
Screening of Bifidobacteria with Probiotic Potential from Healthy Infant Feces by Using 2′-Fucosyllactose
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Bifidobacteria
2.2. Strains and Culture Conditions
2.3. Screening of Bifidobacteria with Effective 2′-FL Utilization
2.4. Identification and Molecular Characterization of Isolated Bifidobacteria
2.5. Morphological Observation of Bifidobacterium by SEM
2.6. Determination of the Exopolysaccharide Yield
2.7. Determination of Bacterial Surface Protein Content
2.8. Determination of Auto-Aggregation Ability
2.9. Analysis of the of Simulated Gastric Juice and Bile Salt Tolerance
2.10. Determination of the Bacteriostatic Activity of Acellular Supernatant
2.11. Statistical Analyses
3. Results and Discussion
3.1. Screening of Bifidobacteria Able to Utilize of 2′-FL
3.2. Identification of Bifidobacterium by RAPD and 16S rDNA Pattern Analysis
3.3. Observation of Bifidobacterium Morphology by SEM
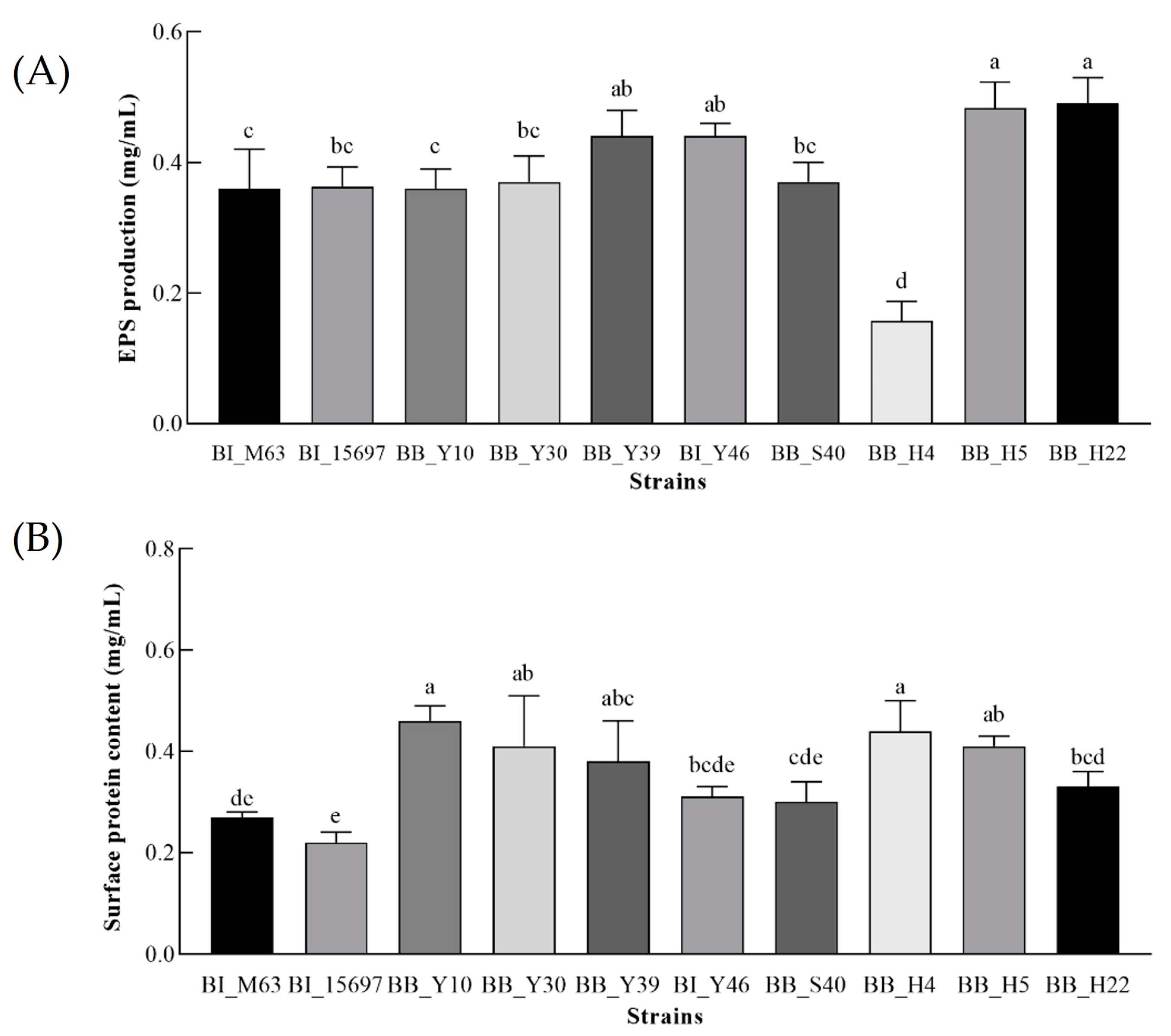
3.4. The EPS Yield of Bifidobacteria
3.5. Determination of the Bacterial Surface Protein Content
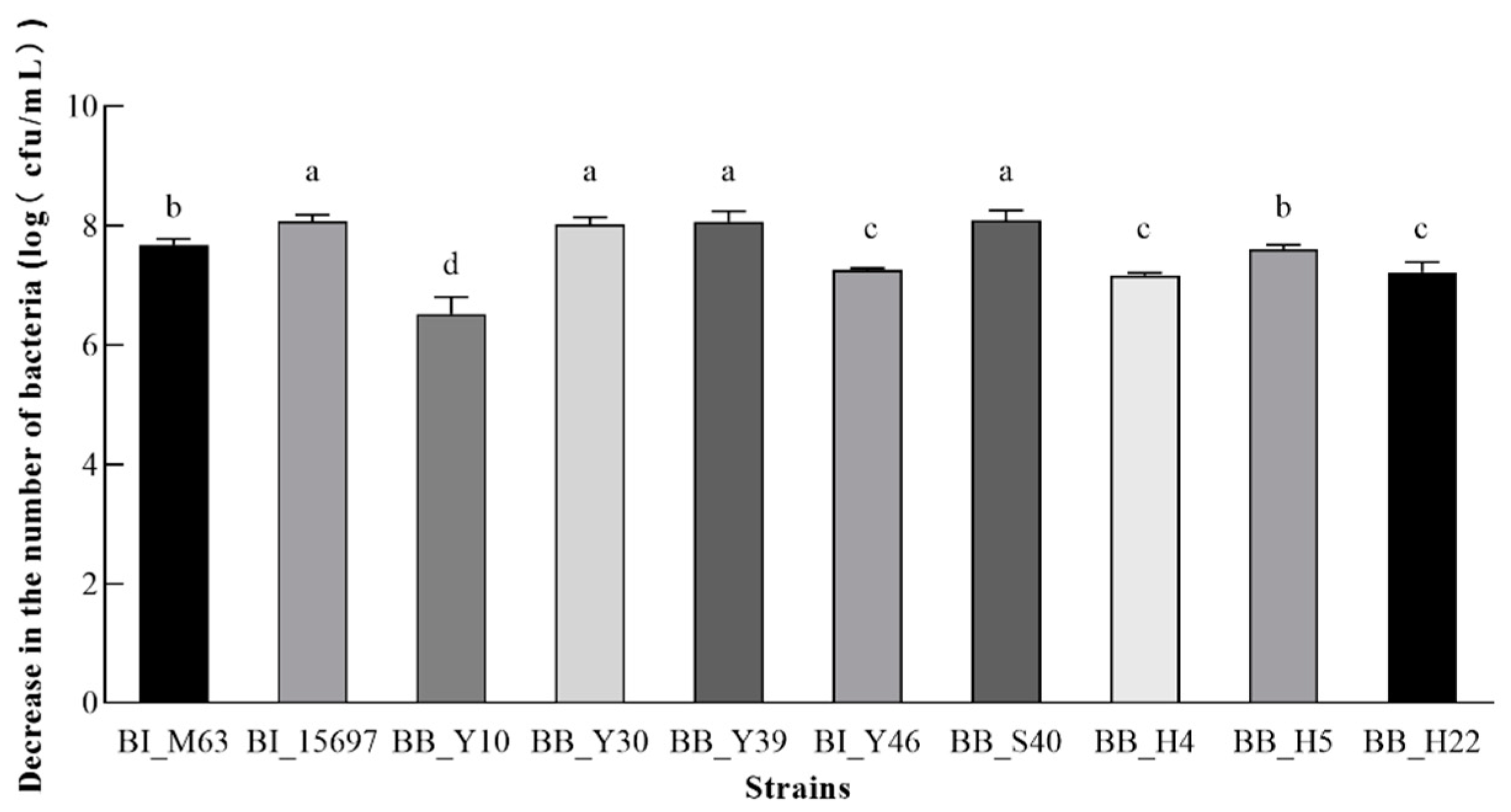
3.6. Analysis of Auto-Aggregation Ability
3.7. Analysis of Tolerance of Simulated Gastric Juice and Bile Salt
3.8. Evaluation of the Bacteriostatic Ability of Acellular Supernatant
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Quinn, E.M.; Joshi, L.; Hickey, R.M. Symposium review: Dairy-derived oligosaccharides-Their influence on host-microbe interactions in the gastrointestinal tract of infants. J. Dairy Sci. 2020, 103, 3816–3827. [Google Scholar] [CrossRef] [PubMed]
- Kulinich, A.; Liu, L. Human milk oligosaccharides: The role in the fine-tuning of innate immune responses. Carbohydr. Res. 2016, 432, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Thomson, P.; Medina, D.A.; Garrido, D. Human milk oligosaccharides and infant gut bifidobacteria: Molecular strategies for their utilization. Food Microbiol. 2018, 75, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Duysburgh, C.; Vazquez, E.; Chow, J.; Buck, R.; Marzorati, M. 2′-Fucosyllactose alters the composition and activity of gut microbiota from formula-fed infants receiving complementary feeding in a validated intestinal model. J. Funct. Foods 2019, 61, 103484. [Google Scholar] [CrossRef]
