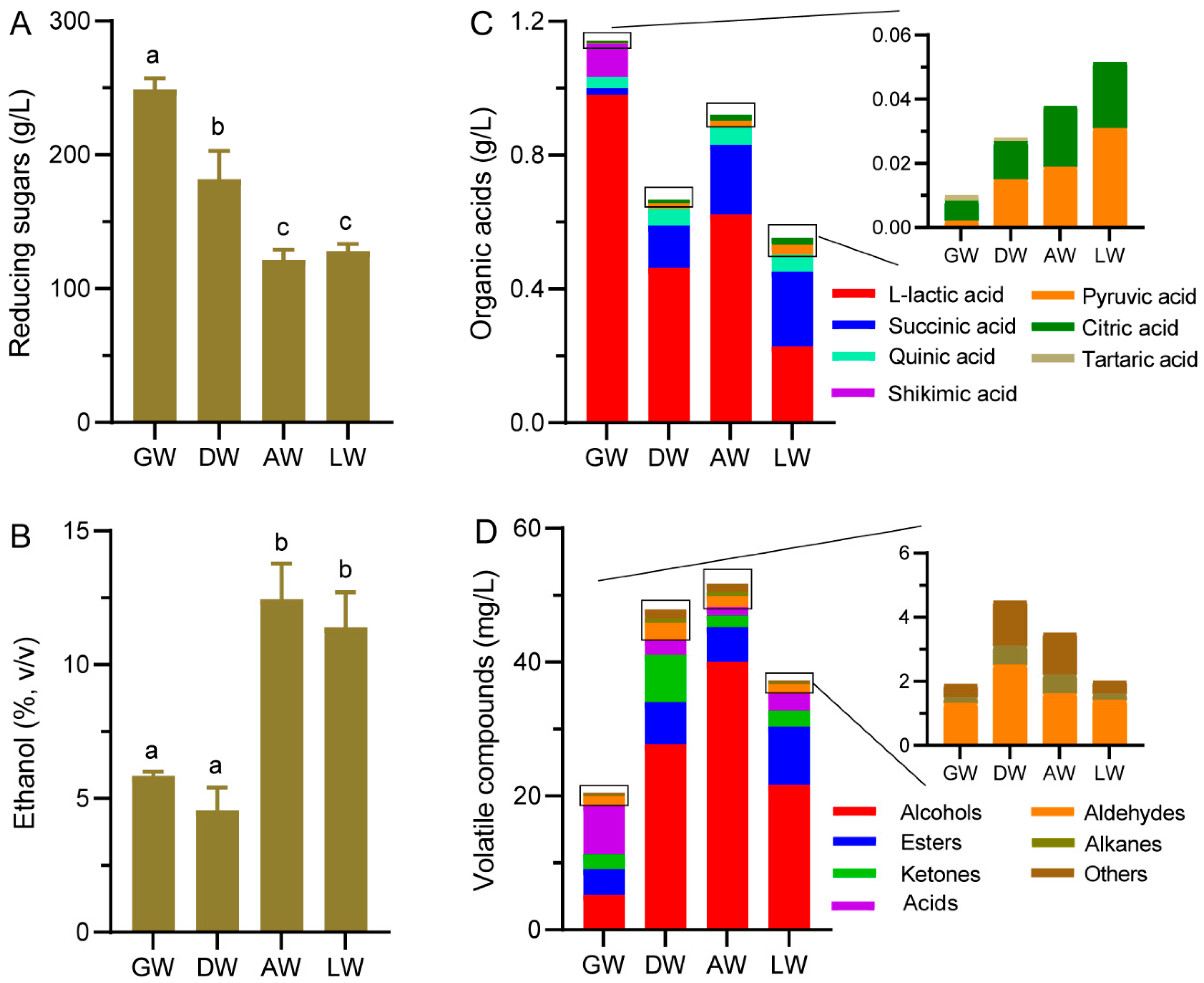
3.2. The Chemical Indexes in TRW
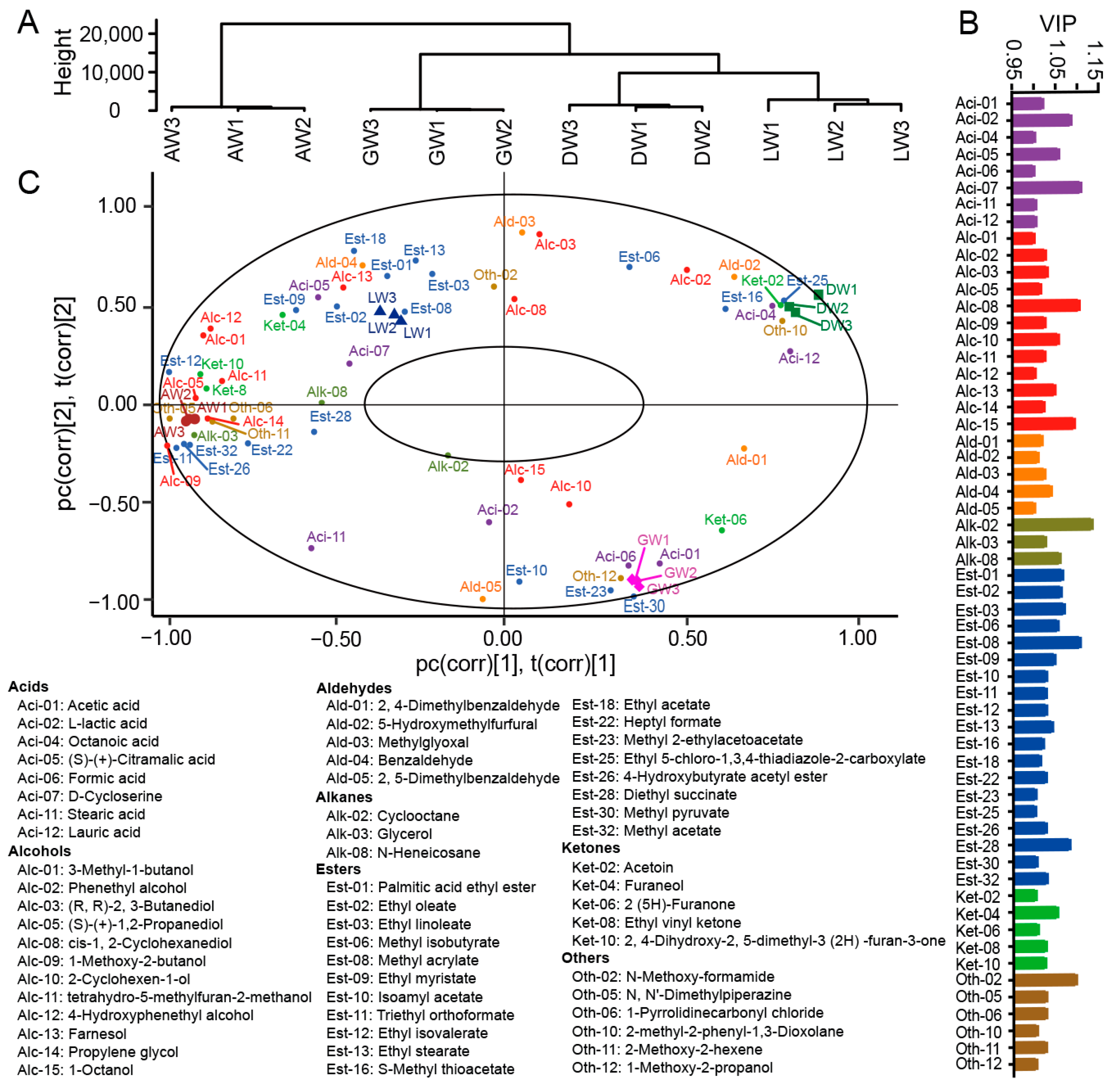
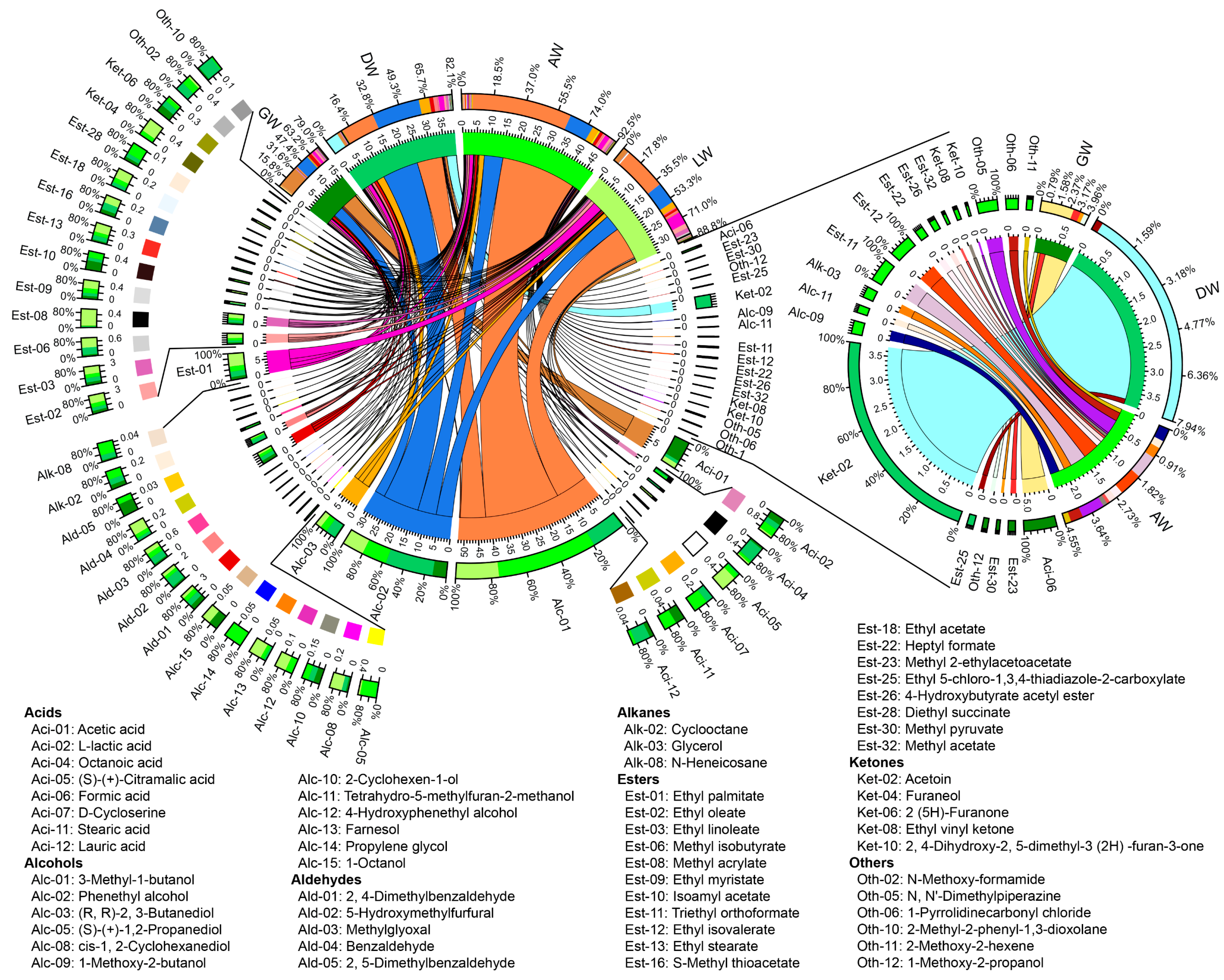
3.3. Volatile Compound Profiles in TRW
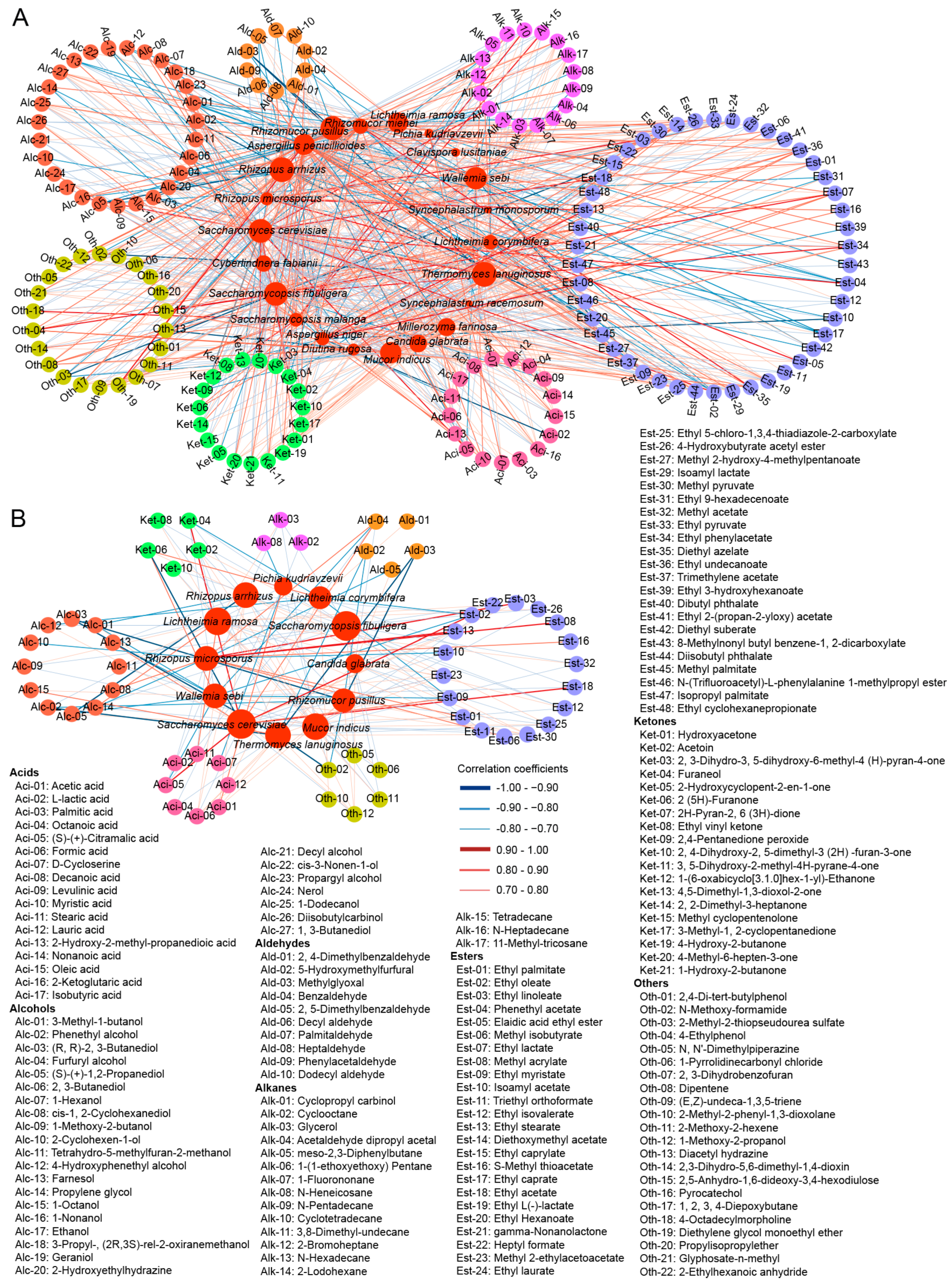
3.4. Correlations between Fungi and Volatile Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, Y.J.; Ai, L.Z.; Mu, Z.Y.; Liu, H.D.; Yan, X.; Ni, L.; Zhang, H.; Xia, Y.J. Flavor compounds with high odor activity values (OAV > 1) dominate the aroma of aged Chinese rice wine (Huangjiu) by molecular association. Food Chem. 2022, 383, 132370. [Google Scholar] [CrossRef]
- Cai, H.Y.; Zhang, T.; Zhang, Q.; Luo, J.; Cai, C.G.; Mao, J.W. Microbial diversity and chemical analysis of the starters used in traditional Chinese sweet rice wine. Food Microbiol. 2018, 73, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.J.; Ren, X.R.; Bian, S.C.; Xu, E.B.; Jin, Z.Y.; Jiao, A.Q. Study on the relationship between the degradation degrees of enzymatically extruded glutinous rice and the qualities of fermented Chinese rice wine. J. Cereal Sci. 2022, 105, 103476. [Google Scholar] [CrossRef]
- Zhu, F.B.; Li, S.; Guan, X.; Huang, K.; Li, Q.Y. Influence of vacuum soaking on the brewing properties of japonica rice and the quality of Chinese rice wine. J. Biosci. Bioeng. 2020, 130, 159–165. [Google Scholar] [CrossRef]
- Xiang, W.L.; Xu, Q.; Zhang, N.D.; Rao, Y.; Zhu, L.; Zhang, Q. Mucor indicus and Rhizopus oryzae co-culture to improve the flavor of Chinese turbid rice wine. J. Sci. Food Agric. 2019, 99, 5577–5585. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Y.; Liang, Z.C.; Lin, X.Z.; He, Z.G.; Ren, X.Y.; Li, W.X.; Molnár, I. Fungal community diversity and fermentation characteristics in regional varieties of traditional fermentation starters for Hong Qu glutinous rice wine. Food Res. Int. 2021, 141, 110146. [Google Scholar] [CrossRef]
