
The Development of Dyslipidemia in Chronic Kidney Disease and Associated Cardiovascular Damage, and the Protective Effects of Curcuminoids

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Pathophysiological Features of Dyslipidemia in CKD
3. Dyslipidemia in CKD-Induced Cardiovascular Damage
4. Mechanisms Involved in CKD-Induced Dyslipidemia and Associated Cardiovascular Damage
4.1. Oxidative Stress
4.2. Inflammation and Fibrosis
4.3. Metabolic Reprogramming
5. Drugs and Their Drawbacks in CKD-Induced Dyslipidemia and Associated Cardiovascular Damage
6. Curcuminoids
6.1. Curcuminoids Bioavailability
6.2. Curcuminoids on CKD

{kind=link}
{kind=link}
{kind=link}
| Reference | Model | Compound | Dosage | Effect |
|---|---|---|---|---|
| [132] | 5/6 nephrectomized Wistar rats | Curcumin | 120 mg/kg | Reduces proteinuria, creatinine, and BUN serum levels, Improves renal function and blood pressure. Decreases oxidative stress through the Nrf2 pathway and monocyte infiltration by the reduction of MCP-1 |
| [134] | Sprague–Dawley rats, renal injury induced by 0.25% adenine | Curcumin | 37.5, 75 and 150 mg/kg | Decreases renal damage markers, inflammation (IL-1ß and IL-6), and fibrosis (caspase 3); and increases antioxidant indices (glutathione and super oxide dismutase). |
| [131] | 5/6 nephrectomized Wistar rats | Curcumin | 60 mg/kg | Reduces proteinuria, creatinine, BUN serum levels, and systolic pressure. Improve renal hemodynamics and mitochondrial respiration. Decrease oxidative stress, interstitial inflammation, and fibrosis. |
| [130] | 5/6 nephrectomized Wistar rats | Curcumin | 60 and 120 mg/kg | Reverts glomerular and systemic hypertension. Restores kidney tubular atrophy, reduces the mesangial area and mesangial cells proliferation, prevents the expansion of the glomerular matrix |
| [135] | Dahl salt-sensitive rats, nephrosclerosis induced by salt | Curcumin | 10 mg/kg | The antifibrotic effect could be through the inhibition of histone acetylation (H3K9) |
| [133] | 5/6 nephrectomized Sprague–Dawley rats | Tetrahydro curcumin | 1% in food | Improves the expression of antioxidant enzymes in the kidney, decreases renal apoptosis and fibrosis and ameliorates proteinuria, hypertension, and cardiac hypertrophy. |
| [136] | Mice with unilateral ureteral obstruction | Bisdemethoxycurcumin | 100 and 200 mg/kg | Reduces fibrosis throw fibroblast apoptosis |
| Reference | Model | Compound | Dosage | Effect |
|---|---|---|---|---|
| [152] | CKD patients’ mononuclear cell culture | Curcumin | 1 and 3 mM | Decreases secretion of IL-6 and IL-1β. Decreases the procoagulant activity of mononuclear cells. |
| [127] | Clinical trial, CKD patients with coronary angiography or angioplasty | Curcumin | 1.5 g | There are no significant changes |
| [153] | Clinical trial, stage 3 to 4 CKD patients | Meriva® | 1 g/day | Increases plasma creatinine, decreases eGFR, and changes microbiota diversity. Reduces plasma pro-inflammatory mediators (MCP-1, IFN-γ, and IL-4) and lipid peroxidation. |
| [156] | Clinical trial, CKD patients with coronary angiography | Curcuminoids | 1.5 g/day | Preserve changes in eGFR preventing post-contrast acute kidney injury. |
| [157] | Clinical trial, non-dialysis CKD patients | Curcuminoids and Boswellia serrata | 824 and 510 mg/day, respectively | There was a time effect and time x compliance interaction effect for IL-6 |
| [158] | Clinical trial, stage 2 to 3 CKD patients | Curcuminoids and Boswellia serrata | 824 and 510 mg/day, respectively | There was a group effect and a trend for group × time interaction for prostaglandin E2. |
| [154] | Clinical trial, hemodialyzed CKD patients | Turmeric | 2.5 g | Decreases in NF-κB mRNA expression in mononuclear cells and in plasma high-sensitivity CRP levels |
| [126] | Clinical trial, hemodialyzed CKD patients | Turmeric | 2.5 g | Decreases in pCS plasma levels, suggesting gut microbiota regulation |
| [155] | Clinical trial, hemodialyzed CKD patients | Turmeric | 2.5 g | Reduces TNF-α plasma levels |
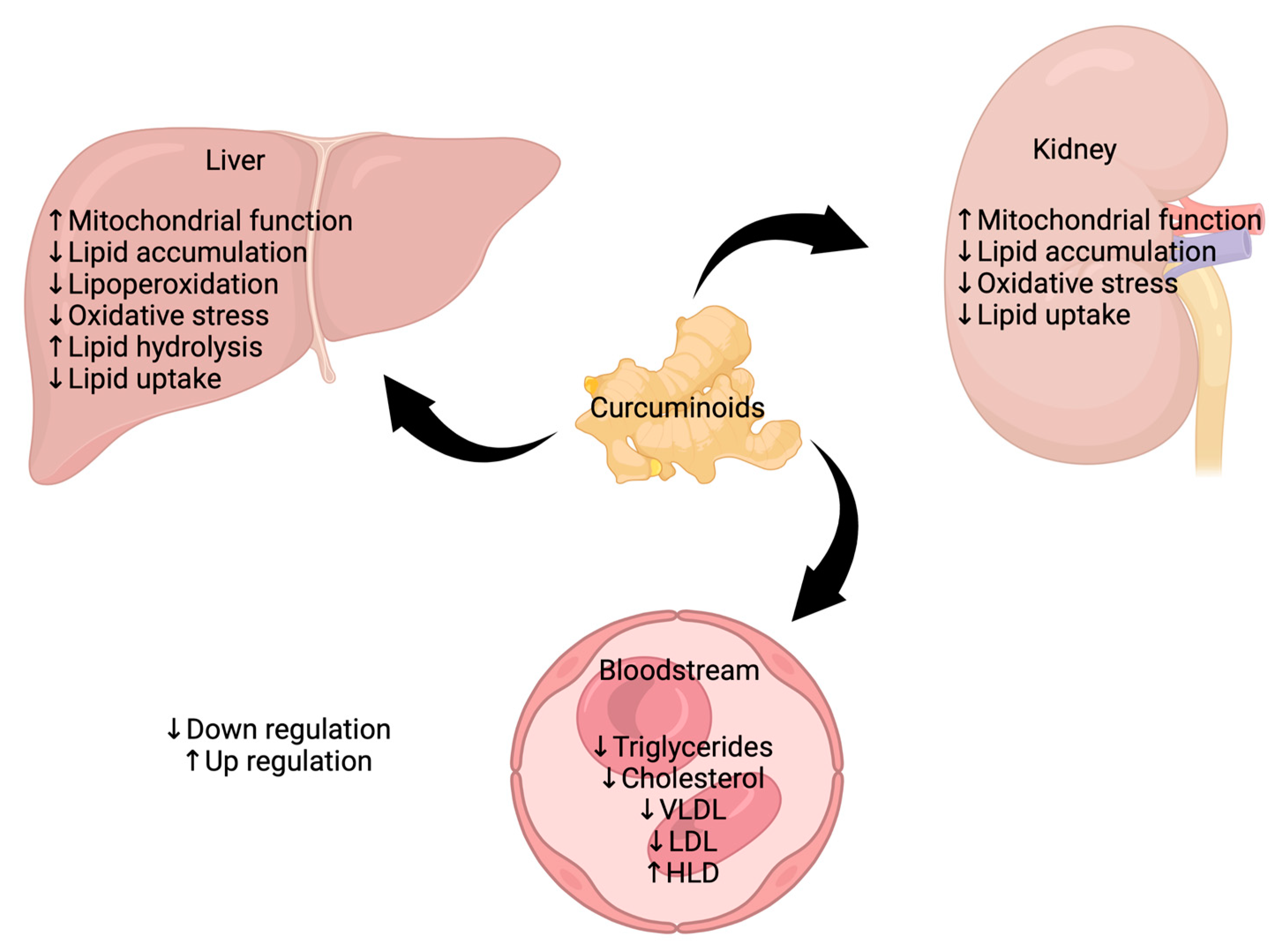
6.3. Effects of Curcuminoids on CKD and Associated Cardiovascular Damage
| Reference | Model | Compound | Dosage | Effect |
|---|---|---|---|---|
| [138] | Sprague–Dawley rats, renal injury induced by 0.25% adenine | Curcumin | 75 mg/kg | Prevents increased creatinine, proteinuria, and BUN levels in CKD during exercise. Prevents the increase in systolic blood pressure and increases the activity of antioxidant enzymes. Decreases fibrosis and inflammation. |
| [140] | C57Bl/6 mice with 5/6 nephrectomy and western diet | Curcumin | 100 mg/kg | Reduces the urine albumin-creatinine ratio and decreases arteriosclerotic lesions. Improve glucose tolerance, and decrease inflammation and blood pressure. |
| [142] | 5/6 nephrectomized Sprague–Dawley rats | Curcumin | 150 mg/kg | Attenuates cardiac hypertrophy and remodeling through the reduction in pGSK-3ß, ß-catenin, and NFAT levels. |
| [160] | 5/6 nephrectomized Sprague–Dawley rats | Theracurmin® | 100 mg/kg/day | Improves ventricular function and avoids heart hypertrophy and interstitial fibrosis by reducing ß-MHC and collagen type 1 |
| [161] | 5/6 nephrectomized Wistar rats | Curcumin | 120 mg/kg | Decreases principal CKD biochemical markers. Prevents ventricular hypertrophy and decreases ischemic events and oxidative stress in heart tissue. |
| [162] | 5/6 nephrectomized Wistar rats | Curcumin | 120 mg/kg | Prevents the tissular remodeling process reducing MMP-2, the activity of gelatinase, and the activation of the IP3K/AKT/ERK signaling pathway |
6.4. Regulation of Dyslipidemia by Curcuminoids in CKD
| Reference | Model | Compound | Dosage | Effect |
|---|---|---|---|---|
| [173] | 5/6 nephrectomized Sprague–Dawley rats | Curcumin | 75 mg/kg | Decreases LDL, total cholesterol, and total triglycerides |
| [174] | Sprague–Dawley rats, renal injury induced by 0.25% adenine | Curcumin | 100 mg/kg | HDL cholesterol increases and decreases total cholesterol, triglycerides, LDL cholesterol, VLDL, NEFA, atherogenic index, and the coronary risk index. In the liver, it increases cholesterol and decreases triglycerides and NEFA. |
| [176] | Meta-analysis | _ | _ | Reduces total cholesterol and TNF-α. Not confirm significant changes in triglyceride, LDL-cholesterol, HDL-cholesterol, and CRP. |
| [177] | Clinical trial, patients with nondiabetic proteinuria CKD | Curcumin | 320 mg/day | Attenuates lipid peroxidation and enhances the antioxidant capacity. |
