
Using Front-Face Fluorescence Spectroscopy and Biochemical Analysis of Honey to Assess a Marker for the Level of Varroa destructor Infestation of Honey Bee (Apis mellifera) Colonies

 ,
,  , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents and Solutions
2.2. Samples
2.3. Methods
2.3.1. Determination of V. destructor Infestation Rates
2.3.2. Melissopalynological Analysis
2.3.3. Determination of the Catalase Activity in Honey Samples
2.3.4. Determination of Diastase Activity in Honey Samples
2.3.5. Determination of Total Phenolic Content (TotPhC)
2.3.6. Determination of Total Protein Content (TotPrC)
2.3.7. Fluorescence Spectroscopy
2.4. Data Analysis
2.4.1. Data Analysis and Modelling of Spectral Features
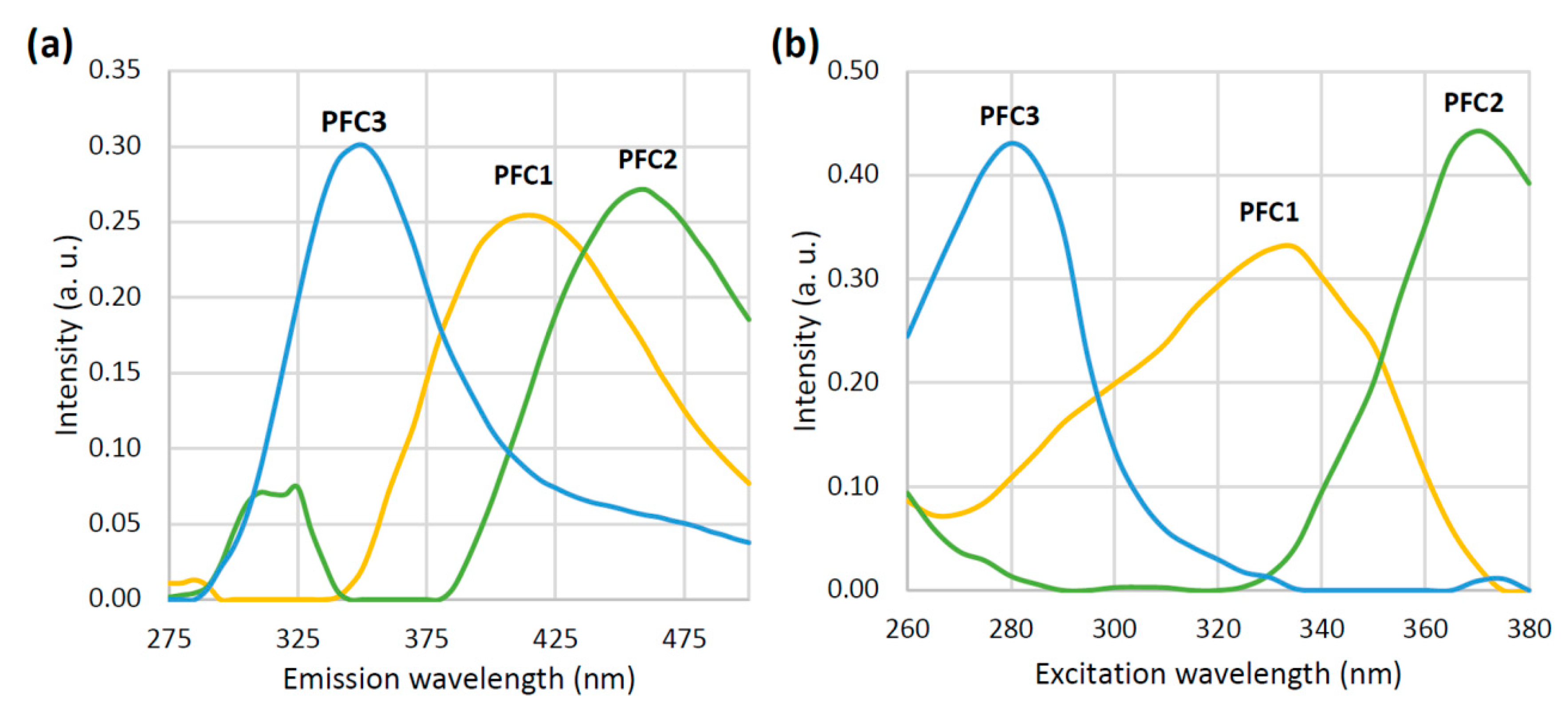
2.4.2. PARAFAC
2.4.3. Correlation Analysis
3. Results
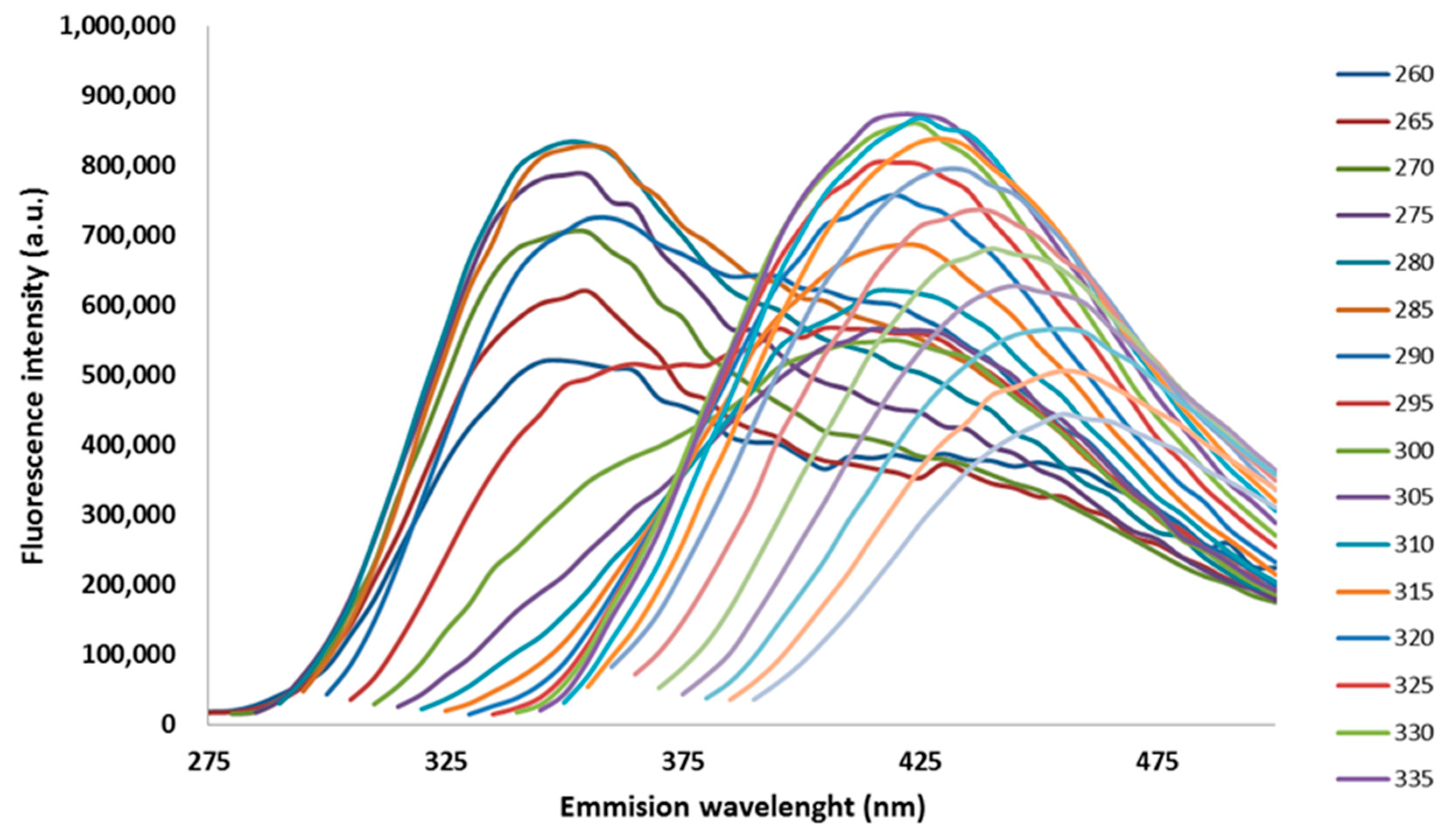
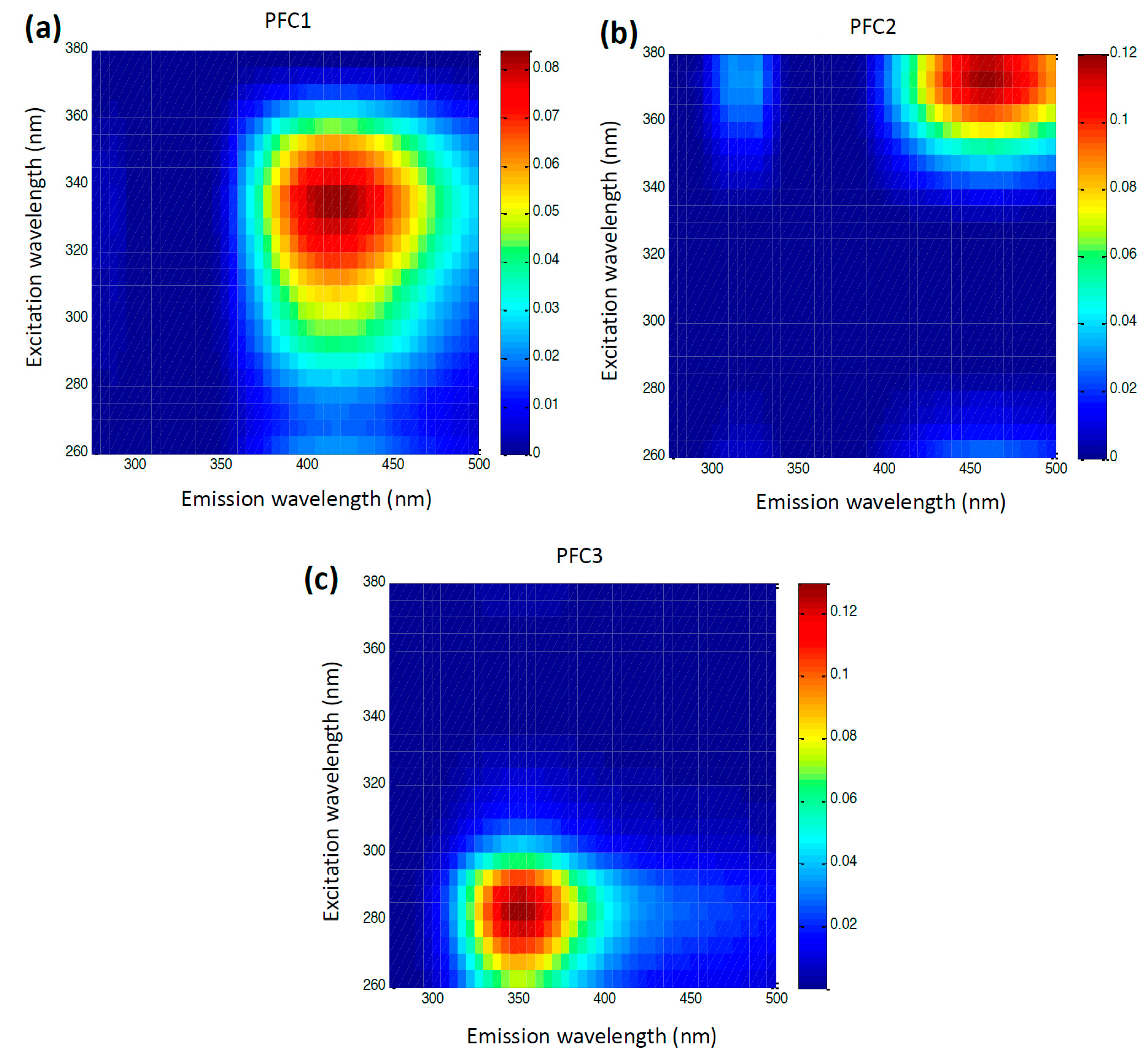
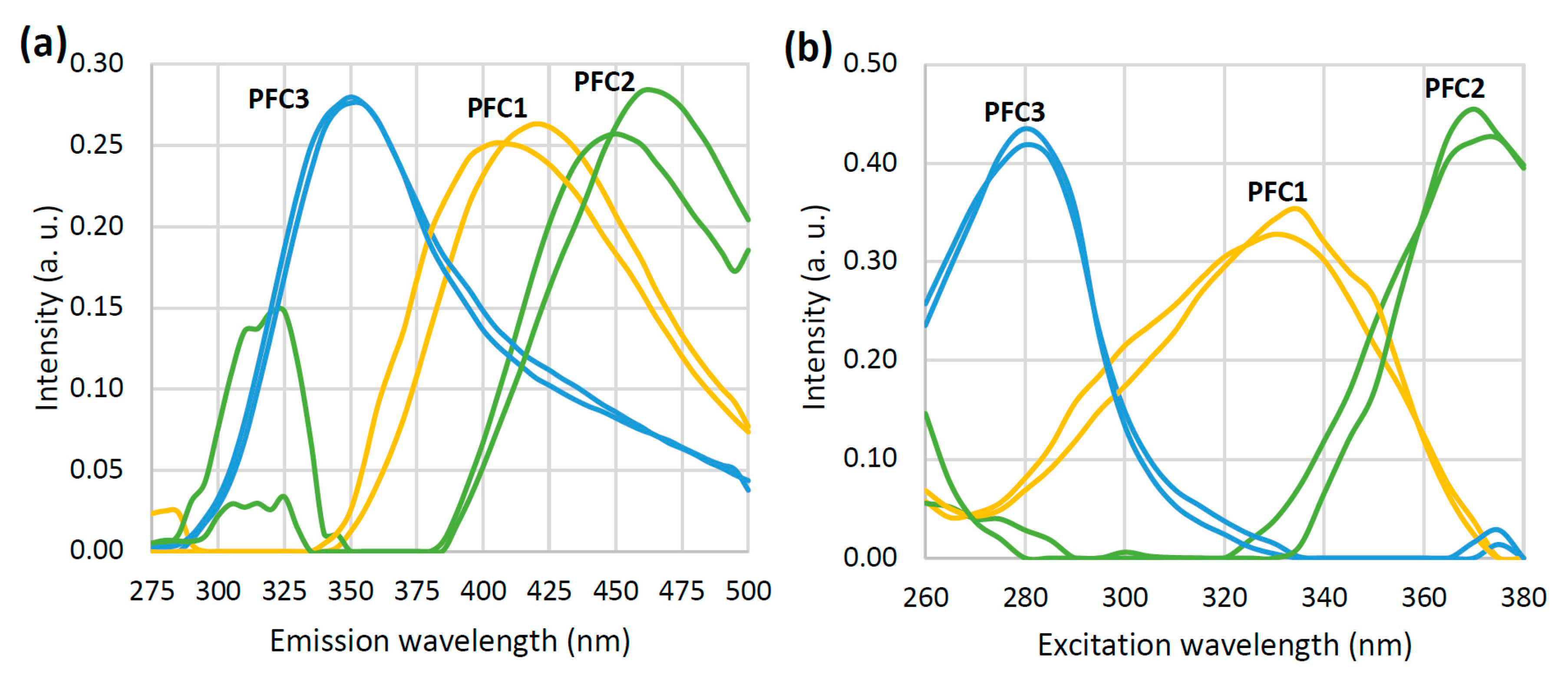
3.1. Emission Spectra and PARAFAC
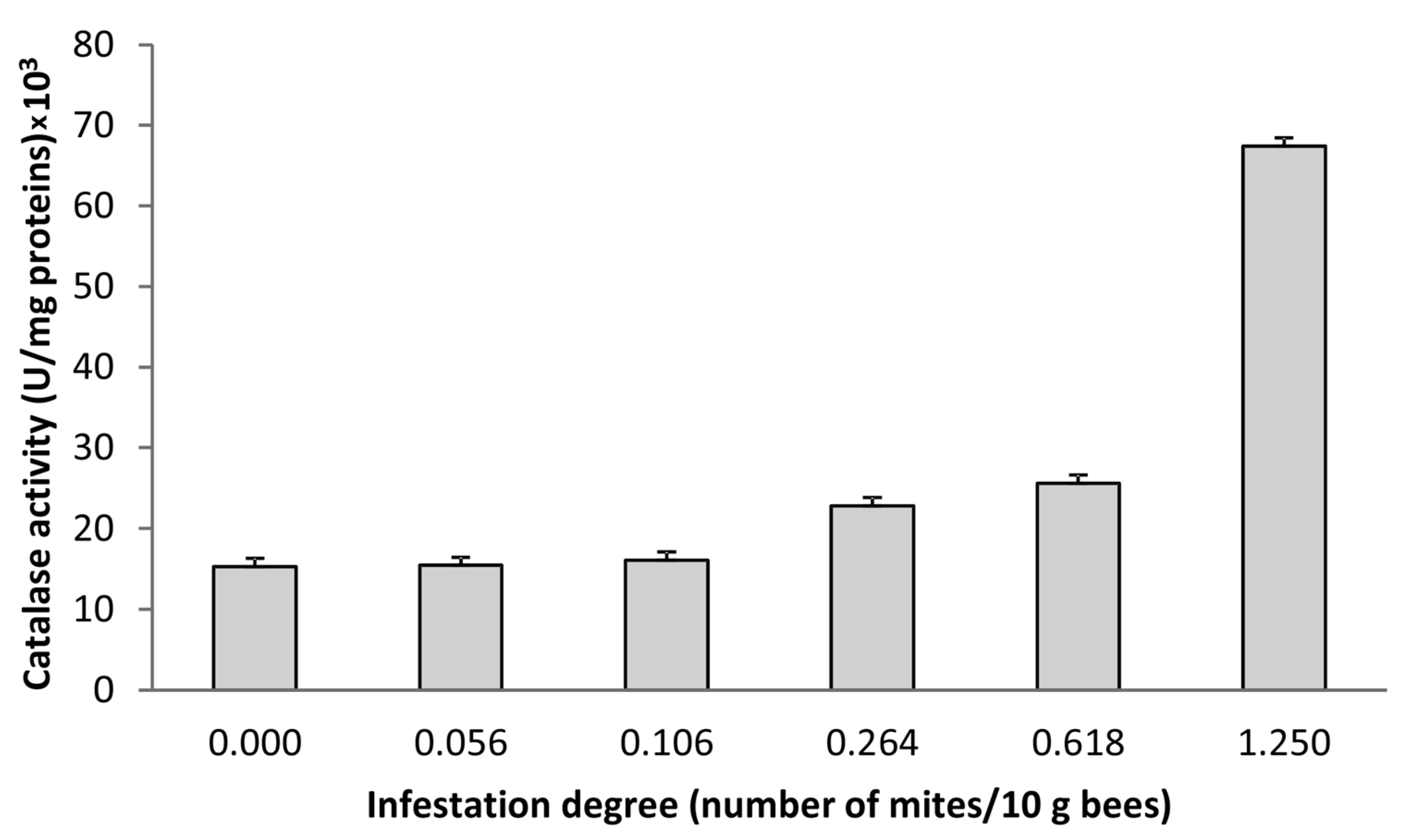
3.2. Correlation Analysis of Spectral, Biochemical, and Melissopalynological Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fakhlaei, R.; Selamat, J.; Khatib, A.; Razis, A.F.A.; Sukor, R.; Ahmad, S.; Babadi, A.A. The Toxic Impact of Honey Adulteration: A Review. Foods 2020, 9, 1538. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Currie, R.W.; Pernal, S.F.; Guzman-Novoa, E. Honey bee colony losses in Canada. J. Apic. Res. 2010, 49, 104–106. [Google Scholar] [CrossRef]
- Neumann, P.; Carreck, N.L. Honey bee colony losses. J. Apic. Res. 2010, 49, 1–6. [Google Scholar] [CrossRef]
- Colin, T.; Lim, M.Y.; Quarrell, S.R.; Allen, G.R.; Barron, A.B. Effects of thymol on European honey bee hygienic behaviour. Apidologie 2019, 50, 141–152. [Google Scholar] [CrossRef]
