

Purification and Characterization of the Enzyme Fucoidanase from Cobetia amphilecti Utilizing Fucoidan from Undaria pinnatifida

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Fucoidans
2.2. Screening and Identification of Strains of Marine Organisms
2.3. Mutagenesis of Strains of Marine Organisms
2.4. Enzyme Production and Growth Monitoring
2.5. Enzyme Extraction
2.6. Preliminary Purification of Fucoidanase
2.7. Assay of Enzymatic Activity
2.8. Determination of Enzymatic Characteristics
2.9. Preliminary Determination of Mode of Fucoidanase Degradation
2.10. Purification of Fucoidanase
2.11. Preparation of Enzymatic Solution of Fucoidan
3. Results and Discussion
3.1. Sequence Analysis of 16s rDNA
3.2. Production and Selection of Fucoidanase
3.3. Optimum Conditions and Factors Affecting Enzymatic Activity during Ultrasonication
3.4. Purification of Fucoidanase via Acetone Precipitation
3.5. Characteristics of Fucoidanase
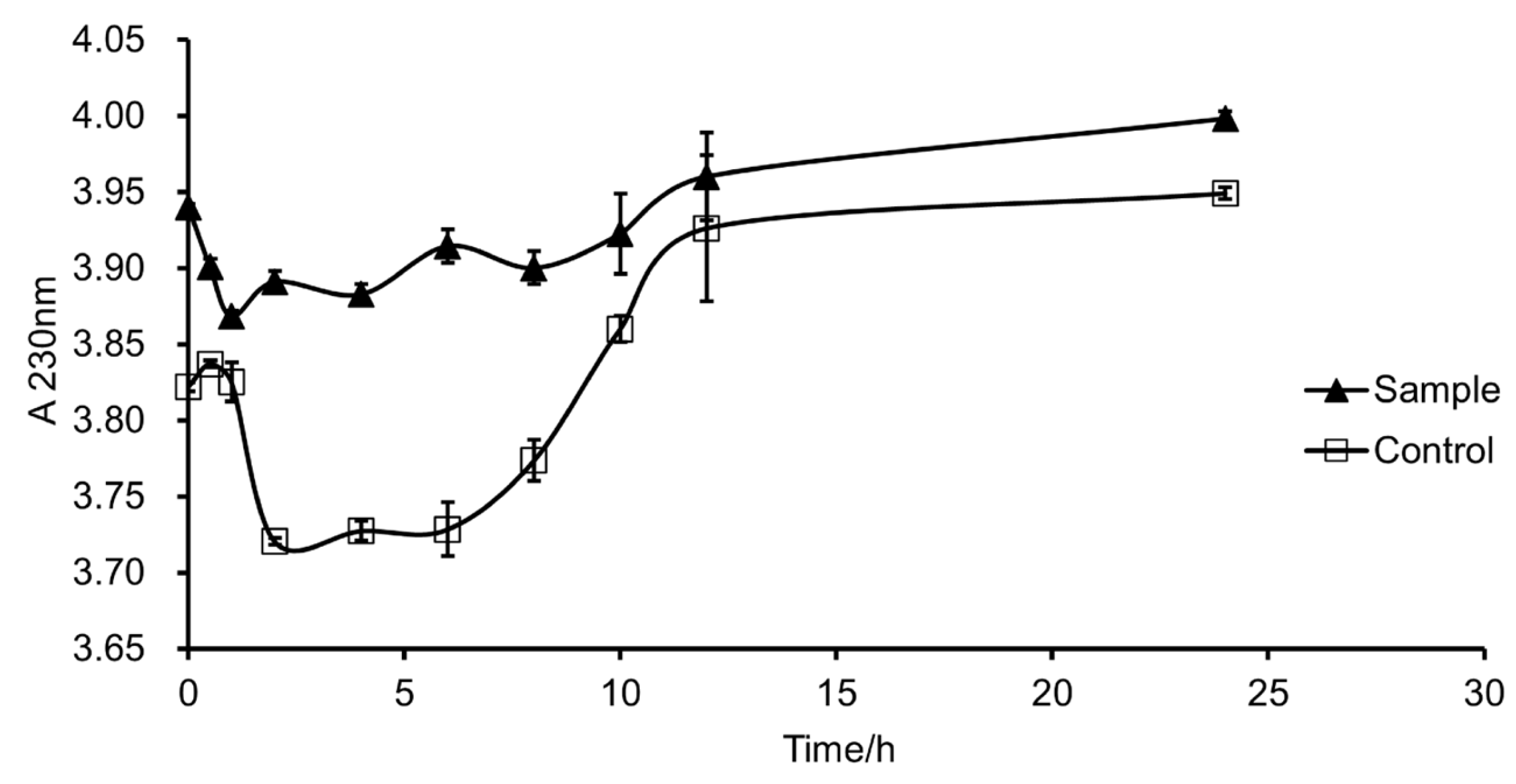
3.6. Degradation Mode of Fucoidanase
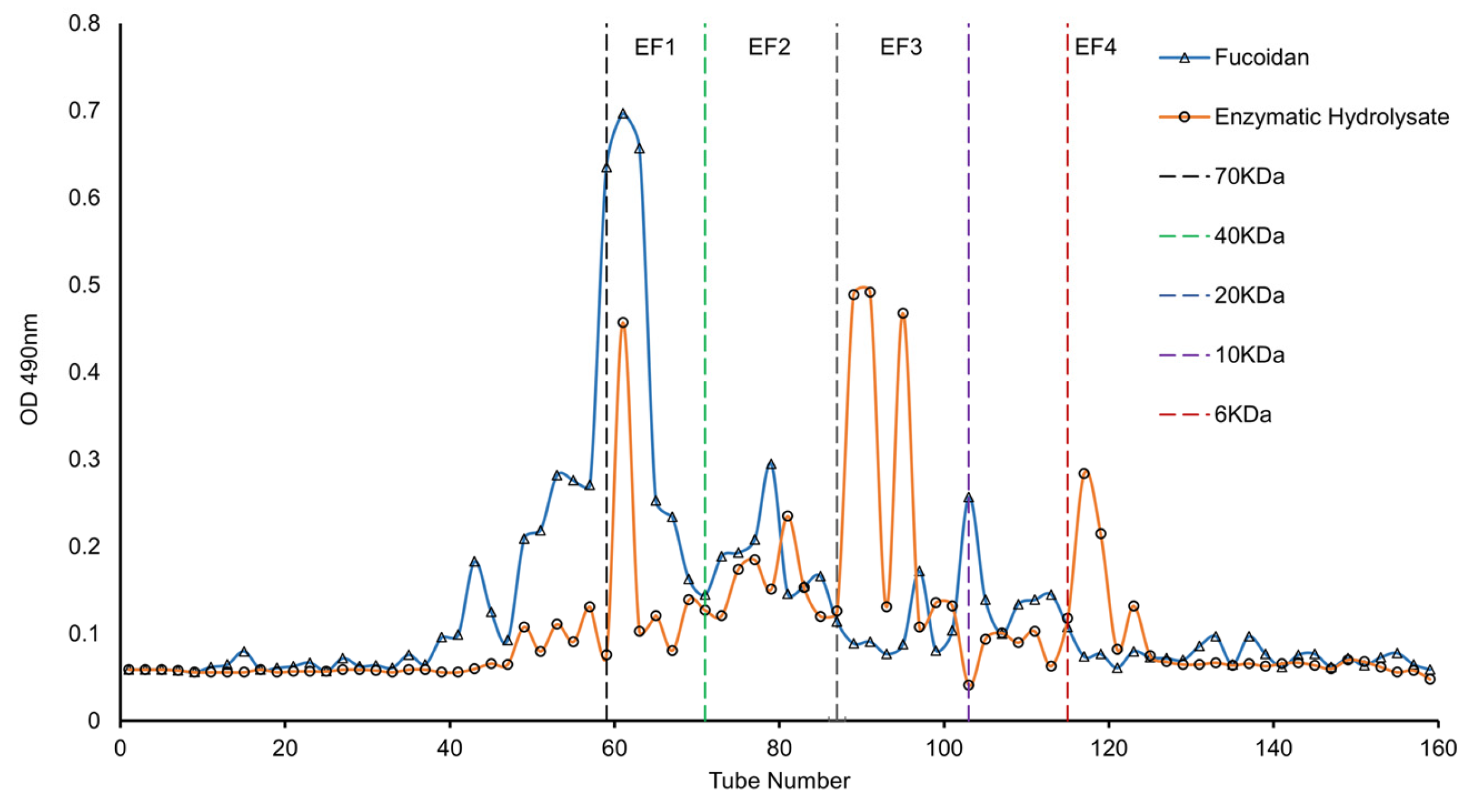
3.7. Molecular Weights of Degradation Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peng, Y.; Song, Y.; Wang, Q.; Hu, Y.; He, Y.; Ren, D.; Wu, L.; Liu, S.; Cong, H.; Zhou, H. In vitro and in vivo immunomodulatory effects of fucoidan compound agents. Int. J. Biol. Macromol. 2019, 127, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.J.; Lin, M.H.; Kuo, T.C.; Chou, C.M.; Mi, F.L.; Cheng, C.H.; Lin, C.W. Fucoidan from Laminaria japonica exerts antitumor effects on angiogenesis and micrometastasis in triple-negative breast cancer cells. Int. J. Biol. Macromol. 2020, 149, 600–608. [Google Scholar] [CrossRef]
- Sun, Q.L.; Li, Y.; Ni, L.Q.; Li, Y.X.; Cui, Y.S.; Jiang, S.L.; Xie, E.Y.; Du, J.; Deng, F.; Dong, C.X. Structural characterization and antiviral activity of two fucoidans from the brown algae Sargassum henslowianum. Carbohydr. Polym. 2020, 229, 115487. [Google Scholar] [CrossRef]
- Albuquerque, I.R.L.; Queiroz, K.C.S.; Alves, L.G.; Santos, E.A.; Leite, E.L.; Rocha, H.A.O. Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz. J. Med. Biol. Res. 2004, 37, 167–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, D.J.; Krylov, V.S. Anti-HIV activity of extracts and compounds from algae and cyanobacteria. Ecotoxicol. Environ. Saf. 2000, 45, 208–227. