
α-Linolenic Acid Inhibits RANKL-Induced Osteoclastogenesis In Vitro and Prevents Inflammation In Vivo

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Cell Induction
2.2. Preparation of Fatty Acids
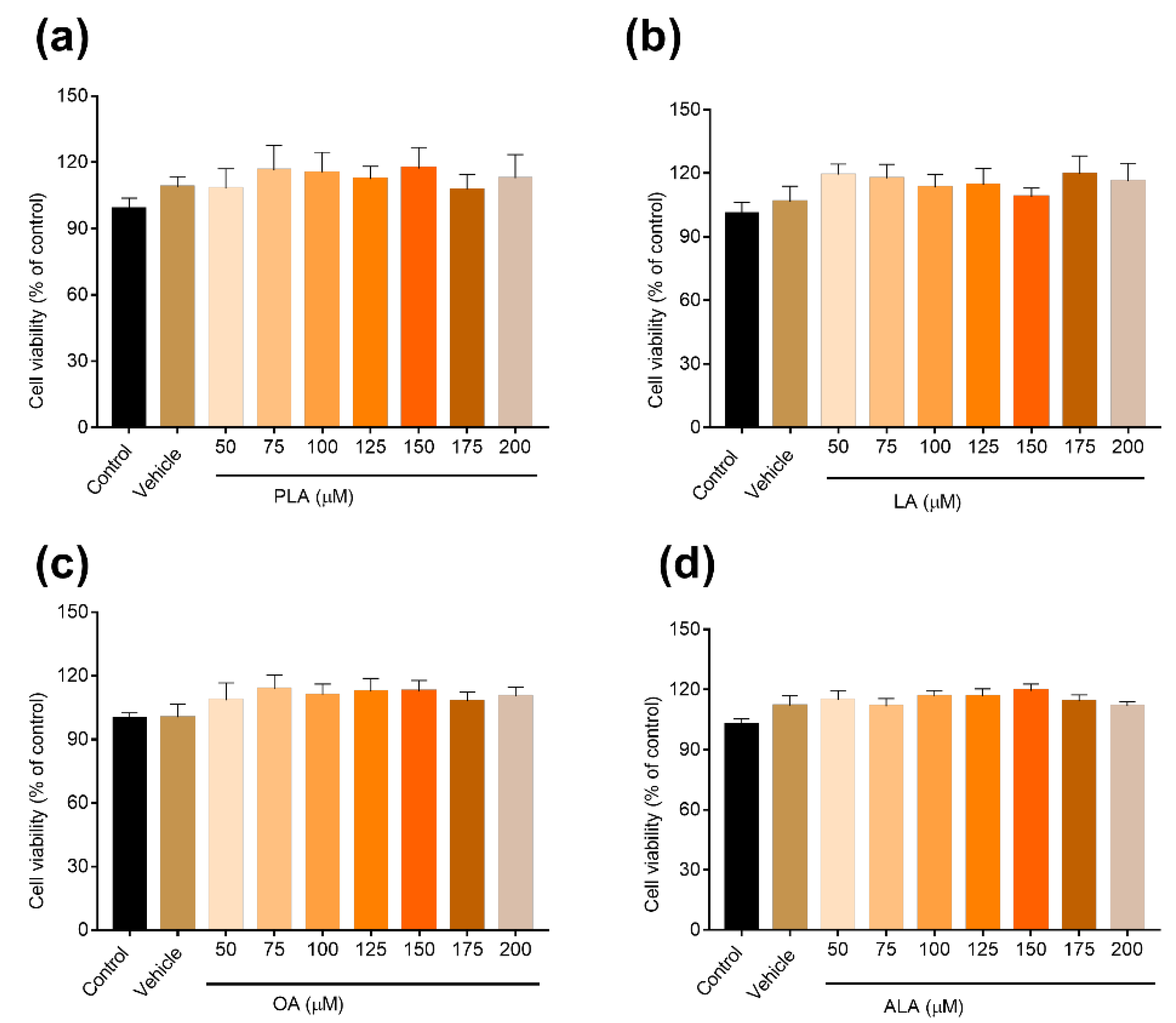
2.3. Cell Viability Assays
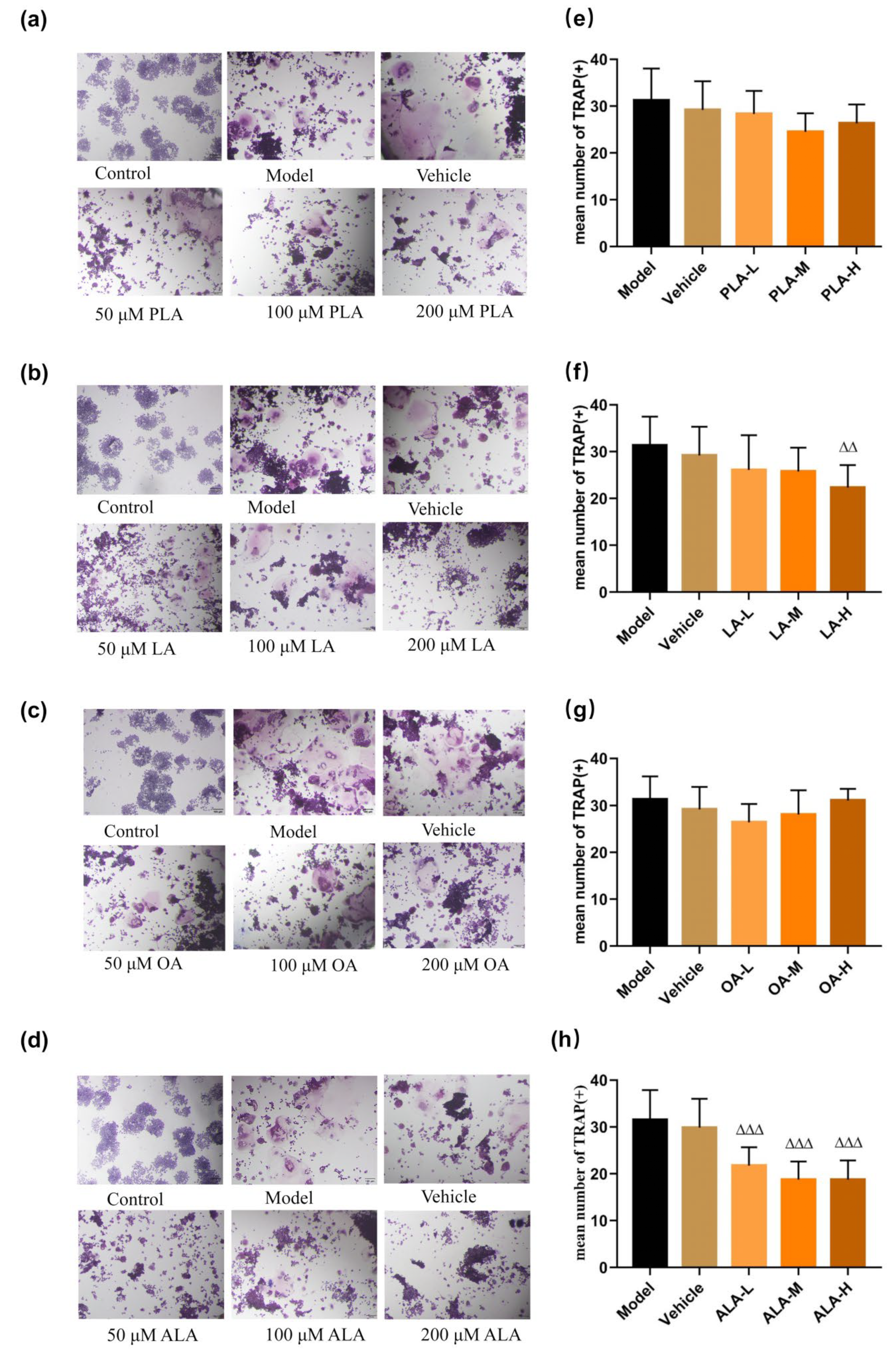
2.4. TRAP Staining
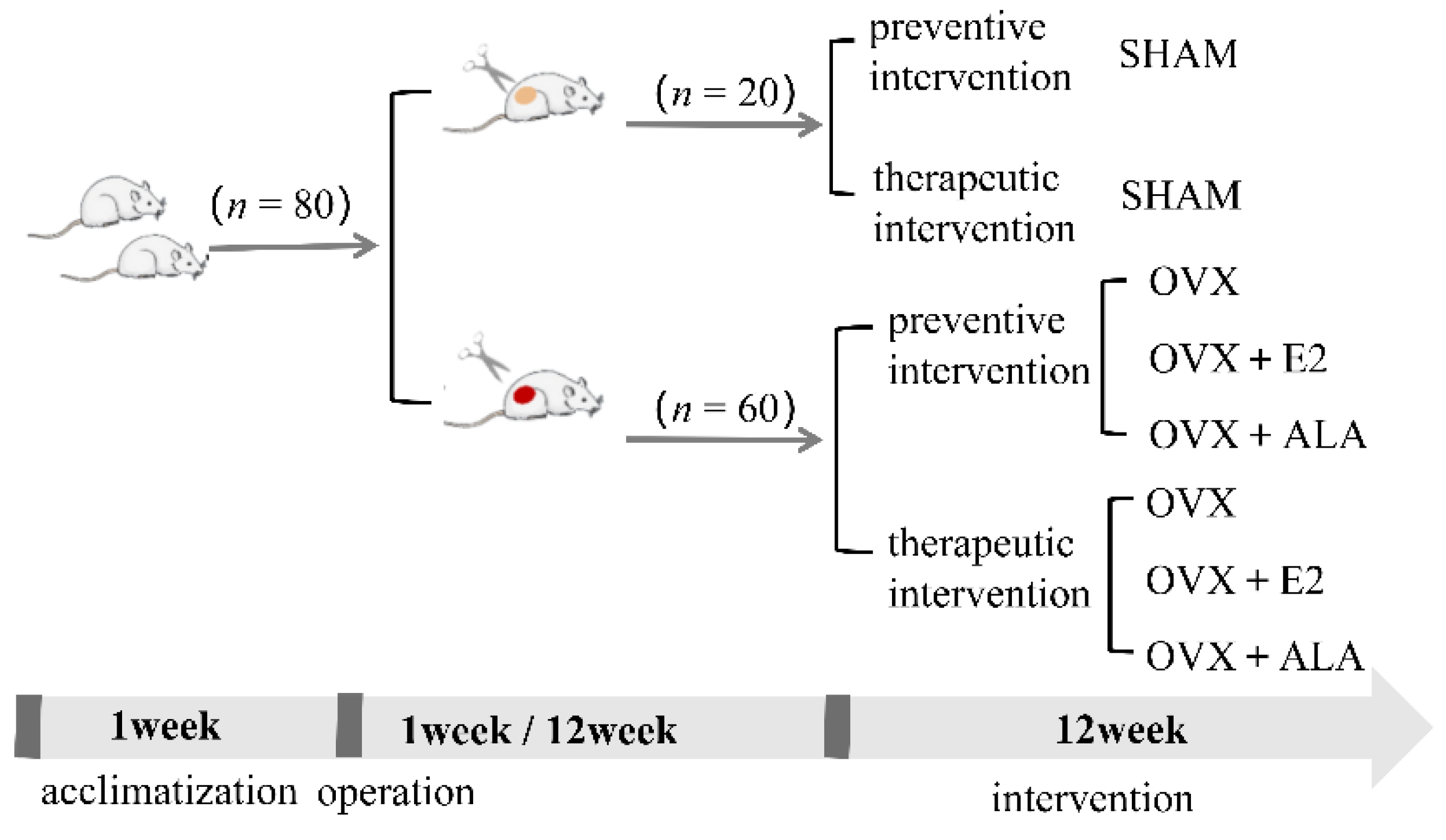
2.5. Animals and Experimental Design
2.6. Micro-CT
2.7. Blood Routine Examination and Serum Inflammatory Factor Analysis
2.8. RNA Isolation and Real-Time Quantitative
2.9. Statistical Analyses
3. Results
3.1. Fatty Acids Inhibited RANKL-Induced Osteoclastogenesis
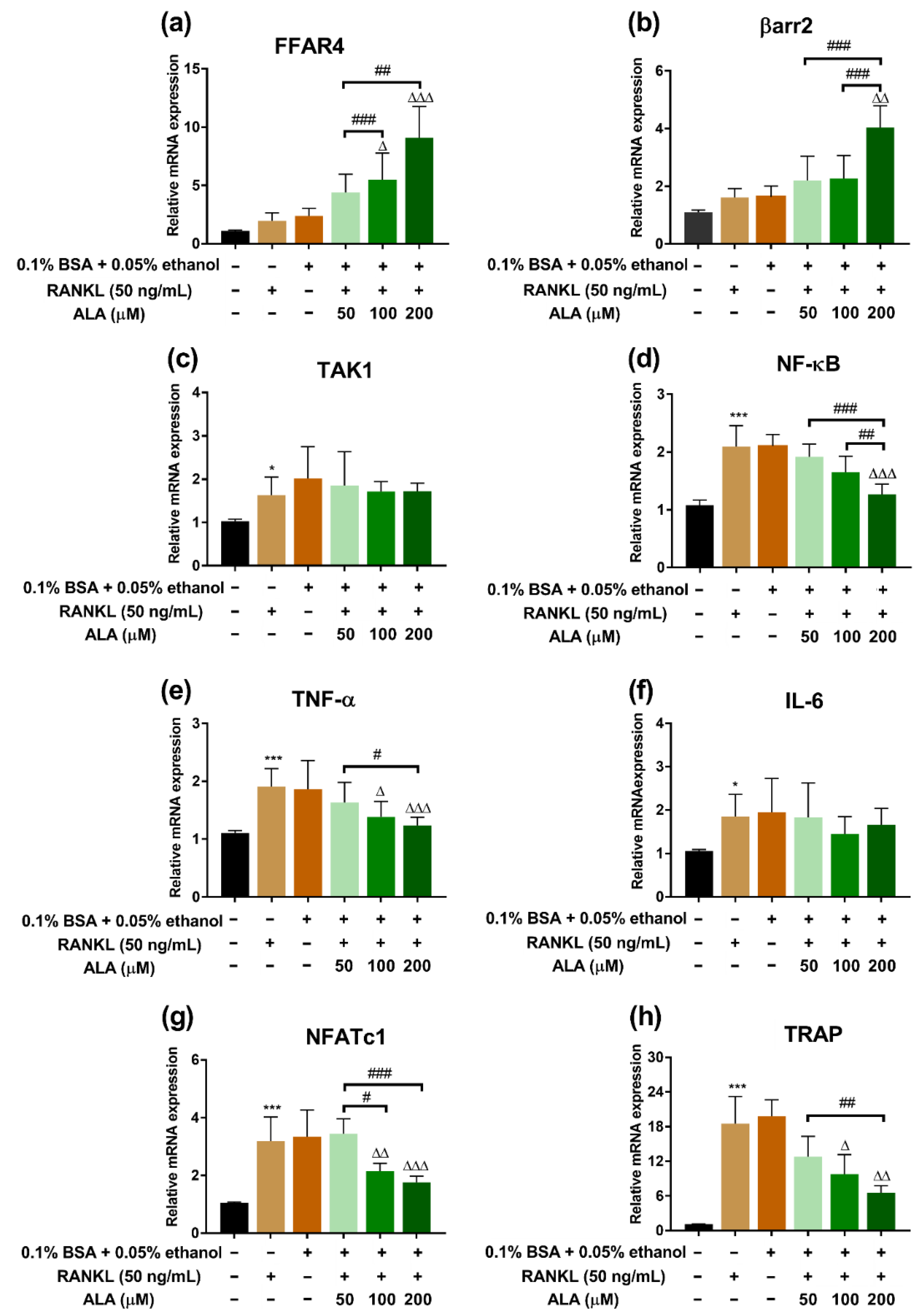
3.2. ALA Inhibited RANKL-Induced Osteoclastogenesis Genes Expression
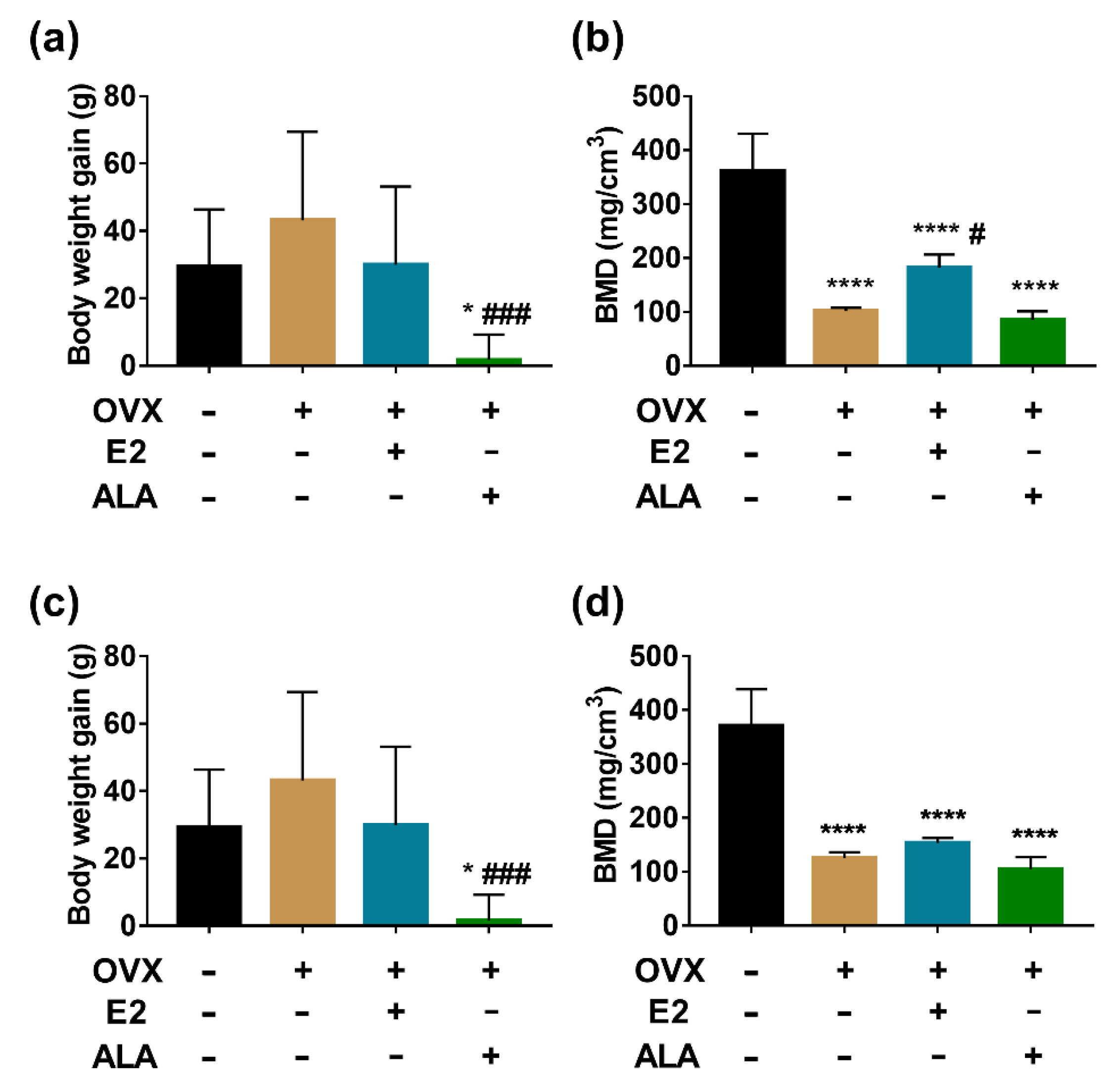
3.3. Effects of ALA on Body Weight and BMD
3.4. Effects of ALA on Routine Blood Indexes
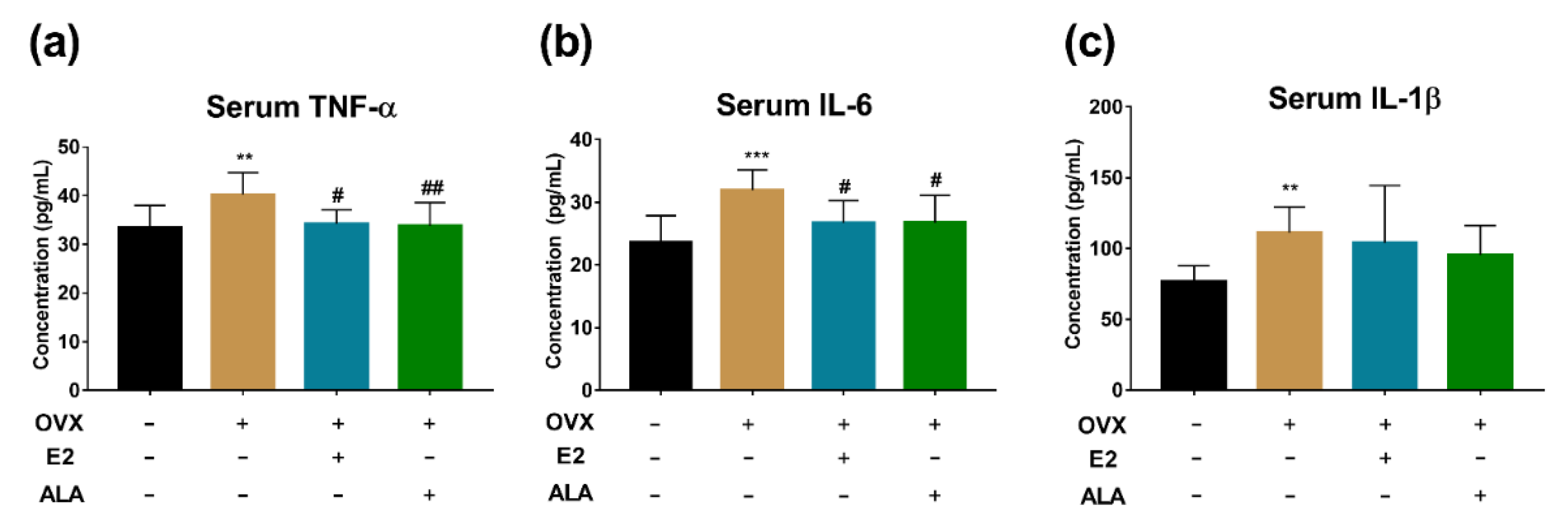
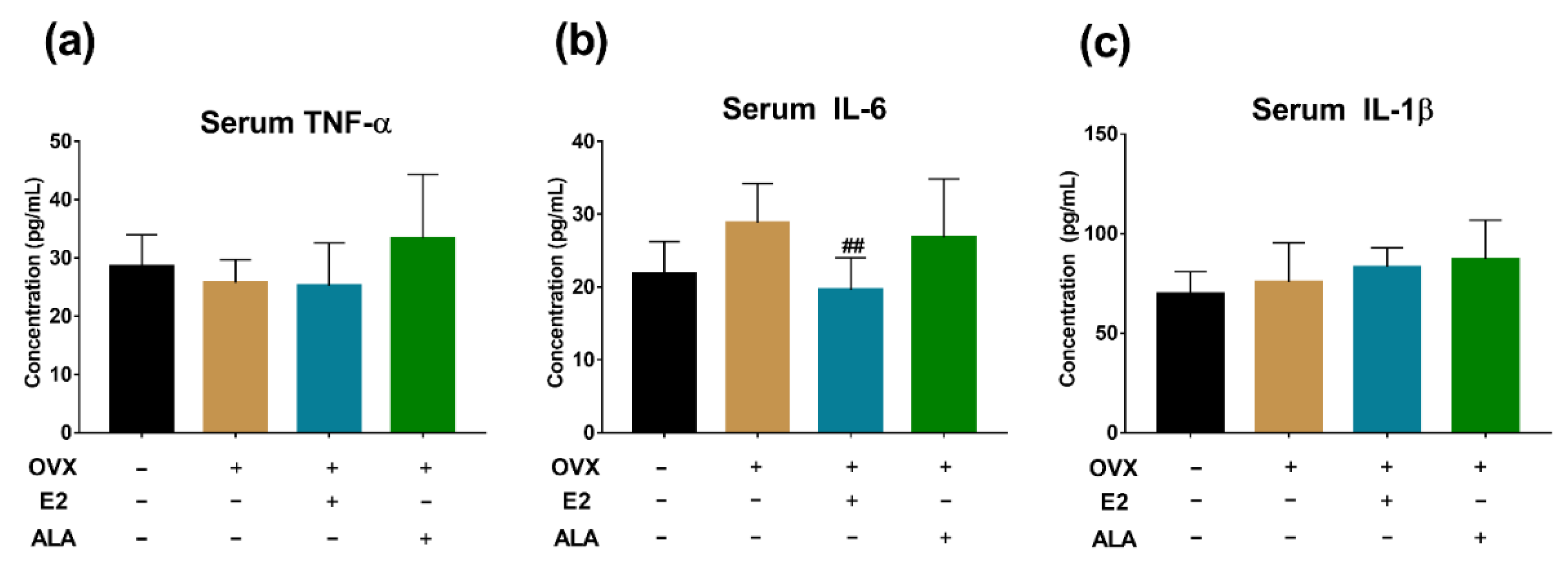
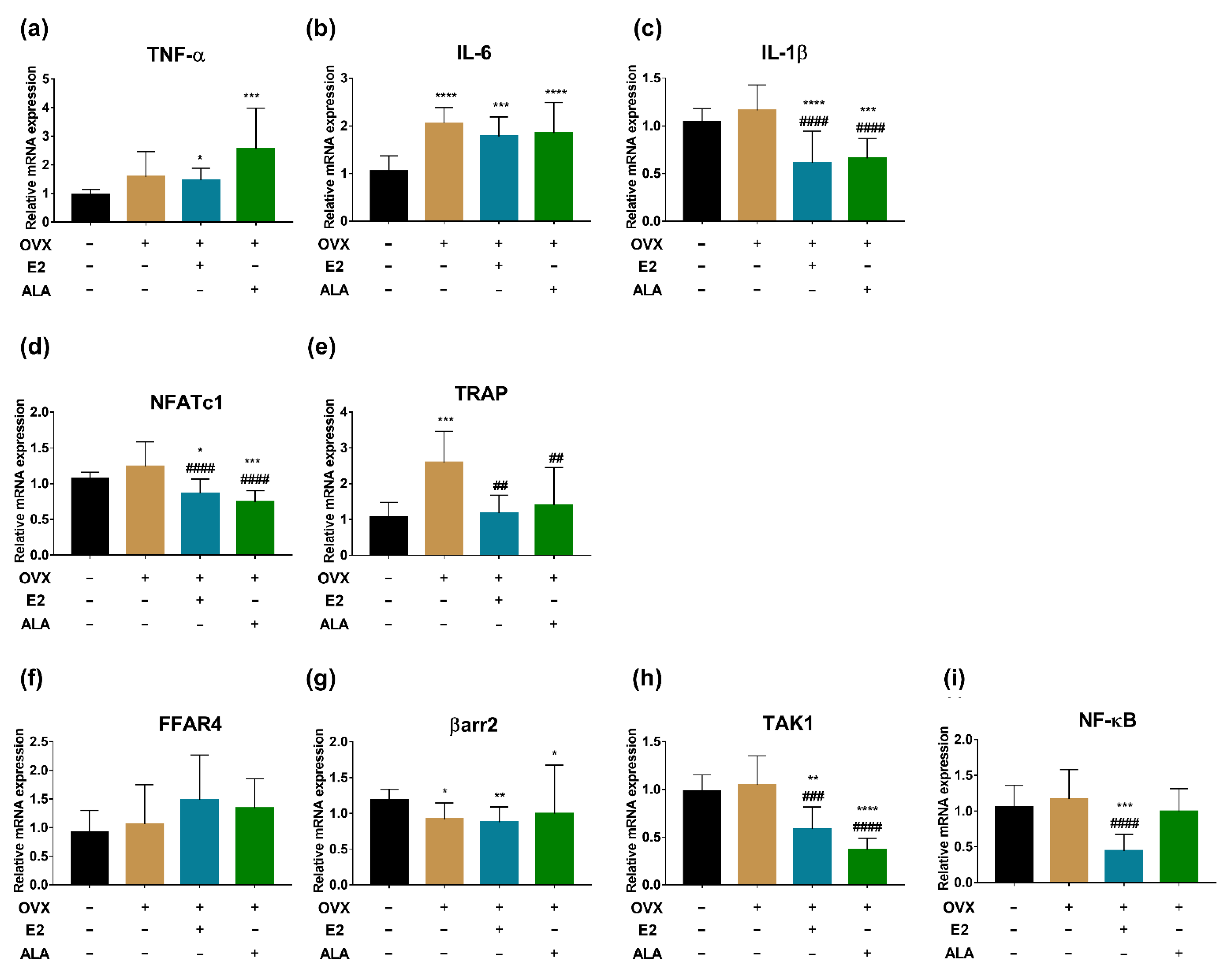
3.5. Effects of ALA on Serum Inflammatory Factors In Vivo
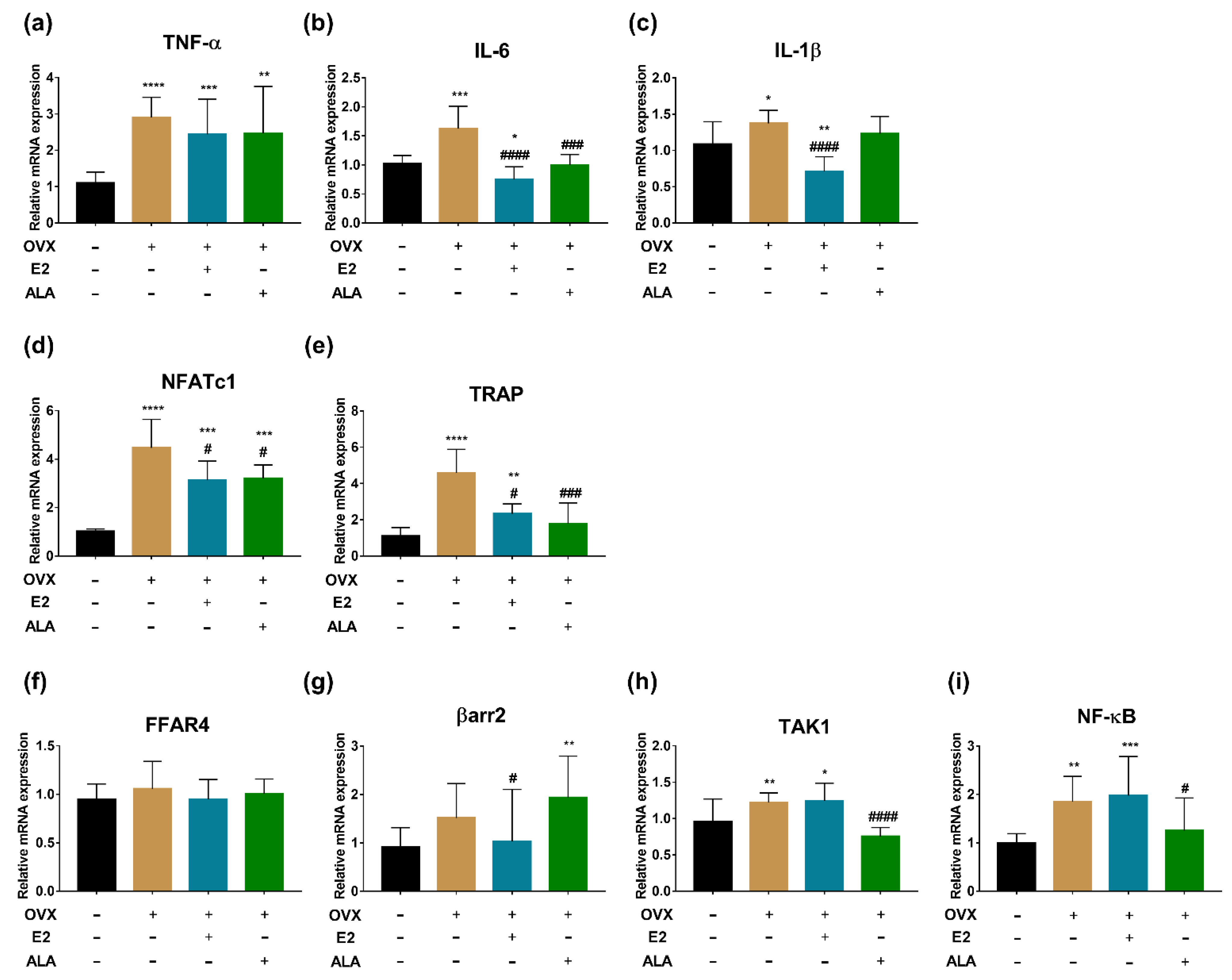
3.6. Effects of ALA on Expression of Inflammatory Factors and Osteoclast Genes in Bone of Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Scrivo, R.; Vasile, M.; Bartosiewicz, I.; Valesini, G. Inflammation as “common soil” of the multifactorial diseases. Autoimmun. Rev. 2011, 10, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Gordon, I.O.; Abushamma, S.; Kurowski, J.A.; Holubar, S.D.; Kou, L.; Lyu, R.; Rieder, F. Paediatric Ulcerative Colitis is a Fibrotic Disease and is Linked with Chronicity of Inflammation. J. Crohn’s Colitis 2022, 16, 804–821. [Google Scholar] [CrossRef] [PubMed]
- Sethwala, A.M.; Goh, I.; Amerena, J.V. Combating Inflammation in Cardiovascular Disease. Heart Lung Circ. 2021, 30, 197–206. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Matsumae, G.; Shimizu, T.; Takahashi, D.; Kadoya, K.; Iwasaki, N. Interplay between Inflammation and Pathological Bone Resorption: Insights into Recent Mechanisms and Pathways in Related Diseases for Future Perspectives. Int. J. Mol. Sci. 2022, 23, 1786. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Oursler, M.J.; Monroe, D.G. Estrogen and the skeleton. Trends Endocrinol. Metab. TEM 2012, 23, 576–581. [Google Scholar] [CrossRef]