- Cao, Y.; Xia, Q.L.; Chen, J.B.; Jin, Z.; Aniya. Understanding of microbial diversity in three representative Qu in China and characterization of the volatile compounds in the corresponding Chinese rice wine. Lwt Food Sci. Technol. 2022, 164, 113680. [Google Scholar] [CrossRef]
- Liu, Z.B.; Wang, Z.Y.; Sun, J.Y.; Ni, L. The dynamics of volatile compounds and their correlation with the microbial succession during the traditional solid-state fermentation of Gutian Hong Qu glutinous rice wine. Food Microbiol. 2020, 86, 103347. [Google Scholar] [CrossRef]
- Zhang, Q.; Sun, Q.; Tan, X.; Zhang, S.M.; Zeng, L.; Tang, J.; Xiang, W.L. Characterization of gamma-aminobutyric acid (GABA)-producing Saccharomyces cerevisiae and coculture with Lactobacillus plantarum for mulberry beverage brewing. J. Biosci. Bioeng. 2020, 129, 447–453. [Google Scholar] [CrossRef]
- Xiang, W.L.; Zhao, Q.H.; Lu, Y.; Tang, J.; Cai, T.; Rao, Y.; Liu, L.; Zhang, Q. Tetracycline residue alters profile of lactic acid bacterial communities and metabolites of ginger pickle during spontaneous fermentation. Food Res. Int. 2022, 155, 111109. [Google Scholar] [CrossRef]
- Chen, L.H.; Li, D.N.; Rong, Y.Z. Fermentation mechanism of ginkgo rice wine using an ultra-high-performance liquid chromatography–quadrupole/time-of-flight mass spectrometry based metabolomics method. J. Food Compos. Anal. 2022, 105, 104230. [Google Scholar] [CrossRef]
- An, X.W.; Wang, Z.J.; Li, J.M.; Nie, X.Y.; Liu, K.X.; Zhang, Y.F.; Zhao, Z.H.; Chitrakar, B.; Ao, C.W. Analysis of flavor-related compounds in fermented persimmon beverages stored at different temperatures. Lwt Food Sci. Technol. 2022, 163, 113524. [Google Scholar] [CrossRef]