| [172] | C57BL/6J mice, renal injury induced by a high-fat diet | Bisdemethoxycurcumin | 20 and 40 mg/kg | Decrease renal injury markers, inflammatory cytokines, and tissue fibrosis. Decreases body and white adipose weight, serum glucose, insulin, TC, TG, and HDL-C levels. Increases antioxidant activity and decreases lipid accumulation through Keap1/Nrf2. |
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lv, J.-C.; Zhang, L.-X. Prevalence and Disease Burden of Chronic Kidney Disease. Adv. Exp. Med. Biol. 2019, 1165, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Jager, K.J.; Kovesdy, C.; Langham, R.; Rosenberg, M.; Jha, V.; Zoccali, C. A Single Number for Advocacy and Communication-Worldwide More than 850 Million Individuals Have Kidney Diseases. Nephrol. Dial. Transpl. 2019, 34, 1803–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gansevoort, R.T.; Correa-Rotter, R.; Hemmelgarn, B.R.; Jafar, T.H.; Heerspink, H.J.L.; Mann, J.F.; Matsushita, K.; Wen, C.P. Chronic Kidney Disease and Cardiovascular Risk: Epidemiology, Mechanisms, and Prevention. Lancet 2013, 382, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Sud, M.; Tangri, N.; Pintilie, M.; Levey, A.S.; Naimark, D. Risk of End-Stage Renal Disease and Death after Cardiovascular Events in Chronic Kidney Disease. Circulation 2014, 130, 458–465. [Google Scholar] [CrossRef] [Green Version]
- Schefold, J.C.; Filippatos, G.; Hasenfuss, G.; Anker, S.D.; von Haehling, S. Heart Failure and Kidney Dysfunction: Epidemiology, Mechanisms and Management. Nat. Rev. Nephrol. 2016, 12, 610–623. [Google Scholar] [CrossRef]
- Mittalhenkle, A.; Stehman-Breen, C.O.; Shlipak, M.G.; Fried, L.F.; Katz, R.; Young, B.A.; Seliger, S.; Gillen, D.; Newman, A.B.; Psaty, B.M.; et al. Cardiovascular Risk Factors and Incident Acute Renal Failure in Older Adults: The Cardiovascular Health Study. Clin. J. Am. Soc. Nephrol. 2008, 3, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Vallianou, N.G.; Mitesh, S.; Gkogkou, A.; Geladari, E. Chronic Kidney Disease and Cardiovascular Disease: Is There Any Relationship? Curr. Cardiol. Rev. 2019, 15, 55–63. [Google Scholar] [CrossRef]
- Roubille, F.; Morena, M.; Leray-Moragues, H.; Canaud, B.; Cristol, J.-P.; Klouche, K. Pharmacologic Therapies for Chronic and Acute Decompensated Heart Failure: Specific Insights on Cardiorenal Syndromes. Blood Purif. 2014, 37 (Suppl. S2), 20–33. [Google Scholar] [CrossRef]
- Rucker, D.; Tonelli, M. Cardiovascular Risk and Management in Chronic Kidney Disease. Nat. Rev. Nephrol. 2009, 5, 287–296. [Google Scholar] [CrossRef]
- Johnson, D.W.; Craven, A.-M.; Isbel, N.M. Modification of Cardiovascular Risk in Hemodialysis Patients: An Evidence-Based Review. Hemodial. Int. 2007, 11, 1–14. [Google Scholar] [CrossRef]
- Kocaadam, B.; Şanlier, N. Curcumin, an Active Component of Turmeric (Curcuma longa), and Its Effects on Health. Crit Rev Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef] [PubMed]
- Rysz, J.; Franczyk, B.; Kujawski, K.; Sacewicz-Hofman, I.; Ciałkowska-Rysz, A.; Gluba-Brzózka, A. Are Nutraceuticals Beneficial in Chronic Kidney Disease? Pharmaceutics 2021, 13, 231. [Google Scholar] [CrossRef] [PubMed]
- de Boer, I.H.; Caramori, M.L.; Chan, J.C.N.; Heerspink, H.J.L.; Hurst, C.; Khunti, K.; Liew, A.; Michos, E.D.; Navaneethan, S.D.; Olowu, W.A.; et al. KDIGO 2020 Clinical Practice Guideline for Diabetes Management in Chronic Kidney Disease. Kidney Int. 2020, 98, S1–S115. [Google Scholar] [CrossRef] [PubMed]
- GBD Chronic Kidney Disease Collaboration Global, Regional, and National Burden of Chronic Kidney Disease, 1990-2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [CrossRef] [PubMed] [Green Version]
- Jha, V.; Garcia-Garcia, G.; Iseki, K.; Li, Z.; Naicker, S.; Plattner, B.; Saran, R.; Wang, A.Y.-M.; Yang, C.-W. Chronic Kidney Disease: Global Dimension and Perspectives. Lancet 2013, 382, 260–272. [Google Scholar] [CrossRef]
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef]
- Go, A.S.; Chertow, G.M.; Fan, D.; McCulloch, C.E.; Hsu, C. Chronic Kidney Disease and the Risks of Death, Cardiovascular Events, and Hospitalization. N. Engl. J. Med. 2004, 351, 1296–1305. [Google Scholar] [CrossRef]
- Burst, V.; Benzing, T. Dyslipidemia Treatment and Cardiovascular Disease in the Renal Patient. Curr. Pharm. Des. 2011, 17, 894–907. [Google Scholar] [CrossRef]
- Hager, M.R.; Narla, A.D.; Tannock, L.R. Dyslipidemia in Patients with Chronic Kidney Disease. Rev. Endocr. Metab. Disord. 2017, 18, 29–40. [Google Scholar] [CrossRef]
- Diabetes Canada Clinical Practice Guidelines Expert, Committee; Mancini, G.B.J.; Hegele, R.A.; Leiter, L.A. Dyslipidemia. Can. J. Diabetes 2018, 42 (Suppl. S1), S178–S185. [Google Scholar] [CrossRef] [Green Version]
- Illingworth, D.R. Lipoprotein Metabolism. Am. J. Kidney Dis. 1993, 22, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ortega, M.; Rayego-Mateos, S.; Lamas, S.; Ortiz, A.; Rodrigues-Diez, R.R. Targeting the Progression of Chronic Kidney Disease. Nat. Rev. Nephrol. 2020, 16, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Barter, P. Lipoprotein Metabolism and CKD: Overview. Clin. Exp. Nephrol. 2014, 18, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Go, A.S.; Chandra, M.; Fan, D.; Kaysen, G.A. GFR, Body Mass Index, and Low High-Density Lipoprotein Concentration in Adults with and without CKD. Am. J. Kidney Dis. 2007, 50, 552–558. [Google Scholar] [CrossRef]
- Attman, P.O.; Samuelsson, O.; Alaupovic, P. Lipoprotein Metabolism and Renal Failure. Am. J. Kidney Dis. 1993, 21, 573–592. [Google Scholar] [CrossRef]
- Rosenstein, K.; Tannock, L.R. KDOQI Clinical Practice Guideline for Diabetes and CKD: 2012 Update. Am. J. Kidney Dis. 2012, 60, 850–886. [Google Scholar] [CrossRef]
- Atar, D.; Jukema, J.W.; Molemans, B.; Taub, P.R.; Goto, S.; Mach, F.; CerezoOlmos, C.; Underberg, J.; Keech, A.; Tokgözoğlu, L.; et al. New Cardiovascular Prevention Guidelines: How to Optimally Manage Dyslipidaemia and Cardiovascular Risk in 2021 in Patients Needing Secondary Prevention? Atherosclerosis 2021, 319, 51–61. [Google Scholar] [CrossRef]
- Sadeghi, M.; Haghdoost, A.A.; Bahrampour, A.; Dehghani, M. Modeling the Burden of Cardiovascular Diseases in Iran from 2005 to 2025: The Impact of Demographic Changes. Iran. J. Public Health 2017, 46, 506–516. [Google Scholar]
- Hedayatnia, M.; Asadi, Z.; Zare-Feyzabadi, R.; Yaghooti-Khorasani, M.; Ghazizadeh, H.; Ghaffarian-Zirak, R.; Nosrati-Tirkani, A.; Mohammadi-Bajgiran, M.; Rohban, M.; Sadabadi, F.; et al. Dyslipidemia and Cardiovascular Disease Risk among the MASHAD Study Population. Lipids Health Dis. 2020, 19, 42. [Google Scholar] [CrossRef] [Green Version]
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D.R. Global Prevalence of Chronic Kidney Disease—A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [Green Version]