- Tao, H.; Li, W.; Zhang, W.; Yang, C.; Zhang, C.; Liang, X.; Yin, J.; Bai, J.; Ge, G.; Zhang, H.; et al. Urolithin A suppresses RANKL-induced osteoclastogenesis and postmenopausal osteoporosis by, suppresses inflammation and downstream NF-κB activated pyroptosis pathways. Pharmacol. Res. 2021, 174, 105967. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, K.A.; Papavassiliou, A.G. Redox and NF-κB signaling in osteoarthritis. Free Radic. Biol. Med. 2019, 132, 90–100. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, X.; Li, D.; Cai, L.; Xu, Q. METTL3 Regulates Osteoblast Differentiation and Inflammatory Response via Smad Signaling and MAPK Signaling. Int. J. Mol. Sci. 2019, 21, 199. [Google Scholar] [CrossRef]
- Qiu, J.; Jiang, T.; Yang, G.; Gong, Y.; Zhang, W.; Zheng, X.; Hong, Z.; Chen, H. Neratinib exerts dual effects on cartilage degradation and osteoclast production in Osteoarthritis by inhibiting the activation of the MAPK/NF-κB signaling pathways. Biochem. Pharmacol. 2022, 205, 115155. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Lu, J. Interleukin-35 Promote Osteogenesis and Inhibit Adipogenesis: Role of Wnt/β-Catenin and PPARγ Signaling Pathways. Inflammation 2022. [Google Scholar] [CrossRef]
- Ruaro, B.; Casabella, A.; Paolino, S.; Pizzorni, C.; Ghio, M.; Seriolo, C.; Molfetta, L.; Odetti, P.; Smith, V.; Cutolo, M. Dickkopf-1 (Dkk-1) serum levels in systemic sclerosis and rheumatoid arthritis patients: Correlation with the Trabecular Bone Score (TBS). Clin. Rheumatol. 2018, 37, 3057–3062. [Google Scholar] [CrossRef]
- Blaschke, M.; Koepp, R.; Cortis, J.; Komrakova, M.; Schieker, M.; Hempel, U.; Siggelkow, H. IL-6, IL-1β, and TNF-α only in combination influence the osteoporotic phenotype in Crohn’s patients via bone formation and bone resorption. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2018, 27, 45–56. [Google Scholar] [CrossRef]
- Trombetta, A.C.; Smith, V.; Gotelli, E.; Ghio, M.; Paolino, S.; Pizzorni, C.; Vanhaecke, A.; Ruaro, B.; Sulli, A.; Cutolo, M. Vitamin D deficiency and clinical correlations in systemic sclerosis patients: A retrospective analysis for possible future developments. PLoS ONE 2017, 12, e0179062. [Google Scholar] [CrossRef]
- Redlich, K.; Smolen, J.S. Inflammatory bone loss: Pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef]
- Lin, J.; Zhu, J.; Wang, Y.; Zhang, N.; Gober, H.J.; Qiu, X.; Li, D.; Wang, L. Chinese single herbs and active ingredients for postmenopausal osteoporosis: From preclinical evidence to action mechanism. Biosci. Trends 2017, 11, 496–506. [Google Scholar] [CrossRef]
- Allijn, I.E.; Brinkhuis, R.P.; Storm, G.; Schiffelers, R.M. Anti-Inflammatory Properties of Plant Derived Natural Products—A Systematic Review. Curr. Med. Chem. 2019, 26, 4506–4536. [Google Scholar] [CrossRef]
- Feng, S.; Liu, Z.; Hu, Y.; Tian, J.; Yang, T.; Wei, A. Genomic analysis reveals the genetic diversity, population structure, evolutionary history and relationships of Chinese pepper. Hortic. Res. 2020, 7, 158. [Google Scholar] [CrossRef]