- Chen, S.; Xu, Y. The Influence of Yeast Strains on the Volatile Flavour Compounds of Chinese Rice Wine. J. I. Brewing. 2010, 116, 190–196. [Google Scholar] [CrossRef]
- Yang, H.Y.; Peng, Q.; Zhang, H.B.; Sun, J.Q.; Shen, C.; Han, X.Y. The volatile profiles and microbiota structures of the wheat Qus used as traditional fermentation starters of Chinese rice wine from Shaoxing region. Lwt Food Sci. Technol. 2022, 154, 112649. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Y.; Tian, H.X.; Ai, L.Z.; Yu, H.Y. Metagenomic analysis reveals the impact of JIUYAO microbial diversity on fermentation and the volatile profile of Shaoxing-jiu. Food Microbiol. 2020, 86, 103326. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, L.; Zhang, Y.G.; Tu, T.Y.; Wang, S.T.; Shen, C.H.; Yuan, H.W.; Zhong, X.Z. A comparison of microbial communities and volatile compounds in wheat Qu from different geographic locations. Lwt Food Sci. Technol. 2021, 148, 111752. [Google Scholar] [CrossRef]
- Liu, Z.B.; Wang, Z.Y.; Lv, X.C.; Zhu, X.P.; Chen, L.L.; Ni, L. Comparison study of the volatile profiles and microbial communities of Wuyi Qu and Gutian Qu, two major types of traditional fermentation starters of Hong Qu glutinous rice wine. Food Microbiol. 2018, 69, 105–115. [Google Scholar] [CrossRef]
- Huang, Z.R.; Guo, W.L.; Zhou, W.B.; Li, L.; Xu, J.X.; Hong, J.L.; Liu, H.P.; Zeng, F.; Bai, W.D.; Liu, B.; et al. Microbial communities and volatile metabolites in different traditional fermentation starters used for Hong Qu glutinous rice wine. Food Res. Int. 2019, 121, 593–603. [Google Scholar] [CrossRef]