- Kottgen, A.; Russell, S.D.; Loehr, L.R.; Crainiceanu, C.M.; Rosamond, W.D.; Chang, P.P.; Chambless, L.E.; Coresh, J. Reduced Kidney Function as a Risk Factor for Incident Heart Failure: The Atherosclerosis Risk in Communities (ARIC) Study. J. Am. Soc. Nephrol. 2007, 18, 1307–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalita, S.; Khandelwal, S.; Madan, J.; Pandya, H.; Sesikeran, B.; Krishnaswamy, K. Almonds and Cardiovascular Health: A Review. Nutrients 2018, 10, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostom, A.G.; Cupples, L.A.; Jenner, J.L.; Ordovas, J.M.; Seman, L.J.; Wilson, P.W.; Schaefer, E.J.; Castelli, W.P. Elevated Plasma Lipoprotein(a) and Coronary Heart Disease in Men Aged 55 Years and Younger. A Prospective Study. JAMA 1996, 276, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Abbas Torki, S.; Bahadori, E.; Shekari, S.; Fathi, S.; Gholamalizadeh, M.; Hasanpour Ardekanizadeh, N.; Aminnezhad, B.; Ahmadzadeh, M.; Sotoudeh, M.; Shafie, F.; et al. Association between the Index of Nutritional Quality and Lipid Profile in Adult Women. Endocrinol. Diabetes Metab. 2022, 5, e358. [Google Scholar] [CrossRef] [PubMed]
- Després, J.-P.; Lemieux, I.; Dagenais, G.-R.; Cantin, B.; Lamarche, B. HDL-Cholesterol as a Marker of Coronary Heart Disease Risk: The Québec Cardiovascular Study. Atherosclerosis 2000, 153, 263–272. [Google Scholar] [CrossRef]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef]
- Hill, M.F.; Bordoni, B. Hyperlipidemia; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- LUCCHI, T. Dyslipidemia and Prevention of Atherosclerotic Cardiovascular Disease in the Elderly. Minerva Med. 2022, 112. [Google Scholar] [CrossRef]
- Kones, R. Molecular Sources of Residual Cardiovascular Risk, Clinical Signals, and Innovative Solutions: Relationship with Subclinical Disease, Undertreatment, and Poor Adherence: Implications of New Evidence upon Optimizing Cardiovascular Patient Outcomes. Vasc. Health Risk Manag. 2013, 617. [Google Scholar] [CrossRef] [Green Version]
- Cholesterol Treatment Trialists’ (CTT) Collaboration; Fulcher, J.; O’Connell, R.; Voysey, M.; Emberson, J.; Blackwell, L.; Mihaylova, B.; Simes, J.; Collins, R.; Kirby, A.; et al. Efficacy and Safety of LDL-Lowering Therapy among Men and Women: Meta-Analysis of Individual Data from 174,000 Participants in 27 Randomised Trials. Lancet 2015, 385, 1397–1405. [Google Scholar] [CrossRef]
- Pirillo, A.; Casula, M.; Olmastroni, E.; Norata, G.D.; Catapano, A.L. Global Epidemiology of Dyslipidaemias. Nat. Rev. Cardiol. 2021, 18, 689–700. [Google Scholar] [CrossRef]
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgözoğlu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Prim. 2019, 5, 56. [Google Scholar] [CrossRef]
- Jankowski, J.; Floege, J.; Fliser, D.; Böhm, M.; Marx, N. Cardiovascular Disease in Chronic Kidney Disease: Pathophysiological Insights and Therapeutic Options. Circulation 2021, 143, 1157–1172. [Google Scholar] [CrossRef] [PubMed]
- Tsuruya, K.; Eriguchi, M. Cardiorenal Syndrome in Chronic Kidney Disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Wali, R.K.; Wang, G.S.; Gottlieb, S.S.; Bellumkonda, L.; Hansalia, R.; Ramos, E.; Drachenberg, C.; Papadimitriou, J.; Brisco, M.A.; Blahut, S.; et al. Effect of Kidney Transplantation on Left Ventricular Systolic Dysfunction and Congestive Heart Failure in Patients with End-Stage Renal Disease. J. Am. Coll. Cardiol. 2005, 45, 1051–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziani, G.; Pini, D.; Oldani, S.; Cucchiari, D.; Podestà, M.A.; Badalamenti, S. Renal Dysfunction in Acute Congestive Heart Failure: A Common Problem for Cardiologists and Nephrologists. Heart Fail. Rev. 2014, 19, 699–708. [Google Scholar] [CrossRef]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial Energetics in the Kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef]
- Chen, C.-M.; Chung, Y.-P.; Liu, C.-H.; Huang, K.-T.; Guan, S.-S.; Chiang, C.-K.; Wu, C.-T.; Liu, S.-H. Withaferin A Protects against Endoplasmic Reticulum Stress-Associated Apoptosis, Inflammation, and Fibrosis in the Kidney of a Mouse Model of Unilateral Ureteral Obstruction. Phytomedicine 2020, 79, 153352. [Google Scholar] [CrossRef]
- Holmström, K.M.; Kostov, R.V.; Dinkova-Kostova, A.T. The Multifaceted Role of Nrf2 in Mitochondrial Function. Curr. Opin. Toxicol. 2016, 1, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.D.; Xu, S.; Johns, D.G.; Du, Y.; Quinn, M.T.; Cayatte, A.J.; Cohen, R.A. Role of NADPH Oxidase in the Vascular Hypertrophic and Oxidative Stress Response to Angiotensin II in Mice. Circ. Res. 2001, 88, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Zhang, L.; Brett-Morris, A.; Aguila, B.; Kerner, J.; Hoppel, C.L.; Puchowicz, M.; Serra, D.; Herrero, L.; Rini, B.I.; et al. HIF Drives Lipid Deposition and Cancer in CcRCC via Repression of Fatty Acid Metabolism. Nat. Commun. 2017, 8, 1769. [Google Scholar] [CrossRef] [Green Version]
- Fontecha-Barriuso, M.; Martin-Sanchez, D.; Martinez-Moreno, J.; Monsalve, M.; Ramos, A.; Sanchez-Niño, M.; Ruiz-Ortega, M.; Ortiz, A.; Sanz, A. The Role of PGC-1α and Mitochondrial Biogenesis in Kidney Diseases. Biomolecules 2020, 10, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulianiti, K.P.; Kaltsatou, A.; Mitrou, G.I.; Jamurtas, A.Z.; Koutedakis, Y.; Maridaki, M.; Stefanidis, I.; Sakkas, G.K.; Karatzaferi, C. Systemic Redox Imbalance in Chronic Kidney Disease: A Systematic Review. Oxid. Med. Cell. Longev. 2016, 2016, 8598253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi Tosi, M.E.; Bocanegra, V.; Manucha, W.; Gil Lorenzo, A.; Vallés, P.G. The Nrf2-Keap1 Cellular Defense Pathway and Heat Shock Protein 70 (Hsp70) Response. Role in Protection against Oxidative Stress in Early Neonatal Unilateral Ureteral Obstruction (UUO). Cell Stress Chaperones 2011, 16, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Guo, H.; Li, J.; Ma, T.; Zhou, S.; Zhang, Z.; Miao, L.; Cai, L. Sulforaphane Prevents Type 2 Diabetes-Induced Nephropathy via AMPK-Mediated Activation of Lipid Metabolic Pathways and Nrf2 Antioxidative Function. Clin. Sci. (Lond.) 2020, 134, 2469–2487. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Aparicio-Trejo, O.E.; Tapia, E.; Sánchez-Lozada, L.G.; García-Arroyo, F.E.; Amador-Martínez, I.; Orozco-Ibarra, M.; Fernández-Valverde, F.; Pedraza-Chaverri, J. Sulforaphane Protects against Unilateral Ureteral Obstruction-Induced Renal Damage in Rats by Alleviating Mitochondrial and Lipid Metabolism Impairment. Antioxidants 2022, 11, 1854. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Pedraza-Chaverri, J.; Scholze, A. Nrf2 Activation in Chronic Kidney Disease: Promises and Pitfalls. Antioxidants 2022, 11, 1112. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Itoh, K.; Ruiz, E.; Leake, D.S.; Unoki, H.; Yamamoto, M.; Mann, G.E. Role of Nrf2 in the Regulation of CD36 and Stress Protein Expression in Murine Macrophages: Activation by Oxidatively Modified LDL and 4-Hydroxynonenal. Circ. Res. 2004, 94, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Clarke, R.; Hammami, I.; Sherliker, P.; Valdes-Marquez, E.; Watkins, H.; Hill, M.; Yang, X.; Tsimikas, S.; Hopewell, J.C. Oxidized Phospholipids on Apolipoprotein B-100 versus Plasminogen and Risk of Coronary Heart Disease in the PROCARDIS Study. Atherosclerosis 2022, 354, 15–22. [Google Scholar] [CrossRef]