- Tang, W.; Xie, Q.; Guan, J.; Jin, S.; Zhao, Y. Phytochemical profiles and biological activity evaluation of Zanthoxylum bungeanum Maxim seed against asthma in murine models. J. Ethnopharmacol. 2014, 152, 444–450. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, J.; Zhu, L.; Li, T.; Jiang, W.; Zhou, J.; Peng, W.; Wu, C. Zanthoxylum bungeanum Maxim. (Rutaceae): A Systematic Review of Its Traditional Uses, Botany, Phytochemistry, Pharmacology, Pharmacokinetics, and Toxicology. Int. J. Mol. Sci. 2017, 18, 2172. [Google Scholar] [CrossRef]
- Yang, Q.; Cao, W.; Zhou, X.; Cao, W.; Xie, Y.; Wang, S. Anti-thrombotic effects of alpha-linolenic acid isolated from Zanthoxylum bungeanum Maxim seeds. BMC Complement. Altern. Med. 2014, 14, 348. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Wang, J.; Meng, J.; Zhang, X.; Niu, Y.; Gao, J.; Bai, Y.; Zhou, J. Zanthoxylum bungeanum Seed Oil Attenuates LPS-Induced BEAS-2B Cell Activation and Inflammation by Inhibiting the TLR4/MyD88/NF-kappaB Signaling Pathway. Evid. Based Complement. Altern. Med. Ecam 2021, 2021, 2073296. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Kang, R.; Huo, J.C.; Xie, Y.H.; Wang, S.W.; Cao, W. Wound-healing Activity of Zanthoxylum bungeanum Maxim Seed Oil on Experimentally Burned Rats. Pharmacogn. Mag. 2017, 13, 363–371. [Google Scholar] [PubMed]
- Pang, W.; Liu, S.; He, F.; Li, X.; Saira, B.; Zheng, T.; Chen, J.; Dong, K.; Pei, X.F. Anticancer activities of Zanthoxylum bungeanum seed oil on malignant melanoma. J. Ethnopharmacol. 2019, 229, 180–189. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Luo, S.; Liu, S.; Wan, S.; Li, J.; Chen, J.; Zuo, H.; Pei, X. Zanthoxylum bungeanum seed oil inhibits RANKL-induced osteoclastogenesis by suppressing ERK/c-JUN/NFATc1 pathway and regulating cell cycle arrest in RAW264.7 cells. J. Ethnopharmacol. 2022, 289, 115094. [Google Scholar] [CrossRef]
- Bao, M.; Zhang, K.; Wei, Y.; Hua, W.; Gao, Y.; Li, X.; Ye, L. Therapeutic potentials and modulatory mechanisms of fatty acids in bone. Cell Prolif. 2020, 53, e12735. [Google Scholar] [CrossRef]
- Wu, J.; Wu, Q.; Li, J.J.; Chen, C.; Sun, S.; Wang, C.H.; Sun, S.R. Autophagy mediates free fatty acid effects on MDA-MB-231 cell proliferation, migration and invasion. Oncol. Lett. 2017, 14, 4715–4721. [Google Scholar] [CrossRef]
- Yang, M.; Wei, D.; Mo, C.; Zhang, J.; Wang, X.; Han, X.; Wang, Z.; Xiao, H. Saturated fatty acid palmitate-induced insulin resistance is accompanied with myotube loss and the impaired expression of health benefit myokine genes in C2C12 myotubes. Lipids Health Dis. 2013, 12, 104. [Google Scholar] [CrossRef]