- Lv, X.C.; Chen, Z.C.; Jia, R.B.; Liu, Z.B.; Zhang, W.; Chen, S.J.; Rao, P.F.; Ni, L. Microbial community structure and dynamics during the traditional brewing of Fuzhou Hong Qu glutinous rice wine as determined by culture-dependent and culture-independent techniques. Food Control. 2015, 57, 216–224. [Google Scholar] [CrossRef]
- Lv, X.C.; Jiang, Y.J.; Liu, J.; Guo, W.L.; Liu, Z.B.; Zhang, W.; Rao, P.F.; Ni, L. Evaluation of different PCR primers for denaturing gradient gel electrophoresis (DGGE) analysis of fungal community structure in traditional fermentation starters used for Hong Qu glutinous rice wine. Int. J. Food Microbiol. 2017, 255, 58–65. [Google Scholar] [CrossRef]
- Zhou, Q.F.; Ma, K.; Song, Y.; Wang, Z.W.; Fu, Z.J.; Wang, Y.H.; Zhang, X.Y.; Cui, M.F.; Tang, N.; Xing, X.L. Exploring the diversity of the fungal community in Chinese traditional Baijiu daqu starters made at low-, medium- and high-temperatures. Lwt Food Sci. Technol. 2022, 162, 113408. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Jin, B.; Kelly, J.M. Enhancement of l(+)-lactic acid production using acid-adapted precultures of Rhizopus arrhizus in a bubble column reactor. J. Biosci. Bioeng. 2009, 108, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhang, K.Z.; Cao, X.Z.; Yang, J.G. Effects of Saccharomycopsis fibuligera and Saccharomyces cerevisiae inoculation on small fermentation starters in Sichuan-style Xiaoqu liquor. Food Res. Int. 2020, 137, 109425. [Google Scholar] [CrossRef]
- Song, Y.R.; Lim, B.U.; Baik, S.H. Microbial Diversity and Volatile Flavor Changes during Gayangju Fermentation, a Traditional Korean House Rice Wine. Foods 2022, 11, 2604. [Google Scholar] [CrossRef]
- Larroque, M.N.; Carrau, F.; Farina, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and non-Saccharomyces native yeasts on beer aroma compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef]