- Kon, V.; Linton, M.F.; Fazio, S. Atherosclerosis in Chronic Kidney Disease: The Role of Macrophages. Nat. Rev. Nephrol. 2011, 7, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.-M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression, and Outcome. J. Immunol Res. 2018, 2018, 2180373. [Google Scholar] [CrossRef] [Green Version]
- Zeisberg, M.; Kalluri, R. Cellular Mechanisms of Tissue Fibrosis. 1. Common and Organ-Specific Mechanisms Associated with Tissue Fibrosis. Am. J. Physiol. Cell Physiol. 2013, 304, C216–C225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Klimova, E.; Aparicio-Trejo, O.E.; Tapia, E.; Pedraza-Chaverri, J. Unilateral Ureteral Obstruction as a Model to Investigate Fibrosis-Attenuating Treatments. Biomolecules 2019, 9, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Guo, Y.; Zhang, Y.; Zhang, X.; Zhu, L.; Yan, T. Salidroside Ameliorates Renal Interstitial Fibrosis by Inhibiting the TLR4/NF-ΚB and MAPK Signaling Pathways. Int. J. Mol. Sci. 2019, 20, 1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, P.M.-K.; Nikolic-Paterson, D.J.; Lan, H.-Y. Macrophages: Versatile Players in Renal Inflammation and Fibrosis. Nat. Rev. Nephrol. 2019, 15, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Love-Gregory, L.; Sherva, R.; Sun, L.; Wasson, J.; Schappe, T.; Doria, A.; Rao, D.C.; Hunt, S.C.; Klein, S.; Neuman, R.J.; et al. Variants in the CD36 Gene Associate with the Metabolic Syndrome and High-Density Lipoprotein Cholesterol. Hum. Mol. Genet. 2008, 17, 1695–1704. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.C.P.; Bocharov, A.V.; Baranova, I.N.; Vishnyakova, T.G.; Huang, Y.G.; Wilkins, K.J.; Hu, X.; Street, J.M.; Alvarez-Prats, A.; Mullick, A.E.; et al. Antagonism of Scavenger Receptor CD36 by 5A Peptide Prevents Chronic Kidney Disease Progression in Mice Independent of Blood Pressure Regulation. Kidney Int. 2016, 89, 809–822. [Google Scholar] [CrossRef] [Green Version]
- Hossain, E.; Ota, A.; Karnan, S.; Takahashi, M.; Mannan, S.B.; Konishi, H.; Hosokawa, Y. Lipopolysaccharide Augments the Uptake of Oxidized LDL by Up-Regulating Lectin-like Oxidized LDL Receptor-1 in Macrophages. Mol. Cell. Biochem. 2015, 400, 29–40. [Google Scholar] [CrossRef]
- Liu, W.; Yin, Y.; Zhou, Z.; He, M.; Dai, Y. OxLDL-Induced IL-1beta Secretion Promoting Foam Cells Formation Was Mainly via CD36 Mediated ROS Production Leading to NLRP3 Inflammasome Activation. Inflamm. Res. 2014, 63, 33–43. [Google Scholar] [CrossRef]
- Okamura, D.M.; Pennathur, S.; Pasichnyk, K.; López-Guisa, J.M.; Collins, S.; Febbraio, M.; Heinecke, J.; Eddy, A.A. CD36 Regulates Oxidative Stress and Inflammation in Hypercholesterolemic CKD. J. Am. Soc. Nephrol. 2009, 20, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Himmelfarb, J. Linking Oxidative Stress and Inflammation in Kidney Disease: Which Is the Chicken and Which Is the Egg? Semin Dial. 2004, 17, 449–454. [Google Scholar] [CrossRef]
- Geloen, A.; Helin, L.; Geeraert, B.; Malaud, E.; Holvoet, P.; Marguerie, G. CD36 Inhibitors Reduce Postprandial Hypertriglyceridemia and Protect against Diabetic Dyslipidemia and Atherosclerosis. PLoS ONE 2012, 7, e37633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedigo, C.E.; Ducasa, G.M.; Leclercq, F.; Sloan, A.; Mitrofanova, A.; Hashmi, T.; Molina-David, J.; Ge, M.; Lassenius, M.I.; Forsblom, C.; et al. Local TNF Causes NFATc1-Dependent Cholesterol-Mediated Podocyte Injury. J. Clin. Investig. 2016, 126, 3336–3350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Andres, O.; Suarez-Alvarez, B.; Sánchez-Ramos, C.; Monsalve, M.; Sanchez-Niño, M.D.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A.; Sanz, A.B. The Inflammatory Cytokine TWEAK Decreases PGC-1α Expression and Mitochondrial Function in Acute Kidney Injury. Kidney Int. 2016, 89, 399–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio-Trejo, O.E.; Tapia, E.; Sánchez-Lozada, L.G.; Pedraza-Chaverri, J. Mitochondrial Bioenergetics, Redox State, Dynamics and Turnover Alterations in Renal Mass Reduction Models of Chronic Kidney Diseases and Their Possible Implications in the Progression of This Illness. Pharm. Res. 2018, 135, 1–11. [Google Scholar] [CrossRef]
- Martínez-Klimova, E.; Aparicio-Trejo, O.E.; Gómez-Sierra, T.; Jiménez-Uribe, A.P.; Bellido, B.; Pedraza-Chaverri, J. Mitochondrial Dysfunction and Endoplasmic Reticulum Stress in the Promotion of Fibrosis in Obstructive Nephropathy Induced by Unilateral Ureteral Obstruction. BioFactors 2020, 46, 716–733. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, W.; Rittinger, M. Quantitative Morphology of the Rat Kidney. Int. J. Biochem. 1980, 12, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.M.; Rhodes, G.J.; Sandoval, R.M.; Corridon, P.R.; Molitoris, B.A. In Vivo Multiphoton Imaging of Mitochondrial Structure and Function during Acute Kidney Injury. Kidney Int. 2014, 83, 72–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A.M.; Unwin, R.J.; Parker, N.; Duchen, M.R. Multiphoton Imaging Reveals Differences in Mitochondrial Function between Nephron Segments. J. Am. Soc. Nephrol. 2009, 20, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, M.; Dubach, U.C. Peroxisomal and Mitochondrial Beta-Oxidation in the Rat Kidney. Distribution of Fatty Acyl-Coenzyme A Oxidase and 3-Hydroxyacyl-Coenzyme A Dehydrogenase Activities along the Nephron. J. Histochem. Cytochem. 1982, 30, 441–444. [Google Scholar] [CrossRef] [Green Version]
- Sekine, T.; Endou, H. Solute Transport, Energy Consumption, and Production in the Kidney. In Seldin and Geibisch’s the Kidney; Elsevier: New York, NY, USA, 2013; Volume 1, pp. 143–175. ISBN 9780123814623. [Google Scholar]
- Goncalves, R.L.S.; Quinlan, C.L.; Perevoshchikova, I.V.; Hey-Mogensen, M.; Brand, M.D. Sites of Superoxide and Hydrogen Peroxide Production by Muscle Mitochondria Assessed Ex Vivo under Conditions Mimicking Rest and Exercise. J. Biol. Chem. 2015, 290, 209–227. [Google Scholar] [CrossRef] [Green Version]