- Van Engelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar] [CrossRef]
- Emsen, B.; Guzman-Novoa, E.; Kelly, P.G. Honey production of honey bee (Hymenoptera: Apidae) colonies with high and low Varroa destructor (Acari: Varroidae) infestation rates in eastern Canada. Entomol. Soc. Can. 2013, 146, 236–240. [Google Scholar] [CrossRef]
- Revainera, P.D.; Quintana, S.; Fernandez de Landa, G.; García Iza, C.; Olivera, E.; Fuentes, G.; Plischuk, S.; Medici, S.; Ruffinengo, S.; Marcangelli, J.; et al. Molecular detection of bee pathogens in honey. J. Insects Food Feed. 2020, 6, 467–474. [Google Scholar] [CrossRef]
- Branko, M.R.; Kidd, N.A.C.; Pickard, R.S. A comparative evaluation of sampling methods for Varroa destructor (Acari: Varroidae) population estimation. Apidologie 2006, 37, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Pirk, C.W.W.; de Miranda, J.R.; Fries, I.; Kramer, M.; Paxton, R.; Murray, T.; Nazzi, F.; Shutler, D.; Van Der Steen, J.J.M.; Van Dooremalen, C. Standard methods for Apis mellifera research. J. Apic. Res. 2013, 52, 1–56. [Google Scholar] [CrossRef]
- Evans, J.D.; Schwarz, R.S.; Chen, Y.P.; Budge, G.; Cornman, R.S.; De La Rua, P.; de Miranda, J.R.; Foret, S.; Foster, L.; Gauthier, L.; et al. Standard methods for molecular research in Apis mellifera. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef]
- Chua, L.S.; Lee, J.Y.; Chan, G.F. Characterization of the Proteins in Honey. Anal. Lett. 2015, 48, 697–709. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Pia Manna, P.; Zhang, J.; Bravo Lamas, L.; Martínez Flórez, S.; Agudo Toyos, P.; et al. Phenolic Compounds in Honey and Their Associated Health Benefits: A Review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef]