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, Q.; Song, Y.; He, Y.; Ren, D.; Cong, H.; Wu, L. Studies on the hepatoprotective effect of fucoidans from brown algae Kjellmaniella crassifolia. Carbohydr. Polym. 2018, 193, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.J.; Wan Aida, W.M.; Schiehser, S.; Rosenau, T.; Bohmdorfer, S. Structural elucidation of fucoidan from Cladosiphon okamuranus (Okinawa mozuku). Food Chem. 2019, 272, 222–226. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Fucoidan-degrading fungal strains: Screening, morphometric evaluation, and influence of medium composition. Appl. Biochem. Biotechnol. 2010, 162, 2177–2188. [Google Scholar] [CrossRef]
- Morya, V.K.; Kim, J.; Kim, E.K. Algal fucoidan: Structural and size-dependent bioactivities and their perspectives. Appl. Microbiol. Biotechnol. 2012, 93, 71–82. [Google Scholar] [CrossRef]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A quantitative method to detect fucoidan in human plasma using a novel antibody. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 705–710. [Google Scholar] [CrossRef]
- Bachelet, L.; Bertholon, I.; Lavigne, D.; Vassy, R.; Jandrot-Perrus, M.; Chaubet, F.; Letourneur, D. Affinity of low molecular weight fucoidan for P-selectin triggers its binding to activated human platelets. Biochim. Biophys. Acta 2009, 1790, 141–146. [Google Scholar] [CrossRef]
- Deux, J.F. Low molecular weight fucoidan prevents neointimal hyperplasia inrabbit iliac artery in-stent restenosis model. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1604–1609. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Zheng, Y.; Zhang, Q.; Wang, J.; Wang, L.; Yang, W.; Guo, C.; Gao, W.; Wang, X.; Luo, D. Low-molecular-weight fucoidan protects endothelial function and ameliorates basal hypertension in diabetic Goto-Kakizaki rats. Lab. Investig. 2014, 94, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Heo, S.J.; Park, E.J.; Lee, K.W.; Jeon, Y.J. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol. 2005, 96, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Je, J.-Y.; Park, P.-J.; Kim, E.-K.; Park, J.-S.; Yoon, H.-D.; Kim, K.-R.; Ahn, C.-B. Antioxidant activity of enzymatic extracts from the brown seaweed Undaria pinnatifida by electron spin resonance spectroscopy. LWT—Food Sci. Technol. 2009, 42, 874–878. [Google Scholar] [CrossRef]