- Klein, K.; Maw-Song, W.; Torikai, S.; Warren, D.D.; Kurakawa, K. Substrate Oxidation by Isolated Single Nephron Segments of the Rat. Kidney Int. 1981, 20, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio-Trejo, O.E.; Aranda-Rivera, A.K.; Osorio-Alonso, H.; Martínez-Klimova, E.; Sánchez-Lozada, L.G.; Pedraza-Chaverri, J.; Tapia, E. Extracellular Vesicles in Redox Signaling and Metabolic Regulation in Chronic Kidney Disease. Antioxidants 2022, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Jiang, L.; Xu, J.; Bai, F.; Zhou, Y.; Yuan, Q.; Luo, J.; Zen, K.; Yang, J. Inhibiting Aerobic Glycolysis Suppresses Renal Interstitial Fibroblast Activation and Renal Fibrosis. Am. J. Physiol. Ren. Physiol. 2017, 313, F561–F575. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Su, J.; Dong, G.; Zhang, M.; Huo, Y.; Dong, Z. Glycolysis Inhibitors Suppress Renal Interstitial Fibrosis via Divergent Effects on Fibroblasts and Tubular Cells. Am. J. Physiol. Ren. Physiol. 2019, 316, F1162–F1172. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Thorburn, D.R. Mitochondrial Dysfunction in Diabetic Kidney Disease. Nat. Rev. Nephrol. 2018, 14, 291–312. [Google Scholar] [CrossRef] [PubMed]
- Caron, A.; Richard, D.; Laplante, M. The Roles of MTOR Complexes in Lipid Metabolism. Annu. Rev. Nutr. 2015, 35, 321–348. [Google Scholar] [CrossRef] [PubMed]
- Norman, J.T.; Hwang, S.; Fine, L.G.; Nephrology, D.; Bradley, T.; Am, J. Hypertrophy of Renal Mitochondria. J. Am. Soc. Nephrol. 1990, I, 822–827. [Google Scholar] [CrossRef]
- Aparicio-Trejo, O.E.; Tapia, E.; Molina-Jijón, E.; Medina-Campos, O.N.; Macías-Ruvalcaba, N.A.; León-Contreras, J.C.; Hernández-Pando, R.; García-Arroyo, F.E.; Cristóbal, M.; Sánchez-Lozada, L.G.; et al. Curcumin Prevents Mitochondrial Dynamics Disturbances in Early 5/6 Nephrectomy: Relation to Oxidative Stress and Mitochondrial Bioenergetics. BioFactors 2017, 43, 293–310. [Google Scholar] [CrossRef]
- Tannenbaum, J.; Purkerson, M.L.; Klahr, S. Effect of Unilateral Ureteral Obstruction on Metabolism of Renal Lipids in the Rat. Am. J. Physiol. Ren. Fluid Electrolyte Physiol. 1983, 14, F254–F262. [Google Scholar] [CrossRef]
- Kang, H.M.; Ahn, S.H.; Choi, P.; Ko, Y.A.; Han, S.H.; Chinga, F.; Park, A.S.D.; Tao, J.; Sharma, K.; Pullman, J.; et al. Defective Fatty Acid Oxidation in Renal Tubular Epithelial Cells Has a Key Role in Kidney Fibrosis Development. Nat. Med. 2015, 21, 37–46. [Google Scholar] [CrossRef]
- Jang, H.S.; Noh, M.R.; Kim, J.; Padanilam, B.J. Defective Mitochondrial Fatty Acid Oxidation and Lipotoxicity in Kidney Diseases. Front. Med. (Lausanne) 2020, 7, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio-Trejo, O.E.; Rojas-Morales, P.; Avila-Rojas, S.H.; León-Contreras, J.C.; Hernández-Pando, R.; Jiménez-Uribe, A.P.; Prieto-Carrasco, R.; Sánchez-Lozada, L.G.; Pedraza-Chaverri, J.; Tapia, E. Temporal Alterations in Mitochondrial β-Oxidation and Oxidative Stress Aggravate Chronic Kidney Disease Development in 5/6 Nephrectomy Induced Renal Damage. Int. J. Mol. Sci. 2020, 21, 6512. [Google Scholar] [CrossRef] [PubMed]
- Afshinnia, F.; Rajendiran, T.M.; Soni, T.; Byun, J.; Wernisch, S.; Sas, K.M.; Hawkins, J.; Bellovich, K.; Gipson, D.; Michailidis, G.; et al. Impaired β-Oxidation and Altered Complex Lipid Fatty Acid Partitioning with Advancing CKD. J. Am. Soc. Nephrol. 2018, 29, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceja-Galicia, Z.A.; García-Arroyo, F.E.; Aparicio-Trejo, O.E.; El-Hafidi, M.; Gonzaga-Sánchez, G.; León-Contreras, J.C.; Hernández-Pando, R.; Guevara-Cruz, M.; Tovar, A.R.; Rojas-Morales, P.; et al. Therapeutic Effect of Curcumin on 5/6Nx Hypertriglyceridemia: Association with the Improvement of Renal Mitochondrial β-Oxidation and Lipid Metabolism in Kidney and Liver. Antioxidant 2022, 11, 2195. [Google Scholar] [CrossRef]
- Gai, Z.; Wang, T.; Visentin, M.; Kullak-Ublick, G.A.; Fu, X.; Wang, Z. Lipid Accumulation and Chronic Kidney Disease. Nutrients 2019, 11, 722. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Hirano, T.; Nagashima, M.; Shiraishi, Y.; Fukui, T.; Adachi, M. Decreased Peroxisome Proliferator-Activated Receptor Alpha Gene Expression Is Associated with Dyslipidemia in a Rat Model of Chronic Renal Failure. Metabolism 2007, 56, 1714–1718. [Google Scholar] [CrossRef]
- Jin, K.; Norris, K.; Vaziri, N.D. Dysregulation of Hepatic Fatty Acid Metabolism in Chronic Kidney Disease. Nephrol. Dial. Transpl. 2013, 28, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.K.; Knicely, D.H.; Grams, M.E. Chronic Kidney Disease Diagnosis and Management. JAMA 2019, 322, 1294. [Google Scholar] [CrossRef]
- Leon, S.J.; Whitlock, R.; Rigatto, C.; Komenda, P.; Bohm, C.; Sucha, E.; Bota, S.E.; Tuna, M.; Collister, D.; Sood, M.; et al. Hyperkalemia-Related Discontinuation of Renin-Angiotensin-Aldosterone System Inhibitors and Clinical Outcomes in CKD: A Population-Based Cohort Study. Am. J. Kidney Dis. 2022. [Google Scholar] [CrossRef]
- Felker, G.M.; Lee, K.L.; Bull, D.A.; Redfield, M.M.; Stevenson, L.W.; Goldsmith, S.R.; LeWinter, M.M.; Deswal, A.; Rouleau, J.L.; Ofili, E.O.; et al. Diuretic Strategies in Patients with Acute Decompensated Heart Failure. N. Engl. J. Med. 2011, 364, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.; Bettencourt, P.; Dias, P.; Pestana, M.; Serrão, P.; Soares-da-Silva, P.; Cerqueira-Gomes, M. Neurohormonal Activation, the Renal Dopaminergic System and Sodium Handling in Patients with Severe Heart Failure under Vasodilator Therapy. Clin. Sci. (Lond.) 2001, 100, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Stancu, C.; Sima, A. Statins: Mechanism of Action and Effects. J. Cell. Mol. Med. 2001, 5, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Pahan, K. Lipid-Lowering Drugs. Cell. Mol. Life Sci. 2006, 63, 1165–1178. [Google Scholar] [CrossRef] [PubMed]
- Drexel, H. Statins, Fibrates, Nicotinic Acid, Cholesterol Absorption Inhibitors, Anion-Exchange Resins, Omega-3 Fatty Acids: Which Drugs for Which Patients? Fundam Clin. Pharm. 2009, 23, 687–692. [Google Scholar] [CrossRef]
- Shek, A.; Ferrill, M.J. Statin-Fibrate Combination Therapy. Ann. Pharm. 2001, 35, 908–917. [Google Scholar] [CrossRef]
- Chauhan, D.; Memon, F.; Patwardhan, V.; Kotwani, P.; Shah, P.; Samala Venkata, V. Comparing Simvastatin Monotherapy V/S Simvastatin-Ezetimibe Combination Therapy for the Treatment of Hyperlipidemia: A Meta-Analysis and Review. Cureus 2022, 14, e31007. [Google Scholar] [CrossRef]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M. Curcumin and Health. Molecules 2016, 21, 264. [Google Scholar] [CrossRef]