- Vassilis, T.; Lars, B.; Joost, V.N.R.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar]
- Yu, Z.-T.; Nanthakumar, N.N.; Newburg, D. The human milk oligosaccharide, 2′-fucosyllactose, quenches Campylobacter jejuni-induced inflammation in intestinal mucosa. FASEB J. 2015, 29, 853–856. [Google Scholar] [CrossRef]
- Mao, X.; Wang, J.; Hang, Y.; Zhang, Y.; Yu, H.; Li, Z.; Pan, L.; Zhiyong, D. A human milk oligosaccharide, 2′-fucosyllactose, enhances the immunity in mice fed an infant formula milk diet. Int. Dairy J. 2019, 98, 38–43. [Google Scholar] [CrossRef]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2′-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.; Malcata, F.X. Bifidobacterium spp. and Lactobacillus acidophilus: Biological, biochemical, technological and therapeutical properties relevant for use as probiotics-ScienceDirect. Trends Food Sci. Technol. 1999, 10, 139–157. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Delgado, S.; Ruiz, L.; Ruas-Madiedo, P.; Sanchez, B.; Margolles, A. Bifidobacteria and Their Health-Promoting Effects. Microbiol. Spectr. 2017, 5, 139–157. [Google Scholar] [CrossRef]
- Alessandri, G.; Ossiprandi, M.C.; MacSharry, J.; van Sinderen, D.; Ventura, M. Bifidobacterial dialogue with its human host and consequent modulation of the immune system. Front. Immunol. 2019, 10, 2348. [Google Scholar] [CrossRef] [Green Version]
- Wanping, A.; Shinji, F. Protective effects of bifidobacteria against enteropathogens. Microb. Biotechnol. 2019, 12, 1097–1100. [Google Scholar]
- Daniels, V.C.; Monaco, M.H.; Wang, M.; Hirvonen, J.; Jensen, H.M.; Ouwehand, A.C.; Mukherjea, R.; Dilger, R.N.; Donovan, S.M. Evaluation of 2′-Fucosyllactose and Bifidobacterium longum Subspecies infantis on Growth, Organ Weights, and Intestinal Development of Piglets. Nutrients 2022, 14, 199. [Google Scholar] [CrossRef] [PubMed]