- Qiu, Y.; Jiang, H.; Dong, Y.; Wang, Y.; Hamouda, H.I.; Balah, M.A.; Mao, X. Expression and Biochemical Characterization of a Novel Fucoidanase from Flavobacteriumalgicola with the Principal Product of Fucoidan-Derived Disaccharide. Foods 2022, 11, 1025. [Google Scholar] [CrossRef]
- Liang, Z.; Zheng, Y.; Wang, J.; Zhang, Q.; Ren, S.; Liu, T.; Wang, Z.; Luo, D. Low molecular weight fucoidan ameliorates streptozotocin-induced hyper-responsiveness of aortic smooth muscles in type 1 diabetes rats. J. Ethnopharmacol. 2016, 191, 341–349. [Google Scholar] [CrossRef]
- Liu, T.; Wang, Z.; Chen, X.; You, H.; Xue, J.; Cai, D.; Zheny, Y.; Xu, Y.; Luo, D. Low molecular weight fucoidan protects against hindlimb ischemic injury in type 2 diabetic mice through enhancing endothelial nitric oxide synthase phosphorylation. J. Diabetes 2018, 10, 820–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.J.; Lee, B.Y. Fucoidan from the sporophyll of Undaria pinnatifida suppresses adipocyte differentiation by inhibition of inflammation-related cytokines in 3T3-L1 cells. Nutr. Res. 2012, 32, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Bleha, R.; Synytsya, A.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu fucoidan: Structural complexity and defensive effects against avian influenza A viruses. Carbohydr. Polym. 2014, 111, 633–644. [Google Scholar] [CrossRef]
- Lu, M.-K.; Cheng, J.-J.; Lin, C.-Y.; Chang, C.-C. Purification, structural elucidation, and anti-inflammatory effect of a water-soluble 1,6-branched 1,3-α-d-galactan from cultured mycelia of Poria cocos. Food Chem. 2010, 118, 349–356. [Google Scholar] [CrossRef]
- Chevolot, L.; Foucault, A.; Chaubet, F.; Kervarec, N.; Sinquin, C.; Fisher, A.; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr. Res. 1999, 319, 154–165. [Google Scholar] [CrossRef]
- Wen, Y.; Gao, L.; Zhou, H.; Ai, C.; Huang, X.; Wang, M.; Zhang, Y.; Zhao, C. Opportunities and challenges of algal fucoidan for diabetes management. Trends Food Sci. Technol. 2021, 111, 628–641. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, Z.; Ren, L.; Jiao, S.; Zhang, X.; Wang, Q.; Li, Z.; Du, Y.; Li, J.J. Overexpression and biochemical characterization of a truncated endo-alpha (1→3)-fucoidanase from Alteromonas sp. SN-1009. Food Chem. 2021, 353, 129460. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, M.; Wu, K.; Liu, B.; Cai, J.; Pan, R. Purification and characteristics of fucoidanase obtained from Dendryphiella arenaria TM94. J. Appl. Phycol. 2010, 23, 197–203. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Shevchenko, L.S.; Nedashkovskaya, O.I.; Shevchenko, N.M.; Alekseeva, S.A.; Mikhailov, V.V.; Zvyagintseva, T.N. Screening of marine bacteria for fucoidanases. Mikrobiologiya 2000, 69, 370–376. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Nedashkovskaya, O.I.; Alekseeva, S.A.; Ivanova, E.P.; Romanenko, L.A.; Gorshkova, N.M.; Isakov, V.V.; Zvyagintseva, T.N.; Mikhailov, V.V. Degradation of Fucoidan by the Marine Proteobacterium Pseudoalteromonas citrea. Microbiology 2002, 71, 41–47. [Google Scholar] [CrossRef]
- Sakai, T.; Ishizuka, K.; Kato, I. Isolation and characterization of a fucoidan-degrading marine bacterium. Mar. Biotechnol. 2003, 5, 409–416. [Google Scholar] [CrossRef]
- Chang, Y.; Xue, C.; Tang, Q.; Li, D.; Wu, X.; Wang, J. Isolation and characterization of a sea cucumber fucoidan-utilizing marine bacterium. Lett. Appl. Microbiol. 2010, 50, 301–307. [Google Scholar] [CrossRef]
- Kim, W.J.; Kim, S.M.; Lee, Y.H.; Kim, H.G.; Park, Y.I. Isolation and characterization of marine bacterial strain degrading fucoidan from korean undaria pinnatifida sporophylls. J. Microbiol. Biotechnol. 2008, 18, 616–623. [Google Scholar]
- Silchenko, A.S.; Kusaykin, M.I.; Zakharenko, A.M.; Menshova, R.V.; Khanh, H.H.N.; Dmitrenok, P.S.; Isakov, V.V.; Zvyagintseva, T.N. Endo-1,4-fucoidanase from Vietnamese marine mollusk Lambis sp. which producing sulphated fucooligosaccharides. J. Mol. Catal. B Enzym. 2014, 102, 154–160. [Google Scholar] [CrossRef]
- Kusaykin, M.I.; Silchenko, A.S.; Zakharenko, A.M.; Zvyagintseva, T.N. Fucoidanases. Glycobiology 2016, 26, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Ono, A.; Suzuki, T.; Gotoh, S.; Kono, H.; Matsui, M.; Aoki, D.; Matsuda, M.; Kawagishi, H.; Ogata, M. Structural investigation of alpha-l-fucosidase from the pancreas of Patiria pectinifera, based on molecular cloning. Carbohydr. Res. 2019, 475, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Sakai, T.; Kojima, K.; Nakayama, S.; Nakanishi, Y.; Kato, I. Partial purification and characterization of an enzyme releasing 2-sulfo-alpha-l-fucopyranose from 2-sulfo-alpha-l-fucopyranosyl-(1→2) pyridylaminated fucose from a sea urchin, Strongylocentrotus nudus. Biosci. Biotechnol. Biochem. 1996, 60, 666–668. [Google Scholar] [CrossRef]