- Lestari, M.L.A.D.; Indrayanto, G. Curcumin. Profiles Drug Subst. Excip. Relat. Methodol. 2014, 39, 113–204. [Google Scholar] [CrossRef]
- Zhang, H.; Jiang, X.; Song, W.; Yang, Z. Novel Synthetic Curcuminoids: Not Merely an Anticancer Agent. Pak. J. Pharm Sci 2019, 32. [Google Scholar]
- Stohs, S.J.; Chen, O.; Ray, S.D.; Ji, J.; Bucci, L.R.; Preuss, H.G. Highly Bioavailable Forms of Curcumin and Promising Avenues for Curcumin-Based Research and Application: A Review. Molecules 2020, 25, 1397. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.H.; Huang, T.M.; Lin, J.K. Biotransformation of Curcumin through Reduction and Glucuronidation in Mice. Drug Metab. Dispos. 1999, 27, 486–494. [Google Scholar] [PubMed]
- Park, S.J.; Garcia, C.V.; Shin, G.H.; Kim, J.T. Improvement of Curcuminoid Bioaccessibility from Turmeric by a Nanostructured Lipid Carrier System. Food Chem. 2018, 251, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From Kitchen to Clinic. Biochem. Pharm. 2008, 75, 787–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.; Lou, H.; Zhao, L.; Fan, P. Validated LC/MS/MS Assay for Curcumin and Tetrahydrocurcumin in Rat Plasma and Application to Pharmacokinetic Study of Phospholipid Complex of Curcumin. J. Pharm. Biomed. Anal. 2006, 40, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Tønnesen, H.H.; Másson, M.; Loftsson, T. Studies of Curcumin and Curcuminoids. XXVII. Cyclodextrin Complexation: Solubility, Chemical and Photochemical Stability. Int. J. Pharm. 2002, 244, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Maiti, K.; Mukherjee, K.; Gantait, A.; Saha, B.P.; Mukherjee, P.K. Curcumin–Phospholipid Complex: Preparation, Therapeutic Evaluation and Pharmacokinetic Study in Rats. Int. J. Pharm. 2007, 330, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Shin, G.H.; Chung, S.K.; Kim, J.T.; Joung, H.J.; Park, H.J. Preparation of Chitosan-Coated Nanoliposomes for Improving the Mucoadhesive Property of Curcumin Using the Ethanol Injection Method. J. Agric. Food Chem. 2013, 61, 11119–11126. [Google Scholar] [CrossRef]
- Hu, Q.; Luo, Y. Chitosan-Based Nanocarriers for Encapsulation and Delivery of Curcumin: A Review. Int. J. Biol. Macromol. 2021, 179, 125–135. [Google Scholar] [CrossRef]
- Hansapaiboon, S.; Bulatao, B.P.; Sorasitthiyanukarn, F.N.; Jantaratana, P.; Nalinratana, N.; Vajragupta, O.; Rojsitthisak, P.; Rojsitthisak, P. Fabrication of Curcumin Diethyl γ-Aminobutyrate-Loaded Chitosan-Coated Magnetic Nanocarriers for Improvement of Cytotoxicity against Breast Cancer Cells. Polymers 2022, 14, 5563. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Y.; Wang, Y.-W.; Huang, M.-T.; Ho, C.-T.; Huang, Q. Enhancing Anti-Inflammation Activity of Curcumin through O/W Nanoemulsions. Food Chem. 2008, 108, 419–424. [Google Scholar] [CrossRef]
- Lopresti, A.L. The Problem of Curcumin and Its Bioavailability: Could Its Gastrointestinal Influence Contribute to Its Overall Health-Enhancing Effects? Adv. Nutr. 2018, 9, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Saud, N.B.S. Impact of Curcumin Treatment on Diabetic Albino Rats. Saudi J. Biol. Sci. 2020, 27, 689–694. [Google Scholar] [CrossRef]
- Lima, T.F.O.; Costa, M.C.; Figueiredo, I.D.; Inácio, M.D.; Rodrigues, M.R.; Assis, R.P.; Baviera, A.M.; Brunetti, I.L. Curcumin, Alone or in Combination with Aminoguanidine, Increases Antioxidant Defenses and Glycation Product Detoxification in Streptozotocin-Diabetic Rats: A Therapeutic Strategy to Mitigate Glycoxidative Stress. Oxid. Med. Cell. Longev. 2020, 2020, 1036360. [Google Scholar] [CrossRef] [PubMed]
- Salarolli, R.T.; Alvarenga, L.; Cardozo, L.F.M.F.; Teixeira, K.T.R.; de SG Moreira, L.; Lima, J.D.; Rodrigues, S.D.; Nakao, L.S.; Fouque, D.; Mafra, D. Can Curcumin Supplementation Reduce Plasma Levels of Gut-Derived Uremic Toxins in Hemodialysis Patients? A Pilot Randomized, Double-Blind, Controlled Study. Int. Urol. Nephrol. 2021, 53, 1231–1238. [Google Scholar] [CrossRef]
- Hami, M.; Bigdeli, A.; Khameneh Bagheri, R.; Rajabi, O.; Salehi, M.; Zahedi Avval, F. The Effect of Curcumin in Prevention of Contrast Nephropathy Following Coronary Angiography or Angioplasty in CKD Patients. Iran. J. Kidney Dis. 2019, 13, 304–309. [Google Scholar] [PubMed]
- Cas, M.D.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef] [Green Version]
- Kindgen-Milles, D.; Slowinski, T.; Dimski, T. New Kidney Function Tests: Renal Functional Reserve and Furosemide Stress Test. Med. Klin. Intensiv. Notfmed 2020, 115, 37–42. [Google Scholar] [CrossRef]
- Tapia, E.; Zatarain-Barrón, Z.L.; Hernández-Pando, R.; Zarco-Márquez, G.; Molina-Jijón, E.; Cristóbal-García, M.; Santamaría, J.; Pedraza-Chaverri, J. Curcumin Reverses Glomerular Hemodynamic Alterations and Oxidant Stress in 5/6 Nephrectomized Rats. Phytomedicine 2013, 20, 359–366. [Google Scholar] [CrossRef]
- Tapia, E.; Soto, V.; Ortiz-Vega, K.M.; Zarco-Márquez, G.; Molina-Jijón, E.; Cristóbal-García, M.; Santamaría, J.; García-Niño, W.R.; Correa, F.; Zazueta, C.; et al. Curcumin Induces Nrf2 Nuclear Translocation and Prevents Glomerular Hypertension, Hyperfiltration, Oxidant Stress, and the Decrease in Antioxidant Enzymes in 5/6 Nephrectomized Rats. Oxid. Med. Cell. Longev. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Tapia, E.; García-Arroyo, F.; Silverio, O.; Rodríguez-Alcocer, A.N.; Jiménez-Flores, A.B.; Cristobal, M.; Arellano, A.S.; Soto, V.; Osorio-Alonso, H.; Molina-Jijón, E.; et al. Mycophenolate Mofetil and Curcumin Provide Comparable Therapeutic Benefit in Experimental Chronic Kidney Disease: Role of Nrf2-Keap1 and Renal Dopamine Pathways. Free Radic. Res. 2016, 50, 781–792. [Google Scholar] [CrossRef]
- Lau, W.L.; Khazaeli, M.; Savoj, J.; Manekia, K.; Bangash, M.; Thakurta, R.G.; Dang, A.; Vaziri, N.D.; Singh, B. Dietary Tetrahydrocurcumin Reduces Renal Fibrosis and Cardiac Hypertrophy in 5/6 Nephrectomized Rats. Pharm. Res. Perspect. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Al-Salam, S.; al Suleimani, Y.; al Kalbani, J.; al Bahlani, S.; Ashique, M.; Manoj, P.; al Dhahli, B.; al Abri, N.; Naser, H.T.; et al. Curcumin Ameliorates Kidney Function and Oxidative Stress in Experimental Chronic Kidney Disease. Basic Clin. Pharm. Toxicol. 2018, 122, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muta, K.; Obata, Y.; Oka, S.; Abe, S.; Minami, K.; Kitamura, M.; Endo, D.; Koji, T.; Nishino, T. Curcumin Ameliorates Nephrosclerosis via Suppression of Histone Acetylation Independent of Hypertension. Nephrol. Dial. Transplant. 2016, 31, 1615–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, F.; Jin, Y.; Du, J.; Jiang, L.; Zhang, Y.; Zhao, Z.; Yang, B.; Luo, P.; He, Q. Bisdemethoxycurcumin Protects against Renal Fibrosis via Activation of Fibroblast Apoptosis. Eur. J. Pharm. 2019, 847, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Villanego, F.; Naranjo, J.; Vigara, L.A.; Cazorla, J.M.; Montero, M.E.; García, T.; Torrado, J.; Mazuecos, A. Impact of Physical Exercise in Patients with Chronic Kidney Disease: Sistematic Review and Meta-Analysis. Nefrologia 2020, 40, 237–252. [Google Scholar] [CrossRef]