- Stanković, M.; Bartolić, D.; Šikoparija, B.; Spasojević, D.; Mutavdžić, D.; Natić, M.; Radotić, K. Variability estimation of the protein and phenol total content in honey using front face fluorescence spectroscopy coupled with MCR–ALS analysis. J. Appl. Spectrosc. 2019, 86, 256–263. [Google Scholar] [CrossRef]
- Nazarian, H.; Taghavizad, R.; Majd, A. Origin of honey proteins and method for its quality control. Pak. J. Bot. 2010, 42, 3221–3228. [Google Scholar]
- Sarmento da Silva, T.M.; dos Santos, F.P.; Evangelista-Rodrigues, A.; Sarmento da Silva, E.M.; Sarmento da Silva, G.; Santos de Novais, J.; dos Santos, F.A.R.; Camara, C.A. Phenolic compounds, melissopalynological, physicochemical analysis and antioxidant activity of jandaíra (Melipona subnitida) honey. J. Food Compost. Anal. 2013, 29, 10–18. [Google Scholar] [CrossRef]
- Stanković, M.; Bartolić, D.; Mutavdžić, D.; Marković, S.; Grubić, S.; Jovanović, N.M.; Radotić, K. Estimation of honey bee colony infection with Nosema ceranae and Varroa destructor using fluorescence spectroscopy in combination with differential scanning calorimetry of honey samples. J. Apic. Res. 2021, 1–7. [Google Scholar] [CrossRef]
- Negri, P.; Villalobos, E.; Szawarski, N.; Damiani, N.; Gende, L.; Garrido, M.; Maggi, M.; Quintana, S.; Lamattina, L.; Eguaras, M. Towards Precision Nutrition: A Novel Concept Linking Phytochemicals, Immune Response and Honey Bee Health. Insects 2019, 10, 401. [Google Scholar] [CrossRef]
- Wegener, J.; Ruhnke, H.; Scheller, K.; Mispagel, S.; Knollmann, U.; Kamp, G.; Bienefeld, K. Pathogenesis of varroosis at the level of the honey bee (Apis mellifera) colony. J. Insect Physiol. 2016, 91, 1–9. [Google Scholar] [CrossRef]
- Gillette, C. Honey catalase. J. Econ. Entomol. 1931, 24, 605–606. [Google Scholar] [CrossRef]