- Thanassi, N.M.; Nakada, H.I. Enzymic degradation of fucoidan by enzymes from the hepatopancreas of abalone, Haliotus species. Arch. Biochem. Biophys. 1967, 118, 172–177. [Google Scholar] [CrossRef]
- Kitamura, K.; Matsuo, M.; Yasui, T. Enzymatic degradation of fucoidan by fucoidanase from the hepatopancreas of Patinopecten yessoensis. Biosci. Biotechnol. Biochem. 1992, 56, 490–494. [Google Scholar] [CrossRef]
- Sakai, T.; Kimura, H.; Kato, I. A marine strain of flavobacteriaceae utilizes brown seaweed fucoidan. Mar. Biotechnol. 2002, 4, 399–405. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujikawa, T.; Koga, D.; Ide, A. Purification and some properties of exotype fucoidanases from Vibrio sp. N-5. Biosci. Biotechnol. Biochem. 1992, 56, 1829–1834. [Google Scholar] [CrossRef]
- Nagao, T.; Arai, Y.; Yamaoka, M.; Komatsu, F.; Yagi, H.; Suzuki, H.; Ohshiro, T. Identification and characterization of the fucoidanase gene from Luteolibacter algae H18. J. Biosci. Bioeng. 2018, 126, 567–572. [Google Scholar] [CrossRef]
- Vuillemin, M.; Silchenko, A.S.; Cao, H.T.T.; Kokoulin, M.S.; Trang, V.T.D.; Holck, J.; Ermakova, S.P.; Meyer, A.S.; Mikkelsen, M.D. Functional Characterization of a New GH107 Endo-alpha-(1,4)-Fucoidanase from the Marine Bacterium Formosa haliotis. Mar. Drugs 2020, 18, 562. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Jasso, R.M.; Mussatto, S.I.; Sepúlveda, L.; Agrasar, A.T.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Fungal fucoidanase production by solid-state fermentation in a rotating drum bioreactor using algal biomass as substrate. Food Bioprod. Process. 2013, 91, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Hifney, A.F.; Fawzy, M.A.; Abdel-Gawad, K.M.; Gomaa, M. Upgrading the antioxidant properties of fucoidan and alginate from Cystoseira trinodis by fungal fermentation or enzymatic pretreatment of the seaweed biomass. Food Chem. 2018, 269, 387–395. [Google Scholar] [CrossRef]
- Gomaa, M.; Fawzy, M.A.; Hifney, A.F.; Abdel-Gawad, K.M. Optimization of enzymatic saccharification of fucoidan and alginate from brown seaweed using fucoidanase and alginate lyase from the marine fungus Dendryphiella arenaria. J. Appl. Phycol. 2018, 31, 1955–1965. [Google Scholar] [CrossRef]