- Lakshmi, N.M.; Binoop, M.; Salini, C.N.; Vivek, N.; Sindhu, R.; Pandey, A.; Binod, P. Isobutanol production by Candida glabrata – A potential organism for future fuel demands. Fuel 2021, 306, 121634. [Google Scholar] [CrossRef]
- Garcia, N.F.L.; da Silva Santos, F.R.; Gonçalves, F.A.; da Paz, M.F.; Fonseca, G.G.; Leite, R.S.R. Production of β-glucosidase on solid-state fermentation by Lichtheimia ramosa in agroindustrial residues: Characterization and catalytic properties of the enzymatic extract. Electron. J. Biotechn. 2015, 18, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Garcia, N.F.L.; Santos, F.R.d.S.; Bocchini, D.A.; Paz, M.F.d.; Fonseca, G.G.; Leite, R.S.R. Catalytic properties of cellulases and hemicellulases produced by Lichtheimia ramosa: Potential for sugarcane bagasse saccharification. Ind. Crop Prod. 2018, 122, 49–56. [Google Scholar] [CrossRef] [Green Version]
- de Barros Ranke, F.F.; Shinya, T.Y.; de Figueiredo, F.C.; Fernandez Nunez, E.G.; Cabral, H.; de Oliva Neto, P. Ethanol from rice byproduct using amylases secreted by Rhizopus microsporus var oligosporus. Enzyme partial purification and characterization. J. Environ. Manag. 2020, 266, 110591. [Google Scholar] [CrossRef]
- Liu, A.; Yang, X.; Guo, Q.; Li, B.; Zheng, Y.; Shi, Y.; Zhu, L. Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine. Foods 2022, 11, 1097. [Google Scholar] [CrossRef]
- Qian, Y.L.; An, Y.; Chen, S.; Qian, M.C. Characterization of Qingke Liquor Aroma from Tibet. J. Agric. Food Chem. 2019, 67, 13870–13881. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, Y.; Luo, Q.; Xu, N.; Zhou, M.Z.; Gao, B.; Wang, C.; Shi, Y. Fermenting liquid vinegar with higher taste, flavor and healthy value by using discarded Cordyceps militaris solid culture medium. Lwt Food Sci. Technol. 2018, 98, 654–660. [Google Scholar] [CrossRef]