- Fu, L.; Pan, F.; Jiao, Y. Crocin inhibits RANKL-induced osteoclast formation and bone resorption by suppressing NF-κB signaling pathway activation. Immunobiology 2017, 222, 597–603. [Google Scholar] [CrossRef]
- Van Heerden, B.; Kasonga, A.; Kruger, M.C.; Coetzee, M. Palmitoleic Acid Inhibits RANKL-Induced Osteoclastogenesis and Bone Resorption by Suppressing NF-κB and MAPK Signalling Pathways. Nutrients 2017, 9, 441. [Google Scholar] [CrossRef]
- Anbazhagan, A.N.; Priyamvada, S.; Gujral, T.; Bhattacharyya, S.; Alrefai, W.A.; Dudeja, P.K.; Borthakur, A. A novel anti-inflammatory role of GPR120 in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2016, 310, C612–C621. [Google Scholar] [CrossRef] [Green Version]
- Kasonga, A.E.; Kruger, M.C.; Coetzee, M. Free fatty acid receptor 4-β-arrestin 2 pathway mediates the effects of different classes of unsaturated fatty acids in osteoclasts and osteoblasts. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2019, 1864, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.J.F.; Maguire, A.R.R.; Rouillard, M.M.; Crozier, R.W.E.; Yousef, M.; Bruton, K.M.; Fajardo, V.A.; MacNeil, A.J. TAK1 signaling activity links the mast cell cytokine response and degranulation in allergic inflammation. J. Leukoc. Biol. 2020, 107, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xiao, F.; Qu, X.; Zhai, Z.; Hu, G.; Chen, X.; Zhang, X. Sitagliptin, An Anti-diabetic Drug, Suppresses Estrogen Deficiency-Induced OsteoporosisIn Vivo and Inhibits RANKL-Induced Osteoclast Formation and Bone Resorption In Vitro. Front. Pharmacol. 2017, 8, 407. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, X.; Zhou, Z.; Mao, Y.; Wang, Y.; Ma, Z.; Xu, W.; Qin, A.; Zhang, S. Nirogacestat suppresses RANKL-Induced osteoclast formation in vitro and attenuates LPS-Induced bone resorption in vivo. Exp. Cell Res. 2019, 382, 111470. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ni, H.; Lan, L.; Wei, X.; Xiang, R.; Wang, Y. Fra-1 protooncogene regulates IL-6 expression in macrophages and promotes the generation of M2d macrophages. Cell Res. 2010, 20, 701–712. [Google Scholar] [CrossRef]
- Zhou, M.; Tang, Y.; Liao, L.; Liu, M.; Deng, Y.; Zhao, X.; Li, Y. Phillygenin inhibited LPS-induced RAW 264.7 cell inflammation by NF-κB pathway. Eur. J. Pharmacol. 2021, 899, 174043. [Google Scholar] [CrossRef]
- Jin, W.; Zhao, Y.; Hu, Y.; Yu, D.; Li, X.; Qin, Y.; Kong, D.; Wang, H. Stromal Cell-Derived Factor-1 Enhances the Therapeutic Effects of Human Endometrial Regenerative Cells in a Mouse Sepsis Model. Stem Cells Int. 2020, 2020, 4820543. [Google Scholar] [CrossRef]