- Wang, Q.; Song, Y.; He, Y.; Ren, D.; Kow, F.; Qiao, Z.; Liu, S.; Yu, X. Structural characterisation of algae Costaria costata fucoidan and its effects on CCl(4)-induced liver injury. Carbohydr. Polym. 2014, 107, 247–254. [Google Scholar] [CrossRef]
- Soedjak, H.S. Colorimetric determination of carrageenans and other anionic hydrocolloids with methylene blue. Anal. Chem. 1994, 66, 4514–4518. [Google Scholar] [CrossRef]
- Thippeswamy, S.; Girigowda, K.; Mulimani, V. Isolation and identification of α-amylase producing Bacillus sp. from dhal industry waste. Indian. J. Biochem. Biophys. 2006, 43, 295–298. [Google Scholar]
- Sugita, T.; Takashima, M.; Kodama, M.; Tsuboi, R.A.N. Description of a new yeast species, Malassezia japonica, and its detection in patients with atopic dermatitis and healthy subjects. J. Clin. Microbiol. 2003, 41, 4695–4699. [Google Scholar] [CrossRef] [Green Version]
- Ying, W. Screening of fucoindanase-producing bacteria fucoindanase characerization, and establishment of the hydrolyzate antioxidant activity predication system. J. Ethnopharmacol. 2013, 87, 241–246. [Google Scholar] [CrossRef]
- Chen, Q.; Kou, L.; Wang, F.; Wang, Y. Size-dependent whitening activity of enzyme-degraded fucoidan from Laminaria japonica. Carbohydr. Polym. 2019, 225, 115211. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, L.A.; Tanaka, N.; Svetashev, V.I.; Falsen, E. Description of Cobetia amphilecti sp. nov., Cobetia litoralis sp. nov. and Cobetia pacifica sp. nov., classification of Halomonas halodurans as a later heterotypic synonym of Cobetia marina and emended descriptions of the genus Cobetia and Cobetia marina. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 1, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Farahat, M.G.; Amr, D.; Galal, A. Molecular cloning, structural modeling and characterization of a novel glutaminase-free L-asparaginase from Cobetia amphilecti AMI6. Int. J. Biol. Macromol. 2020, 143, 685–695. [Google Scholar] [CrossRef]
- Balabanova, L.; Podvolotskaya, A.; Slepchenko, L.; Eliseikina, M.; Noskova, Y.; Nedashkovskaya, O.; Son, O.; Tekutyeva, L.; Rasskazov, V. Nucleolytic enzymes from the marine bacterium Cobetia amphilecti KMM 296 with antibiofilm activity and biopreservative effect on meat products. Food Control. 2017, 78, 270–278. [Google Scholar] [CrossRef]