- Hartemink, R.; Kok, B.J.; Weenk, G.H.; Rombouts, F.M. Raffinose-Bifidobacterium (RB) agar, a new selective medium for bifidobacteria. J. Microbiol. Methods 1996, 27, 10. [Google Scholar] [CrossRef]
- Barrangou, R.; Altermann, E.; Hutkins, R.; Cano, R.; Klaenhammer, T.R. Functional and comparative genomic analyses of an operon involved in fructooligosaccharide utilization by Lactobacillus acidophilus. Proc. Natl. Acad. Sci. USA 2003, 100, 8957–8962. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Sun, L.; Yi, H.; Wang, S.; Han, J.; Liu, N.; Zhang, S.; Zhang, L. Comparative proteome analysis of Streptomyces mobaraensis under MgCl2 stress shows proteins modulating differentiation and transglutaminase biosynthesis. Food Res. Int. 2019, 121, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chichlowski, M.; Gross, G.; Holle, M.J.; Lbarra-Sánchez, L.A.; Wang, S.; Miller, M.J. Milk Fat Globule Membrane Protects Lactobacillus rhamnosus GG from Bile Stress by Regulating Exopolysaccharide Production and Biofilm Formation. J. Agric. Food Chem. 2020, 68, 6646–6655. [Google Scholar] [CrossRef]
- McKellar, R.C.; van Geest, J.; Cui, W. Influence of culture and environmental conditions on the composition of exopolysaccharide produced by Agrobacterium radiobacter. Food Hydrocoll. 2003, 17, 429–437. [Google Scholar] [CrossRef]
- Zhang, Y.-C.; Zhang, L.-W.; Ma, W.; Yi, H.-X.; Yang, X.; Du, M.; Shan, Y.-J.; Han, X.; Zhang, L.-L. Screening of probiotic lactobacilli for inhibition of Shigella sonnei and the macromolecules involved in inhibition. Anaerobe 2012, 18, 498–503. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef]
- Zhuang, G.; Wang, J.; Yan, L.; Chen, W.; Liu, X.M.; Zhang, H.P. In vitro comparison of probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected probiotic strains. LWT-Food Sci. Technol. 2009, 42, 1640–1646. [Google Scholar]