- Hu, B.; Sun, X.; Yang, Y.; Ying, Z.; Meng, J.; Zhou, C.; Jiang, G.; Li, S.; Wu, F.; Zhao, X.; et al. Tomatidine suppresses osteoclastogenesis and mitigates estrogen deficiency-induced bone mass loss by modulating TRAF6-mediated signaling. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 2574–2586. [Google Scholar] [CrossRef]
- Lu, M.; Zhou, E. Long noncoding RNA LINC00662-miR-15b-5p mediated GPR120 dysregulation contributes to osteoarthritis. Pathol. Int. 2020, 70, 155–165. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, M.; Wang, X.; Zhang, H.; Zhang, J.; Jing, L.; Liao, L.; Wang, M. TNF Accelerates Death of Mandibular Condyle Chondrocytes in Rats with Biomechanical Stimulation-Induced Temporomandibular Joint Disease. PLoS ONE 2015, 10, e0141774. [Google Scholar] [CrossRef] [Green Version]
- Karim, K.; Giribabu, N.; Salleh, N. Marantodes pumilum Var Alata (Kacip Fatimah) ameliorates derangement in RANK/RANKL/OPG pathway and reduces inflammation and oxidative stress in the bone of estrogen-deficient female rats with type-2 diabetes. Phytomedicine Int. J. Phytother. Phytopharm. 2021, 91, 153677. [Google Scholar] [CrossRef]
- Li, J.B.; Xu, L.J.; Dong, H.; Huang, Z.Y.; Zhao, Y.; Chen, G.; Lu, F.E. Effects of Chinese Fructus Mume formula and its separated prescription extract on insulin resistance in type 2 diabetic rats. J. Huazhong Univ. Sci. Technology. Med. Sci. 2013, 33, 877–885. [Google Scholar] [CrossRef]
- Tian, Q.; Xiao, Q.; Yu, W.; Gu, M.; Zhao, N.; Lü, Y. The inhibition of transforming growth factor beta-activated kinase 1 contributed to neuroprotection via inflammatory reaction in pilocarpine-induced rats with epilepsy. Neuroscience 2016, 325, 111–123. [Google Scholar] [CrossRef]
- Mamipour, M.; Yousefi, M.; Dehnad, A.; Faridvand, Y.; Zarezadeh, R.; Khaksar, M.; Pouyafar, A.; Rahbarghazi, R. Protective effect of bacterial lipase on lipopolysaccharide-induced toxicity in rat cardiomyocytes; H9C2 cell line. J. Cardiovasc. Thorac. Res. 2020, 12, 35–42. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Fan, H.; Tang, S.; Weng, J. The preventive effect of Cuscutae Semen polysaccharide on bone loss in the ovariectomized rat model. Biomed. Pharmacother. 2020, 130, 110613. [Google Scholar] [CrossRef]
- Yuan, Q.; Xie, F.; Huang, W.; Hu, M.; Yan, Q.; Chen, Z.; Zheng, Y.; Liu, L. The review of alpha-linolenic acid: Sources, metabolism, and pharmacology. Phytother. Res. 2022, 36, 164–188. [Google Scholar] [CrossRef]
- Kim, K.B.; Nam, Y.A.; Kim, H.S.; Hayes, A.W.; Lee, B.M. alpha-Linolenic acid: Nutraceutical, pharmacological and toxicological evaluation. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2014, 70, 163–178. [Google Scholar] [CrossRef]