- Oddo, L.P.; Baldi, E.; Accorti, M. Diastatic activity in some unifloral honeys. Apidologie 1990, 21, 17–24. [Google Scholar] [CrossRef]
- Lenhardt, L.; Bro, R.; Zeković, I.; Dramićanin, T.; Dramićanin, M.D. Fluorescence spectroscopy coupled with PARAFAC and PLS DA for characterization and classification of honey. Food Chem. 2015, 175, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Bañegila, M.; Martín-Vertedorb, D.; Bosellic, E.; Durán-Merása, I. Control of olive cultivar irrigation by front-face fluorescence excitation-emission matrices in combination with PARAFAC. J. Food Compost. Anal. 2018, 69, 189–196. [Google Scholar] [CrossRef]
- Bartolić, D.; Mojović, M.; Prokopijević, M.; Djikanović, D.; Kalauzi, A.; Mutavdžić, D.; Baošić, R.; Radotić. K. Lignin and organic free radicals in maize (Zea mays L.) seeds in response to aflatoxin B1 contamination: An optical and EPR spectroscopic study. J. Sci. Food Agric. 2022, 102, 2500–2505. [Google Scholar] [CrossRef]
- Bro, R.; Kiers, H.A.L. A new efficient method for determining the number of components in PARAFAC models. J. Chemom. 2003, 17, 274–286. [Google Scholar] [CrossRef]
- Lemanski, N.J.; Cook, C.N.; Smith, B.H.; Pinter-Wollman, N. A Multiscale Review of Behavioral Variation in Collective Foraging Behavior in Honey Bees. Insects 2019, 10, 370. [Google Scholar] [CrossRef]
- Klein, S.; Pasquaretta, C.; Jiang He, X.; Perry, C.; Søvik, E.; Marc Devaud, J.; Barron, A.B.; Lihoreau, M. Honey bees increase their foraging performance and frequency of pollen trips through experience. Sci. Rep. 2019, 9, 6778. [Google Scholar] [CrossRef]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard methods for Varroa research.Vol. 2: Standard methods for Apis mellifera research. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef]