- Zhao, C.; Su, W.; Mu, Y.C.; Jiang, L.; Mu, Y. Correlations between microbiota with physicochemical properties and volatile flavor components in black glutinous rice wine fermentation. Food Res. Int. 2020, 138, 109800. [Google Scholar] [CrossRef]
- Jin, Z.; Cai, G.L.; Wu, C.; Hu, Z.M.; Xu, X.B.; Xie, G.F.; Wu, D.H.; Lu, J. Profiling the key metabolites produced during the modern brewing process of Chinese rice wine. Food Res. Int. 2021, 139, 109955. [Google Scholar] [CrossRef]
- Salvi, N.A.; Patil, P.N.; Udupa, S.R.; Banerji, A. Biotransformations with Rhizopus arrhizus preparation of the enantiomers of 1-phenylethanol and 1-(fo-, m- and p-methoxyphenyl) ethanols. Tetrahedron Asymmetr. 1995, 6, 2287–2290. [Google Scholar] [CrossRef]
- Han, X.Y.; Qing, X.; Yang, S.Y.; Li, R.L.; Zhan, J.C.; You, Y.L.; Huang, W.D. Study on the diversity of non-Saccharomyces yeasts in Chinese wine regions and their potential in improving wine aroma by beta-glucosidase activity analyses. Food Chem. 2021, 360, 129886. [Google Scholar] [CrossRef]
- Sun, J.Y.; Li, Q.Y.; Luo, S.Q.; Zhang, J.L.; Huang, M.Q.; Chen, F.; Zheng, F.P.; Sun, X.T.; Li, H.H. Characterization of key aroma compounds in Meilanchun sesame flavor style baijiu by application of aroma extract dilution analysis, quantitative measurements, aroma recombination, and omission/addition experiments. RSC Adv. 2018, 8, 23757–23767. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, S.P.; Sun, H.L.; Jiang, Z.F.; Xu, Y.Z.; Mao, J.Q.; Qian, B.; Wang, L.; Mao, J. Metagenomics-based insights into the microbial community profiling and flavor development potentiality of baijiu Daqu and huangjiu wheat Qu. Food Res. Int. 2022, 152, 110707. [Google Scholar] [CrossRef]