- Song, J.; Jing, Z.; Hu, W.; Yu, J.; Cui, X. alpha-Linolenic Acid Inhibits Receptor Activator of NF-kappaB Ligand Induced (RANKL-Induced) Osteoclastogenesis and Prevents Inflammatory Bone Loss via Downregulation of Nuclear Factor-KappaB-Inducible Nitric Oxide Synthases (NF-kappaB-iNOS) Signaling Pathways. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2017, 23, 5056–5069. [Google Scholar]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef]
- Grundmann, M.; Bender, E.; Schamberger, J.; Eitner, F. Pharmacology of Free Fatty Acid Receptors and Their Allosteric Modulators. Int. J. Mol. Sci. 2021, 22, 1763. [Google Scholar] [CrossRef]
- Oh, D.Y.; Walenta, E. Omega-3 Fatty Acids and FFAR4. Front. Endocrinol. 2014, 5, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Wang, X.; Liu, Y.; He, A.; Jia, R. NFATc1: Functions in osteoclasts. Int. J. Biochem. Cell Biol. 2010, 42, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Kucukgoncu, S.; Zhou, E.; Lucas, K.B.; Tek, C. Alpha-lipoic acid (ALA) as a supplementation for weight loss: Results from a meta-analysis of randomized controlled trials. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2017, 18, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Mohammadshahi, M.; Zakizadeh, E.; Ahmadi-Angali, K.; Ravanbakhsh, M.; Helli, B. The effect of alpha-lipoic acid supplementation and electrical isotonic contraction on anthropometric parameters, body composition and angiogenesis factor, sirtunin-1 and peroxisome proliferator-activated receptor-γ coactivator-1α in obese people under a weight loss regime: A study protocol for a randomized controlled clinical trial. Nutr. Health 2021, 27, 123–128. [Google Scholar]
- Polat, B.; Halici, Z.; Cadirci, E.; Albayrak, A.; Karakus, E.; Bayir, Y.; Bilen, H.; Sahin, A.; Yuksel, T.N. The effect of alpha-lipoic acid in ovariectomy and inflammation-mediated osteoporosis on the skeletal status of rat bone. Eur. J. Pharmacol. 2013, 718, 469–474. [Google Scholar] [CrossRef]
- Fu, C.; Xu, D.; Wang, C.Y.; Jin, Y.; Liu, Q.; Meng, Q.; Liu, K.X.; Sun, H.J.; Liu, M.Z. Alpha-Lipoic Acid Promotes Osteoblastic Formation in H2O2 -Treated MC3T3-E1 Cells and Prevents Bone Loss in Ovariectomized Rats. J. Cell. Physiol. 2015, 230, 2184–2201. [Google Scholar] [CrossRef]
- Han, X.M.; Wu, S.X.; Wu, M.F.; Yang, X.F. Antioxidant effect of peony seed oil on aging mice. Food Sci. Biotechnol. 2017, 26, 1703–1708. [Google Scholar] [CrossRef]
- Kaveh, M.; Eidi, A.; Nemati, A.; Boskabady, M.H. The Extract of Portulaca oleracea and Its Constituent, Alpha Linolenic Acid Affects Serum Oxidant Levels and Inflammatory Cells in Sensitized Rats. Iran. J. Allergy Asthma Immunol. 2017, 16, 256–270. [Google Scholar]