- Uzunov, A.; Büchler, R.; Bienefeld, K. Performance Testing Protocol. A Guide for European Honey Bee Breeders, 1st ed.; Landesbetrieb Landwirtschaft Hessen: Kirchhain, Germany, 2015. [Google Scholar]
- Von der Ohe, W.; Persano Odo, L.; Piana, M.L.; Morlot, M.; Martin, P. Harmonized methods of melissopalynology. Apidologie 2004, 35, 18–25. [Google Scholar] [CrossRef]
- Moore, P.D.; Webb, J.A. An Illustrated Guide to Pollen Analysis; Hodder & Stoughton Educational Division: London, UK, 1978; 133p. [Google Scholar]
- Bucher, E.; Kofler, V.; Vorwohl, G.; Zieger, E. Das Pollenbild der SudtirolerHonige; Biologisches Labor der Landesagentur fur Umwelt und Arbeitsschutz: Bolzano, Italy, 2004; pp. 17–37. [Google Scholar]
- Riccardelli D'Albore, G. Mediterranean Melissopalynology; Università degli Studi di Perugia: Perugia, Italy, 1998; 466p. [Google Scholar]
- Behm, F.; Von der Ohe, K.; Henrich, W. Reliability of pollen analysis in honey. Dtsch. Lebensm.-Rundsch. 1996, 92, 183–187. [Google Scholar]
- Louveaux, J.; Maurizio, A.; Vorwohl, G. Methods of Melissopalynology. Bee World 1978, 59, 139–157. [Google Scholar] [CrossRef]
- Huidobro, J.F.; Sánchez, M.P.; Muniategui, S.; Sancho, M.T. Precise method for the measurement of catalase activity in honey. J. AOAC Int. 2005, 88, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Schade, J.E.; Marsh, G.L.; Eckert, J.E. Diastatic activity and hydroxymethylfurfural in honey and their usefulness in detecting heat adulteration. Food Res. 1958, 23, 446–463. [Google Scholar] [CrossRef]
- International Honey Commission. Harmonised Methods of the International Honey Commission. Available online: https://www.ihc-platform.net/ihcmethods2009.pdf (accessed on 10 February 2021).