- Ali, B.H.; Karaca, T.; al Suleimani, Y.; al Za’abi, M.; al Kalbani, J.; Ashique, M.; Nemmar, A. The Effect of Swimming Exercise on Adenine-Induced Kidney Disease in Rats, and the Inffuence of Curcumin or Lisinopril Thereon. PLoS ONE 2017, 12, 176316. [Google Scholar] [CrossRef]
- Odermatt, A. The Western-Style Diet: A Major Risk Factor for Impaired Kidney Function and Chronic Kidney Disease. Am. J. Physiol. Ren. Physiol. 2011, 301, F919–F931. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.S.; Righi, S.; Krieg, R.; Kang, L.; Carl, D.; Wang, J.; Massey, H.D.; Sica, D.A.; Gehr, T.W.B.; Ghosh, S. High Fat High Cholesterol Diet (Western Diet) Aggravates Atherosclerosis, Hyperglycemia and Renal Failure in Nephrectomized LDL Receptor Knockout Mice: Role of Intestine Derived Lipopolysaccharide. PLoS ONE 2015, 10, 141109. [Google Scholar] [CrossRef]
- Wang, X.T.; Sun, X.J.; Li, C.; Liu, Y.; Zhang, L.; Li, Y.D.; Wu, Q.H.; Li, S.Y.; Li, Y. Establishing a Cell-Based High-Content Screening Assay for TCM Compounds with Anti-Renal Fibrosis Effects. Evid.-Based Complement. Altern. Med. 2018, 2018. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Salloum, F.N.; Abbate, A.; Krieg, R.; Sica, D.A.; Gehr, T.W.; Kukreja, R.C. Curcumin Prevents Cardiac Remodeling Secondary to Chronic Renal Failure through Deactivation of Hypertrophic Signaling in Rats. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, 975–984. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Lang, X.; Cheng, D.; Yang, Z. Curcumin Ameliorates Chronic Renal Failure in 5/6 Nephrectomized Rats by Regulation of the MTOR/HIF-1α/VEGF Signaling Pathway. Biol. Pharm. Bull. 2019, 42, 886–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo, J.; Chirino, Y.I.; Molina-Jijón, E.; Andérica-Romero, A.C.; Tapia, E.; Pedraza-Chaverrí, J. Renoprotective Effect of the Antioxidant Curcumin: Recent Findings. Redox. Biol. 2013, 1, 448–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Feng, L.; Zhu, M.; Wang, R.-S.; Zhang, M.; Hu, S.; Jia, X.; Wu, J.-J. The In Vitro Protective Effects of Curcumin and Demethoxycurcumin in Curcuma longa Extract on Advanced Glycation End Products-Induced Mesangial Cell Apoptosis and Oxidative Stress. Planta Med. 2012, 78, 1757–1760. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Yang, L.; Ling, L.; Liu, Y.; Yu, Y.; Wu, Q.; Gu, Y.; Niu, J. Curcumin Attenuates Angiotensin II-Induced Podocyte Injury and Apoptosis by Inhibiting Endoplasmic Reticulum Stress. FEBS Open Bio 2020, 10, 1957–1966. [Google Scholar] [CrossRef]
- Afrin, M.R.; Arumugam, S.; Rahman, M.A.; Karuppagounder, V.; Harima, M.; Suzuki, H.; Miyashita, S.; Suzuki, K.; Ueno, K.; Yoneyama, H.; et al. Curcumin Reduces the Risk of Chronic Kidney Damage in Mice with Nonalcoholic Steatohepatitis by Modulating Endoplasmic Reticulum Stress and MAPK Signaling. Int. Immunopharmacol. 2017, 49, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.-Z.; Liu, J.; Zhang, Y.-Y.; Wang, H.-L.; Li, J.-C.; Liu, Y.-H.; Zhong, X.; Zhang, Y.-W.; Yan, Y.; Lan, H.-Y.; et al. Curcumin Relieved Cisplatin-Induced Kidney Inflammation through Inhibiting Mincle-Maintained M1 Macrophage Phenotype. Phytomedicine 2019, 52, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D. Introduction to NF-ΚB: Players, Pathways, Perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [Green Version]
- Song, N.; Thaiss, F.; Guo, L. NFκB and Kidney Injury. Front. Immunol. 2019, 10, 815. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Marakkath, B.; Ye, Y.; Khobahy, E.; Yan, M.; Hutcheson, J.; Zhu, J.; Zhou, X.; Mohan, C. Curcumin Attenuates Both Acute and Chronic Immune Nephritis. Int. J. Mol. Sci. 2020, 21, 1745. [Google Scholar] [CrossRef] [Green Version]
- Shing, C.M.; Adams, M.J.; Fassett, R.G.; Coombes, J.S. Nutritional Compounds Influence Tissue Factor Expression and Inflammation of Chronic Kidney Disease Patients in Vitro. Nutrition 2011, 27, 967–972. [Google Scholar] [CrossRef]
- Pivari, F.; Mingione, A.; Piazzini, G.; Ceccarani, C.; Ottaviano, E.; Brasacchio, C.; Dei Cas, M.; Vischi, M.; Cozzolino, M.G.; Fogagnolo, P.; et al. Curcumin Supplementation (Meriva®) Modulates Inflammation, Lipid Peroxidation and Gut Microbiota Composition in Chronic Kidney Disease. Nutrients 2022, 14, 231. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.; Salarolli, R.; Cardozo, L.F.M.F.; Santos, R.S.; de Brito, J.S.; Kemp, J.A.; Reis, D.; de Paiva, B.R.; Stenvinkel, P.; Lindholm, B.; et al. Impact of Curcumin Supplementation on Expression of Inflammatory Transcription Factors in Hemodialysis Patients: A Pilot Randomized, Double-Blind, Controlled Study. Clin. Nutr. 2020, 39, 3594–3600. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.; Cardozo, L.F.M.F.; da Cruz, B.O.; Paiva, B.R.; Fouque, D.; Mafra, D. Curcumin Supplementation Improves Oxidative Stress and Inflammation Biomarkers in Patients Undergoing Hemodialysis: A Secondary Analysis of a Randomized Controlled Trial. Int. Urol. Nephrol. 2022, 54, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Jaturapisanukul, S.; Kurathong, S.; Ngamvichchukorn, T.; Trakarnvanich, T. Curcuminoids Can Prevent Post-Contrast Acute Kidney Injury in Chronic Kidney Disease Patients: A Randomized Controlled Trial. Medicine 2022, 101, e30753. [Google Scholar] [CrossRef]
- Moreillon, J.J.; Bowden, R.G.; Deike, E.; Griggs, J.; Wilson, R.; Shelmadine, B.; Cooke, M.; Beaujean, A. The Use of an Anti-Inflammatory Supplement in Patients with Chronic Kidney Disease. J. Complement. Integr. Med. 2013, 10. [Google Scholar] [CrossRef]
- Shelmadine, B.D.; Bowden, R.G.; Moreillon, J.J.; Cooke, M.B.; Yang, P.; Deike, E.; Griggs, J.O.; Wilson, R.L. A Pilot Study to Examine the Effects of an Anti-Inflammatory Supplement on Eicosanoid Derivatives in Patients with Chronic Kidney Disease. J. Altern. Complement. Med. 2017, 23, 632–638. [Google Scholar] [CrossRef]
- Zubovic, S.V.; Kristic, S.; Prevljak, S.; Pasic, I.S. Chronic Kidney Disease and Lipid Disorders. Med. Arch. 2016, 70, 191–192. [Google Scholar] [CrossRef] [Green Version]