- Ginaldi, L.; de Martinis, M. Osteoimmunology and Beyond. Curr. Med. Chem. 2016, 23, 3754–3774. [Google Scholar] [CrossRef]
- Fang, H.; Zhang, H.; Wang, Z.; Zhou, Z.; Li, Y.; Lu, L. Systemic immune-inflammation index acts as a novel diagnostic biomarker for postmenopausal osteoporosis and could predict the risk of osteoporotic fracture. J. Clin. Lab. Anal. 2020, 34, e23016. [Google Scholar] [CrossRef]
- Gasparyan, A.Y.; Ayvazyan, L.; Mukanova, U.; Yessirkepov, M.; Kitas, G.D. The Platelet-to-Lymphocyte Ratio as an Inflammatory Marker in Rheumatic Diseases. Ann. Lab. Med. 2019, 39, 345–357. [Google Scholar] [CrossRef]
- Huang, C.; Li, S. Association of blood neutrophil lymphocyte ratio in the patients with postmenopausal osteoporosis. Pak. J. Med. Sci. 2016, 32, 762–765. [Google Scholar] [CrossRef]
- Koseoglu, S.B. Bone loss & platelet-to-lymphocyte ratio. Biomark. Med. 2017, 11, 5–10. [Google Scholar]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′→3′) | Reference |
|---|---|---|
| FFAR4 | Forward: CTGGGGCTCATCTTTGTCGT | [30] |
| Reverse: ACGACGAGCACTAGAGGGAT | ||
| βarr2 | Forward: ATCACTTGTTGAAAGTGGGC | [31] |
| Reverse: GTCTCGTCTTCAAGGATTGG | ||
| TAK1 | Forward: CCTCCTCGTCTTCTGCCAGTGA | [32] |
| Reverse: ACTCCAAAAGCTCCTCTTCCGACA | ||
| NFATc1 | Forward: CGTTGCTTCCAGAAAATAACA | [33] |
| Reverse: TGTGGGATGTGAACTCGGAA | ||
| TRAP | Forward: CTGGGGCTCATCTTTGTCGT | [34] |
| Reverse: CCCCAGAGACATGATGAAGTCA | ||
| IL-6 | Forward: CCACTTCACAAGTCGGAGGCTTA | [35] |
| Reverse: GCAAGTGCATCATCGTTGTTCATAC | ||
| TNF-α | Forward: ATGAGAAGTTCCCAAATGGC | [36] |
| Reverse: CTCCACTTGGTGGTTTGCTA | ||
| NF-κB | Forward: ATGGCAGACGATGATCCCTAC | [37] |
| Reverse: TGTTGACAGTGGTATTTCTGGTG | ||
| β-actin | Forward: TCTGCTGGAAGGTGGACAGT | [38] |
| Reverse: CCTCTATGCCAACACAGTGC |
| Gene | Primer Sequences (5′→3′) | Reference |
|---|---|---|
| Gapdh | Forward: AGTGCCAGCCTCGTCTCATA | [39] |
| Reverse: TGAACTTGCCGTGGGTAGAG | ||
| TNF-α | Forward: AACTCGAGTGACAAGCCCGTAG | [40] |
| Reverse: GTACCACCAGTTGGTTGTCTTTGA | ||
| IL-6 | Forward: ACCCCAACTTCCAATGCTC | [39] |
| Reverse: GGTTTGCCGAGTAGACCTC | ||
| IL-1β | Forward: CCTTGTCGAGAATGGGCAGT | [41] |
| Reverse: ACCAGAATGTGCCACGGTTT | ||
| FFAR4 | Forward: CCACCGTTCTGGGACTCATC | [39] |
| Reverse: CTCCACTTGGTGGTTTGCTA | ||
| βarr2 | Forward: TGGGCAACTCAAGCACGA | [42] |
| Reverse: AGCTTCACCTTGACCCTGTAGGA | ||
| TAK1 | Forward: AGCAGAAACGACAAGGCACT | [43] |
| Reverse: CAGCGAGACAGTGGATTTGA | ||
| NF-κB | Forward: GAGACCTGGAGCAAGCCATT | [44] |
| Reverse: CAGGCTAGGGTCAGCGTATG | ||
| NFATc1 | Forward: TGGAGAAGCAGAGCACAGAC | [45] |
| Reverse: GCGGAAAGGTGGTATCTCAA | ||
| TRAP | Forward: GCTGGAAACCATGATCACCT | [45] |