- Gašić, U.; Kečkeš, S.; Dabić, D.; Trifković, J.; Milojković-Opsenica, D.; Natić, M.; Tešić, Ž. Phenolic profile and antioxidant activity of Serbian polyfloral honeys. Food Chem. 2014, 145, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagent. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. J. Food Sci. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bartolić, D.; Stanković, M.; Mutavdžić, D.; Stanković, S.; Jovanović, D.; Radotić, K. Multivariate Curve Resolution-Alternate Least Square Analysis of Excitation-Emission Matrices for Maize Flour Contaminated with Aflatoxin B1. J. Fluoresc. 2018, 28, 729–733. [Google Scholar] [CrossRef]
- Stanković, M.; Nikčević, M.; Radotić, K. Annual variation of proteins and phenols in honey of a bee society using fluorescence spectroscopy: A way to assess effects of antivarroa treatments on honey composition. Eur. Food Res. Technol. 2020, 246, 1515–1518. [Google Scholar] [CrossRef]
- Bro, R. PARAFAC. Tutorial and applications. Chemom. Intell. Lab. Syst. 1997, 38, 149–171. [Google Scholar] [CrossRef]
- Rodriguez Delgado, M.A.; Malovana, S.; Perez, J.P.; Borges, T.; Garcia Montelongo, F.J. Separation of phenolic compounds by high performance liquid chromatography with absorbance and fluorometric detection. J. Chromatogr. A 2001, 912, 249–257. [Google Scholar] [CrossRef]
- Lang, M.; Stober, F.; Lichtenthaler, H.K. Fluorescence emission spectra of plant leaves and plant constituents. Radiat. Environ. Biophys. 1991, 30, 333–347. [Google Scholar] [CrossRef]
- Radotić, K.; Kalauzi, A.; Djikanović, D.; Jeremić, M.; Leblanc, R.M.; Cerovic, Z.G. Component analysis of the fluorescence spectra of a lignin model compound. J. Photochem. Photobiol. B 2006, 83, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Karoui, R.; Dufour, E.; Bosset, J.O.; De Baerdemaeker, J. The use of front face fluorescence spectroscopy to classify the botanical origin of honey samples produced in Switzerland. Food Chem. 2007, 101, 314–323. [Google Scholar] [CrossRef]