- Chen, C.Y.; Nace, G.W.; Irwin, P.L. A 6 × 6 drop plate method for simultaneous colony counting and MPN enumeration of Campylobacter jejuni, Listeria monocytogenes, and Escherichia coli. J. Microbiol. Methods 2003, 55, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, Z.; Li, X.; Zhang, X.; Du, S.; Jia, M.; Hu, D.; Jia, X.; Cong, B.; Zhang, Y.; et al. A new Lactobacillus gasseri strain HMV18 inhibits the growth of pathogenic bacteria. Food Sci. Hum. Wellness 2022, 11, 247–254. [Google Scholar] [CrossRef]
- Thongaram, T.; Hoeflinger, J.L.; Chow, J.; Miller, M.J. Human milk oligosaccharide consumption by probiotic and human-associated bifidobacteria and lactobacilli. J. Dairy Sci. 2017, 100, 7825–7833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, F.; Serafini, F.; Foroni, E.; Duranti, S.; Motherway, M.O.C.; Taverniti, V.; Mangifesta, M.; Milani, C.; Viappiani, A.; Roversi, T.; et al. Role of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in modulating bacterium–host interactions. Proc. Natl. Acad. Sci. USA 2013, 110, 11151–11156. [Google Scholar] [CrossRef] [Green Version]
- He, Z.J.; Yang, B.; Liu, X.M.; Ross, R.P.; Stanton, C.; Zhao, J.X.; Zhang, H.; Chen, W. Genotype-phenotype association analysis revealed different utilization ability of 2′-fucosyllactose in Bifidobacterium genus. J. Dairy Sci. 2021, 104, 1518–1523. [Google Scholar] [CrossRef]
- Zabel, B.E.; Gerdes, S.; Evans, K.C.; Nedveck, D.; Singles, S.K.; Volk, B.; Budinoff, C. Strain-specific strategies of 2′-fucosyllactose, 3-fucosyllactose, and difucosyllactose assimilation by Bifidobacterium longum subsp. infantis Bi-26 and ATCC 15697. Sci. Rep. 2020, 10, 15919. [Google Scholar] [CrossRef]
- Sela, D.A.; Garrido, D.; Lerno, L.; Wu, S.A.; Tan, K.M.; Eom, H.J.; Joachimiak, A.; Lebrilla, C.B.; Mills, D.A. Bifidobacterium longum subsp infantis ATCC 15697 alpha-Fucosidases Are Active on Fucosylated Human Milk Oligosaccharides. Appl. Environ. Microbiol. 2012, 78, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. Human milk oligosaccharides: Shaping the infant gut microbiota and supporting health. J. Funct. Foods 2020, 72, 104074. [Google Scholar] [CrossRef]