- Bugyei-Twum, A.; Abadeh, A.; Thai, K.; Zhang, Y.; Mitchell, M.; Kabir, G.; Connelly, K.A. Suppression of NLRP3 Inflammasome Activation Ameliorates Chronic Kidney Disease-Induced Cardiac Fibrosis and Diastolic Dysfunction. Sci. Rep. 2016, 6, 39551. [Google Scholar] [CrossRef]
- Correa, F.; Buelna-Chontal, M.; Hernández-Reséndiz, S.; García-Niño, W.R.; Roldán, F.J.; Soto, V.; Silva-Palacios, A.; Amador, A.; Pedraza-Chaverrí, J.; Tapia, E.; et al. Curcumin Maintains Cardiac and Mitochondrial Function in Chronic Kidney Disease. Free Radic. Biol. Med. 2013, 61, 119–129. [Google Scholar] [CrossRef]
- Hernández-Reséndiz, S.; Correa, F.; García-Niño, W.R.; Buelna-Chontal, M.; Roldán, F.J.; Ramírez-Camacho, I.; Delgado-Toral, C.; Carbó, R.; Pedraza-Chaverrí, J.; Tapia, E.; et al. Cardioprotection by Curcumin Post-Treatment in Rats with Established Chronic Kidney Disease. Cardiovasc. Drugs 2015, 29, 111–120. [Google Scholar] [CrossRef]
- Panahi, Y.; Ahmadi, Y.; Teymouri, M.; Johnston, T.P.; Sahebkar, A. Curcumin as a Potential Candidate for Treating Hyperlipidemia: A Review of Cellular and Metabolic Mechanisms. J. Cell. Physiol. 2018, 233, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Labban, R.S.M.; Alfawaz, H.A.; Almnaizel, A.T.; Al-Muammar, M.N.; Bhat, R.S.; El-Ansary, A. Garcinia Mangostana Extract and Curcumin Ameliorate Oxidative Stress, Dyslipidemia, and Hyperglycemia in High Fat Diet-Induced Obese Wistar Albino Rats. Sci. Rep. 2021, 11, 7278. [Google Scholar] [CrossRef] [PubMed]
- Sangartit, W.; Ha, K.B.; Lee, E.S.; Kim, H.M.; Kukongviriyapan, U.; Lee, E.Y.; Chung, C.H. Tetrahydrocurcumin Ameliorates Kidney Injury and High Systolic Blood Pressure in High-Fat Diet-Induced Type 2 Diabetic Mice. Endocrinol. Metab. 2021, 36, 810–822. [Google Scholar] [CrossRef] [PubMed]
- Altobelli, E.; Angeletti, P.M.; Marziliano, C.; Mastrodomenico, M.; Giuliani, A.R.; Petrocelli, R. Potential Therapeutic Effects of Curcumin on Glycemic and Lipid Profile in Uncomplicated Type 2 Diabetes—A Meta-Analysis of Randomized Controlled Trial. Nutrients 2021, 13, 404. [Google Scholar] [CrossRef] [PubMed]
- Sangouni, A.A.; Taghdir, M.; Mirahmadi, J.; Sepandi, M.; Parastouei, K. Effects of Curcumin and/or Coenzyme Q10 Supplementation on Metabolic Control in Subjects with Metabolic Syndrome: A Randomized Clinical Trial. Nutr. J. 2022, 21, 62. [Google Scholar] [CrossRef]
- Pandit, A.P.; Joshi, S.R.; Dalal, P.S.; Patole, V.C. Curcumin as a Permeability Enhancer Enhanced the Antihyperlipidemic Activity of Dietary Green Tea Extract. BMC Complement. Altern. Med. 2019, 19, 129. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cheng, F.; Luo, Y.; Zhan, Z.; Hu, P.; Ren, H.; Tang, H.; Peng, M. PEGylated Curcumin Derivative Attenuates Hepatic Steatosis via CREB/PPAR-γ/CD36 Pathway. BioMed Res. Int. 2017, 2017, 8234507. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Singh, V.K.; Kumar, D.; Yadav, P.; Kumar, S.; Beg, M.; Shankar, K.; Varshney, S.; Rajan, S.; Srivastava, A.; et al. Curcumin-3,4-Dichloro Phenyl Pyrazole (CDPP) Overcomes Curcumin’s Low Bioavailability, Inhibits Adipogenesis and Ameliorates Dyslipidemia by Activating Reverse Cholesterol Transport. Metabolism 2017, 73, 109–124. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Kim, M.-S. The Protective Effects of Curcumin on Metabolic Syndrome and Its Components: In-Silico Analysis for Genes, Transcription Factors, and MicroRNAs Involved. Arch. Biochem. Biophys. 2022, 727, 109326. [Google Scholar] [CrossRef]
- Ding, X.; Chen, Y.; Zhou, L.; Wu, R.; Jian, T.; Lyu, H.; Liu, Y.; Chen, J. Bisdemethoxycurcumin Attenuated Renal Injury via Activation of Keap1/Nrf2 Pathway in High-Fat Diet-Fed Mice. Int. J. Mol. Sci. 2022, 23, 7395. [Google Scholar] [CrossRef]
- Soetikno, V.; Sari, F.R.; Lakshmanan, A.P.; Arumugam, S.; Harima, M.; Suzuki, K.; Kawachi, H.; Watanabe, K. Curcumin Alleviates Oxidative Stress, Inflammation, and Renal Fibrosis in Remnant Kidney through the Nrf2-Keap1 Pathway. Mol. Nutr Food Res. 2013, 57, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- Ghelani, H.; Razmovski-Naumovski, V.; Chang, D.; Nammi, S. Chronic Treatment of Curcumin Improves Hepatic Lipid Metabolism and Alleviates the Renal Damage in Adenine-Induced Chronic Kidney Disease in Sprague-Dawley Rats. BMC Nephrol. 2019, 20, 431. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Woollard, K. Atherosclerosis. Adv. Exp. Med. Biol. 2017, 1003, 121–144. [Google Scholar] [PubMed]
- Futuhi, F.; Naghibzadeh Tahami, A.; Azmandian, J.; Saber, A. The Effects of Curcumin-Containing Supplementations on Inflammatory Markers and Lipid Profiles in Patients with Chronic Kidney Diseases: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Complement. Integr. Med. 2022, 19, 531–541. [Google Scholar] [CrossRef]
- Jiménez-Osorio, A.S.; García-Niño, W.R.; González-Reyes, S.; Álvarez-Mejía, A.E.; Guerra-León, S.; Salazar-Segovia, J.; Falcón, I.; Montes de Oca-Solano, H.; Madero, M.; Pedraza-Chaverri, J. The Effect of Dietary Supplementation With Curcumin on Redox Status and Nrf2 Activation in Patients With Nondiabetic or Diabetic Proteinuric Chronic Kidney Disease: A Pilot Study. J. Ren. Nutr. 2016, 26, 237–244. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceja-Galicia, Z.A.; Aranda-Rivera, A.K.; Amador-Martínez, I.; Aparicio-Trejo, O.E.; Tapia, E.; Trujillo, J.; Ramírez, V.; Pedraza-Chaverri, J. The Development of Dyslipidemia in Chronic Kidney Disease and Associated Cardiovascular Damage, and the Protective Effects of Curcuminoids. Foods 2023, 12, 921. https://doi.org/10.3390/foods12050921
Ceja-Galicia ZA, Aranda-Rivera AK, Amador-Martínez I, Aparicio-Trejo OE, Tapia E, Trujillo J, Ramírez V, Pedraza-Chaverri J. The Development of Dyslipidemia in Chronic Kidney Disease and Associated Cardiovascular Damage, and the Protective Effects of Curcuminoids. Foods. 2023; 12(5):921. https://doi.org/10.3390/foods12050921
Chicago/Turabian StyleCeja-Galicia, Zeltzin Alejandra, Ana Karina Aranda-Rivera, Isabel Amador-Martínez, Omar Emiliano Aparicio-Trejo, Edilia Tapia, Joyce Trujillo, Victoria Ramírez, and José Pedraza-Chaverri. 2023. "The Development of Dyslipidemia in Chronic Kidney Disease and Associated Cardiovascular Damage, and the Protective Effects of Curcuminoids" Foods 12, no. 5: 921. https://doi.org/10.3390/foods12050921
APA StyleCeja-Galicia, Z. A., Aranda-Rivera, A. K., Amador-Martínez, I., Aparicio-Trejo, O. E., Tapia, E., Trujillo, J., Ramírez, V., & Pedraza-Chaverri, J. (2023). The Development of Dyslipidemia in Chronic Kidney Disease and Associated Cardiovascular Damage, and the Protective Effects of Curcuminoids. Foods, 12(5), 921. https://doi.org/10.3390/foods12050921