- Dustman, J.H. UÈber die KatalaseaktivitaÈt in Bienenhonig aus der Tracht der HeidekrautgewaÈchse (Ericaceae). Z. Lebensm. Unters. Forsch. 1971, 145, 292–295. [Google Scholar] [CrossRef]
- Zamocky, M.; Furtmüller, P.G.; Obinger, C. Evolution of Catalases from Bacteria to Humans. Antioxid. Redox Signal. 2008, 10, 1527–1548. [Google Scholar] [CrossRef] [PubMed]
- Badotra, P.; Kumar, N.R.; Harjai, K. Varroa causes oxidative stress in Apis mellifera L. J. Glob. Biosci. 2013, 2, 199–201. [Google Scholar]
- Dmochowska-Ślęzak, K.; Dmitryjuk, M.; Zaobidna, E.; Żółtowska, K. The antioxidant defense system of Varroa destructor mites facilitates the infestation of Apis mellifera. J. Apic. Res. 2016, 60, 163–165. [Google Scholar] [CrossRef]
- Ferreres, F.; Andrade, P.; Gil, M.I.; Tomas-Barberan, F.A. Floral nectar phenolics as biochemical markers for the botanical origin of heather honey. Z. Lebensm. Unters. Forsch. 1996, 202, 40–44. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infestation | Catalase | Diastase | TotPrC | TotPhC | Pro/Phe | Phe/Pro | PFC1 | PFC2 | PFC3 | PFC1/PFC3 | PFC3/PFC1 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Infestation | 1.000 | 0.936 | 0.406 | −0.047 | −0.108 | 0.003 | −0.036 | 0.656 | 0.513 | 0.608 | 0.061 | −0.184 |
| Catalase | 1.000 | 0.308 | 0.160 | 0.049 | 0.104 | −0.154 | 0.753 | 0.672 | 0.662 | 0.223 | −0.272 | |
| Diastase | 1.000 | 0.160 | 0.499 | −0.271 | 0.298 | 0.706 | 0.606 | 0.700 | −0.035 | −0.080 | ||
| TotPrC | 1.000 | 0.653 | 0.648 | −0.583 | 0.476 | 0.563 | 0.452 | 0.129 | −0.066 | |||
| TotPhC | 1.000 | −0.149 | 0.221 | 0.590 | 0.728 | 0.414 | 0.466 | −0.425 | ||||
| Pro/Phe | 1.000 | −0.984 | 0.003 | −0.035 | 0.180 | −0.377 | 0.416 | |||||