| Reverse: GAGTTGCCACACAGCATCAC |
| Detection Indexes | SHAM | OVX | E2 | ALA |
|---|---|---|---|---|
| WBC (109 per/mL) | 3.11 ± 0.57 | 4.61 ± 1.5 | 4.13 ± 1.59 | 8.14 ± 1.63 * |
| NE (109 per/mL) | 0.66 ± 0.32 | 1.14 ± 0.55 | 0.89 ± 0.54 | 1.03 ± 0.65 |
| LY (109 per/mL) | 3.11 ± 0.91 | 2.6 ± 0.98 | 4.4 ± 1.71 | 6.85 ± 1.23 # |
| MO (109 per/mL) | 0.26 ± 0.09 | 0.33 ± 0.17 | 0.35 ± 0.11 | 0.41 ± 0.16 |
| RBC (109 per/mL) | 7.40 ± 0.30 | 8.66 ± 0.28 * | 8.31 ± 0.37 | 8.17 ± 0.27 # |
| HGB (g/L) | 150.25 ± 6.2 | 170.88 ± 5.25 # | 163.13 ± 4.16 | 162.00 ± 6.28 |
| PLT (109 per/mL) | 689.38 ± 54.79 | 648 ± 76.81 | 707.13 ± 90.95 | 647.38 ± 88.91 |
| NLR | 0.21 ± 0.09 | 0.45 ± 0.21 * | 0.21 ± 0.09 # | 0.15 ± 0.09 # |
| PLR | 235.44 ± 56.62 | 285.85 ± 89.37 | 183.92 ± 70.64 # | 96.76 ± 18.95 *,# |
| Detection Indexes | SHAM | OVX | E2 | ALA |
|---|---|---|---|---|
| WBC (109 per/mL) | 2.36 ± 1.01 | 1.45 ± 0.36 | 1.70 ± 0.41 | 3.04 ± 1.10 # |
| NE (109 per/mL) | 0.25 ± 0.13 | 0.30 ± 0.18 | 0.30 ± 0.19 | 0.73 ± 0.30 |
| LY (109 per/mL) | 1.74 ± 0.26 | 1.13 ± 0.39 | 1.4 ± 0.42 | 2.55 ± 1.01 # |
| MO (109 per/mL) | 0.36 ± 0.11 | 0.29 ± 0.16 | 0.2 ± 0.11 | 0.31 ± 0.10 |
| RBC (109 per/mL) | 7.02 ± 0.13 | 7.57 ± 0.44 * | 7.54 ± 0.19 | 7.52 ± 0.70 |
| HGB (g/L) | 137.38 ± 3.34 | 146.13 ± 9.51 * | 145.75 ± 5.73 | 147.13 ± 8.46 |
| PLT (109 per/mL) | 899.38 ± 177.32 | 789.13 ± 63.73 | 757.25 ± 251.28 | 782.13 ± 148.11 |
| NLR | 0.17 ± 0.09 | 0.29 ± 0.18 * | 0.24 ± 0.13 | 0.30 ± 0.14 |
| PLR | 524.46 ± 107.96 | 768.39 ± 262.63 * | 552.06 ± 205.7 # | 343.34 ± 119.37 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; Li, W.; Zhang, Y.; Li, J.; He, F.; Dong, K.; Hong, Z.; Luo, R.; Pei, X. α-Linolenic Acid Inhibits RANKL-Induced Osteoclastogenesis In Vitro and Prevents Inflammation In Vivo. Foods 2023, 12, 682. https://doi.org/10.3390/foods12030682
Deng Y, Li W, Zhang Y, Li J, He F, Dong K, Hong Z, Luo R, Pei X. α-Linolenic Acid Inhibits RANKL-Induced Osteoclastogenesis In Vitro and Prevents Inflammation In Vivo. Foods. 2023; 12(3):682. https://doi.org/10.3390/foods12030682
Chicago/Turabian StyleDeng, Yufeng, Weizhou Li, Yingying Zhang, Jingjing Li, Fangting He, Ke Dong, Zehui Hong, Ruocheng Luo, and Xiaofang Pei. 2023. "α-Linolenic Acid Inhibits RANKL-Induced Osteoclastogenesis In Vitro and Prevents Inflammation In Vivo" Foods 12, no. 3: 682. https://doi.org/10.3390/foods12030682
APA StyleDeng, Y., Li, W., Zhang, Y., Li, J., He, F., Dong, K., Hong, Z., Luo, R., & Pei, X. (2023). α-Linolenic Acid Inhibits RANKL-Induced Osteoclastogenesis In Vitro and Prevents Inflammation In Vivo. Foods, 12(3), 682. https://doi.org/10.3390/foods12030682