- Laursen, M.F.; Sakanaka, M.; von Burg, N.; Morbe, U.; Andersen, D.; Moll, J.M.; Pekmez, C.T.; Rivollier, A.; Michaelsen, K.F.; Molgaard, C.; et al. Bifidobacterium species associated with breastfeeding produce aromatic lactic acids in the infant gut. Nat. Microbiol. 2021, 6, U1367–U1370. [Google Scholar] [CrossRef] [PubMed]
- Good, M.; Sodhi, C.P.; Yamaguchi, Y.; Jia, H.; Lu, P.; Fulton, W.B.; Martin, L.Y.; Prindle, T.; Nino, D.F.; Zhou, Q.; et al. The human milk oligosaccharide 2′-fucosyllactose attenuates the severity of experimental necrotising enterocolitis by enhancing mesenteric perfusion in the neonatal intestine. Br. J. Nutr. 2016, 116, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Zhai, Z.; Lei, Y.; Xiao, B.; Hao, Y. A Novel Major Pilin Subunit Protein FimM Is Involved in Adhesion of Bifidobacterium longum BBMN68 to Intestinal Epithelial Cells. Front. Microbiol. 2020, 11, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Motherway, M.O.; Zomer, A.; Leahy, S.C.; Reunanen, J.; Bottacini, F.; Claesson, M.J.; O’Brien, F.; Flynn, K.; Casey, P.G.; Munoz, J.A.M.; et al. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. USA 2011, 108, 11217–11222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanning, S.; Hall, L.J.; Cronin, M.; Zomer, A.; MacSharry, J.; Goulding, D.; Motherway, M.O.C.; Shanahan, F.; Nally, K.; Dougan, G. Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. USA 2012, 109, 2108–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donot, F.; Fontana, A.; Baccou, J.C.; Schorr-Galindo, S. Microbial exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction. Carbohydr. Polym. 2012, 87, 951–962. [Google Scholar] [CrossRef]
- Russell, D.A.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Metabolic activities and probiotic potential of bifidobacteria. Int. J. Food Microbiol. 2011, 149, 88–105. [Google Scholar] [CrossRef]
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiol.-Sgm 2009, 155, 1749–1757. [Google Scholar] [CrossRef] [Green Version]
- Hoeflinger, J.L.; Miller, M.J. Cronobacter sakazakii ATCC 29544 autoaggregation requires flic flagellation, Not Motility. Front. Microbiol. 2017, 8, 301. [Google Scholar] [CrossRef]