| Phe/Pro | 1.000 | 0.004 | 0.040 | −0.181 | 0.376 | −0.440 | ||||||
| PFC1 | 1.000 | 0.938 | 0.907 | 0.276 | −0.310 | |||||||
| PFC2 | 1.000 | 0.753 | 0.521 | −0.486 | ||||||||
| PFC3 | 1.000 | −0.131 | 0.115 | |||||||||
| PFC1/PFC3 | 1.000 | −0.941 | ||||||||||
| PFC3/PFC1 | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanković, M.; Prokopijević, M.; Šikoparija, B.; Nedić, N.; Andrić, F.; Polović, N.; Natić, M.; Radotić, K. Using Front-Face Fluorescence Spectroscopy and Biochemical Analysis of Honey to Assess a Marker for the Level of Varroa destructor Infestation of Honey Bee (Apis mellifera) Colonies. Foods 2023, 12, 629. https://doi.org/10.3390/foods12030629
Stanković M, Prokopijević M, Šikoparija B, Nedić N, Andrić F, Polović N, Natić M, Radotić K. Using Front-Face Fluorescence Spectroscopy and Biochemical Analysis of Honey to Assess a Marker for the Level of Varroa destructor Infestation of Honey Bee (Apis mellifera) Colonies. Foods. 2023; 12(3):629. https://doi.org/10.3390/foods12030629
Chicago/Turabian StyleStanković, Mira, Miloš Prokopijević, Branko Šikoparija, Nebojša Nedić, Filip Andrić, Natalija Polović, Maja Natić, and Ksenija Radotić. 2023. "Using Front-Face Fluorescence Spectroscopy and Biochemical Analysis of Honey to Assess a Marker for the Level of Varroa destructor Infestation of Honey Bee (Apis mellifera) Colonies" Foods 12, no. 3: 629. https://doi.org/10.3390/foods12030629
APA StyleStanković, M., Prokopijević, M., Šikoparija, B., Nedić, N., Andrić, F., Polović, N., Natić, M., & Radotić, K. (2023). Using Front-Face Fluorescence Spectroscopy and Biochemical Analysis of Honey to Assess a Marker for the Level of Varroa destructor Infestation of Honey Bee (Apis mellifera) Colonies. Foods, 12(3), 629. https://doi.org/10.3390/foods12030629