- Zhao, Y.Z.; Liu, S.P.; Han, X.; Zhou, Z.L.; Mao, J. Combined effects of fermentation temperature and Saccharomyces cerevisiae strains on free amino acids, flavor substances, and undesirable secondary metabolites in huangjiu fermentation. Food Microbiol. 2022, 108, 104091. [Google Scholar] [CrossRef]
- Martinez-Ruiz, A.; Tovar-Castro, L.; Garcia, H.S.; Saucedo-Castaneda, G.; Favela-Torres, E. Continuous ethyl oleate synthesis by lipases produced by solid-state fermentation by Rhizopus microsporus. Bioresour. Technol. 2018, 265, 52–58. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Xiang, W.; Liang, X.; Liu, J.; Zhu, H.; Cai, T.; Zhang, Q.; Tang, J. Fungal Biomarkers in Traditional Starter Determine the Chemical Characteristics of Turbid Rice Wine from the Rim of the Sichuan Basin, China. Foods 2023, 12, 585. https://doi.org/10.3390/foods12030585
Chen L, Xiang W, Liang X, Liu J, Zhu H, Cai T, Zhang Q, Tang J. Fungal Biomarkers in Traditional Starter Determine the Chemical Characteristics of Turbid Rice Wine from the Rim of the Sichuan Basin, China. Foods. 2023; 12(3):585. https://doi.org/10.3390/foods12030585
Chicago/Turabian StyleChen, Lanchai, Wenliang Xiang, Xuemei Liang, Junyu Liu, Haoyu Zhu, Ting Cai, Qing Zhang, and Jie Tang. 2023. "Fungal Biomarkers in Traditional Starter Determine the Chemical Characteristics of Turbid Rice Wine from the Rim of the Sichuan Basin, China" Foods 12, no. 3: 585. https://doi.org/10.3390/foods12030585
APA StyleChen, L., Xiang, W., Liang, X., Liu, J., Zhu, H., Cai, T., Zhang, Q., & Tang, J. (2023). Fungal Biomarkers in Traditional Starter Determine the Chemical Characteristics of Turbid Rice Wine from the Rim of the Sichuan Basin, China. Foods, 12(3), 585. https://doi.org/10.3390/foods12030585