- Giulia, A.; Douwe, V.S.; Marco, V. The genus Bifidobacterium: From genomics to functionality of an important component of the mammalian gut microbiota running title: Bifidobacterial adaptation to and interaction with the host. Computational and Structural. Biotechnol. J. 2021, 19, 5. [Google Scholar] [CrossRef]
- Ruiz, L.; Margolles, A.; Sanchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiri, S.; Mokarram, R.R.; Khiabani, M.S.; Bari, M.R.; Khaledabad, M.A. Characterization of antimicrobial peptides produced by Lactobacillus acidophilus LA-5 and Bifidobacterium lactis BB-12 and their inhibitory effect against foodborne pathogens. LWT 2022, 153, 112449. [Google Scholar] [CrossRef]
- Monteiro, C.R.A.V.; do Carmo, M.S.; Melo, B.O.; Alves, M.S.; dos Santos, C.I.; Monteiro, S.G.; Bomfim, M.R.Q.; Fernandes, E.S.; Monteiro-Neto, V. In vitro antimicrobial activity and probiotic potential of Bifidobacterium and Lactobacillus against species of Clostridium. Nutrients 2019, 11, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]





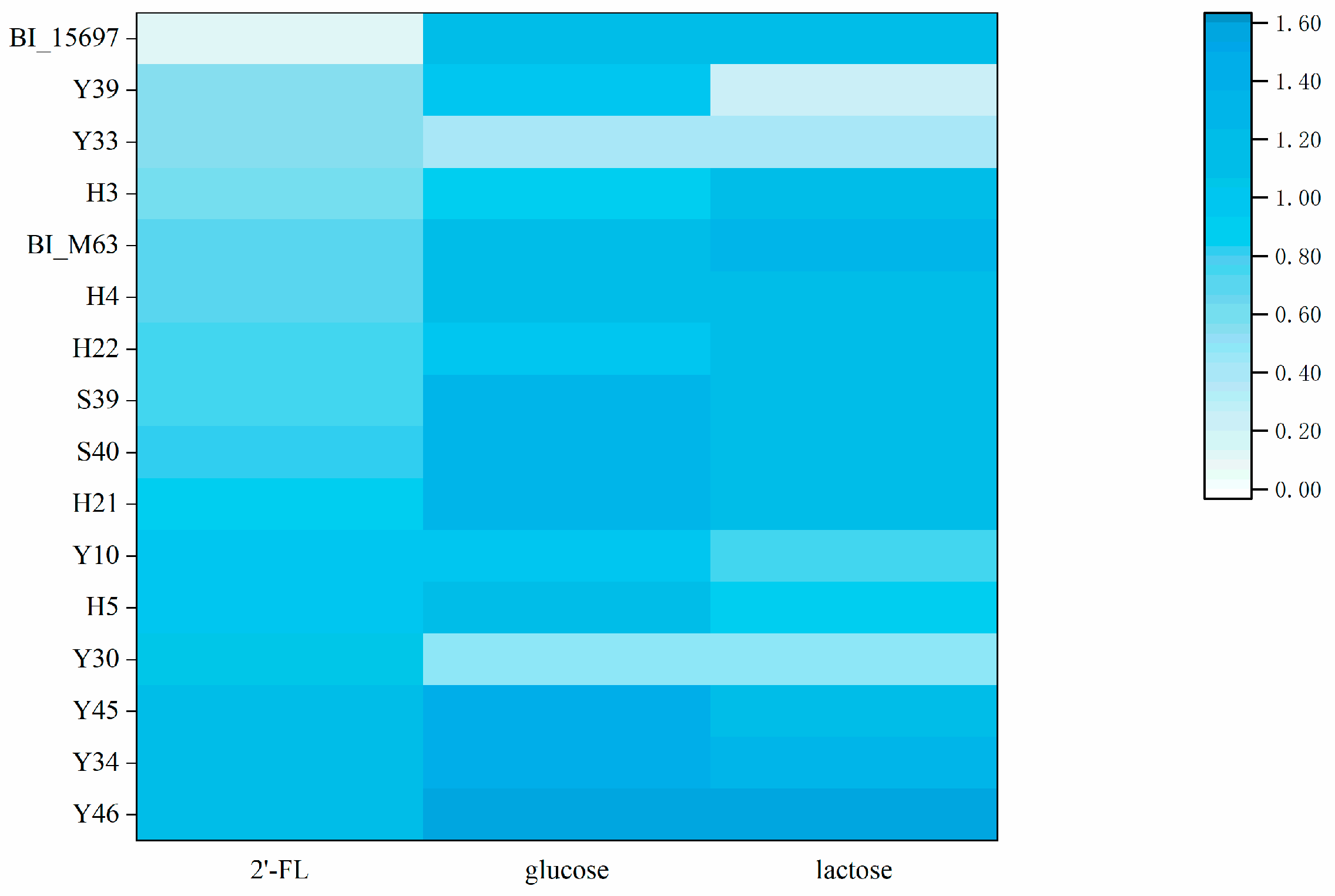{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 2 h (%) | 4 h (%) | 6 h (%) |
|---|---|---|---|
| BI_M63 | 17.67 ± 2.74 Cde | 27.37 ± 1.89 Bde | 38.98 ± 0.96 Ae |
| BI_15697 | 18.92 ± 1.49 Cde | 32.80 ± 3.82 Bd | 48.43 ± 0.90 Ad |
| BB_Y10 | 39.56 ± 3.52 Ca | 58.54 ± 5.15 Bb | 81.61 ± 3.47 Ab |
| BB_Y30 | 28.36 ± 2.75 Cbc | 51.31 ± 6.02 Bbc | 73.42 ± 1.25 Ac |
| BB_Y39 | 17.45 ± 2.24 Bde | 23.72 ± 3.35 Ae | 30.36 ± 0.29 Af |
| BI_Y46 | 30.56 ± 2.51 Cbc | 53.31 ± 4.55 Bbc | 68.56 ± 3.16 Ac |
| BB_S40 | 19.67 ± 0.73 Bde | 47.46 ± 2.06 Ac | 52.22 ± 2.15 Ad |
| BB_H4 | 23.56 ± 2.42 Ccd | 52.58 ± 3.44 Bbc | 72.88 ± 5.76 Ac |
| BB_H5 | 15.31 ± 0.95 Ce | 50.07 ± 2.08 Bbc | 69.56 ± 5.42 Ac |
| BB_H22 | 36.21 ± 3.23 Cab | 72.86 ± 3.31 Ba | 90.72 ± 2.31 Aa |
| Strains | 0 h (log (cfu/mL)) | 30 min (log (cfu/mL)) | 3 h (log (cfu/mL)) | 3 h Survival Rate (%) |
|---|---|---|---|---|
| BI_M63 | 7.08 ± 0.20 Cc | 7.95 ± 0.02 Bbc | 8.23 ± 0.15 Aa | 116.25 ± 2.58 a |
| BI_15697 | 7.88 ± 0.02 Aab | 6.09 ± 0.08 Be | 5.46 ± 0.28 Cf | 69.32 ± 3.56 d |
| BB_Y10 | 7.14 ± 0.26 Ac | 7.27 ± 0.01 Ad | 7.18 ± 0.14 Ad | 100.58 ± 2.70 b |
| BB_Y30 | 7.81 ± 0.04 Aab | 7.85 ± 0.02 Abc | 7.80 ± 0.06 Abc | 99.34 ± 0.71 b |
| BB_Y39 | 7.97 ± 0.02 Aa | 6.48 ± 0.07 Be | 6.13 ± 0.16 Ce | 76.89 ± 2.04 c |
| BI_Y46 | 7.88 ± 0.04 Aab | 7.76 ± 0.01 Bc | 7.81 ± 0.03 Abc | 99.06 ± 0.45 b |
| BB_S40 | 7.72 ± 0.05 Ab | 7.77 ± 0.04 Abc | 7.63 ± 0.07 Bc | 98.72 ± 0.73 b |
| BB_H4 | 7.93 ± 0.02 Bab | 8.26 ± 0.07 Aab | 7.89 ± 0.04 Bb | 99.41 ± 0.63 b |
| BB_H5 | 7.99 ± 0.03 Aa | 7.68 ± 0.54 Bcd | 8.05 ± 0.03 Aab | 100.81 ± 0.87 b |
| BB_H22 | 8.05 ± 0.02 Ba | 8.66 ± 0.53 Aa | 7.95 ± 0.03 Bb | 98.88 ± 0.35 b |
| Strains | Bacteriostatic Effect |
|---|---|
| BI_M63 | ++++ |
| BI_15697 | - |
| BB_Y10 | +++ |
| BB_Y30 | +++ |
| BB_Y39 | ++++ |
| BI_Y46 | ++++ |
| BB_S40 | +++ |
| BB_H4 | ++ |
| BB_H5 | +++ |
| BB_H22 | +++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Sun, H.; Xu, Z.; Tan, Z.; Xiao, L.; He, M.; Shang, J.; Tsapieva, A.N.; Zhang, L. Screening of Bifidobacteria with Probiotic Potential from Healthy Infant Feces by Using 2′-Fucosyllactose. Foods 2023, 12, 858. https://doi.org/10.3390/foods12040858
Zhang G, Sun H, Xu Z, Tan Z, Xiao L, He M, Shang J, Tsapieva AN, Zhang L. Screening of Bifidobacteria with Probiotic Potential from Healthy Infant Feces by Using 2′-Fucosyllactose. Foods. 2023; 12(4):858. https://doi.org/10.3390/foods12040858
Chicago/Turabian StyleZhang, Gongsheng, Hui Sun, Zihe Xu, Ze Tan, Lihong Xiao, Mingxue He, Jiaqi Shang, Anna N. Tsapieva, and Lili Zhang. 2023. "Screening of Bifidobacteria with Probiotic Potential from Healthy Infant Feces by Using 2′-Fucosyllactose" Foods 12, no. 4: 858. https://doi.org/10.3390/foods12040858
APA StyleZhang, G., Sun, H., Xu, Z., Tan, Z., Xiao, L., He, M., Shang, J., Tsapieva, A. N., & Zhang, L. (2023). Screening of Bifidobacteria with Probiotic Potential from Healthy Infant Feces by Using 2′-Fucosyllactose. Foods, 12(4), 858. https://doi.org/10.3390/foods12040858