- Gnaim, R.; Unis, R.; Gnayem, N.; Das, J.; Gozin, M.; Golberg, A. Turning mannitol-rich agricultural waste to poly(3-hydroxybutyrate) with Cobetia amphilecti fermentation and recovery with methyl levulinate as a green solvent. Bioresour. Technol. 2022, 352, 127075. [Google Scholar] [CrossRef]
- Wang, Y.; Niu, D.; Que, F.; Li, Y.; Chen, Q. Low molecular weight fucoidan prepared by fucoidanase degradation—A promising browning inhibitor. LWT 2021, 148, 111739. [Google Scholar] [CrossRef]
- Zhou, J.; Mao, Z.; Chen, M.S.F. Optimization of conditions for extraction of vanillic acid decarboxylase by ultrasonic crushing. Food Ind. 2019, 40, 155–159. [Google Scholar]
- Burtseva, Y.V.; Kusaikin, M.I.; Sova, V.V.; Shevchenko, N.M.; Skobun, A.S.; Zvyagintseva, T.N. Distribution of fucoidan hydrolases and some glycosidases among marine invertebrates. Russ. J. Mar. Biol. 2000, 26, 453–456. [Google Scholar] [CrossRef]
- Burtseva, Y.V.; Verigina, N.S.; Sova, V.V.; Pivkin, M.V.; Zvyagintseva, T.N. Filamentous marine fungi as producers of O-glycosylhydrolases: β-1,3-glucanase from Chaetomium indicum. Mar. Biotechnol. 2003, 5, 349–359. [Google Scholar] [CrossRef]
- Fujikawa, T.; Koyabu, K.; Wada, M. Enzymes in Hepatopancreas of Abalone Active on Fucoidan (1) Crude Enzyme and Unadsorbed Fraction on CM-Cellulose. Nippon. Nōgeikagaku Kaishi 1979, 53, 87–95. [Google Scholar] [CrossRef]
- Kim, W.; Park, J.; Park, J.; Choi, D.; Park, Y. Purification and Characterization of a Fucoidanase (FNase S) from a Marine Bacterium Sphingomonas paucimobilis PF-1. Mar. Drugs 2015, 13, 4398–4417. [Google Scholar] [CrossRef] [Green Version]
- Trang, V.T.D.; Mikkelsen, M.D.; Vuillemin, M.; Meier, S.; Cao, H.T.T.; Muschiol, J.; Perna, V.; Nguyen, T.T.; Tran, V.H.N.; Holck, J.; et al. The Endo-α(1,4) Specific Fucoidanase Fhf2 from Formosa haliotis Releases Highly Sulfated Fucoidan Oligosaccharides. Front Plant Sci. 2022, 13, 823668. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.; Ustyuzhanina, N.; Kusaykin, M.; Krylov, V.; Shashkov, A.; Dmitrenok, A.; Usoltseva, R.; Zueva, A.; Nifantiev, N.; Zvyagintseva, T. Expression and biochemical characterization and substrate specificity of the fucoidanase from Formosa algae. Glycobiology 2016, 27, 254–263. [Google Scholar]
- Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Kalinovsky, A.I.; Miansong, Z.; Changheng, L.; Malyarenko, O.; Zueva, A.O.; Zvyagintseva, T.N.; Ermakova, S.P. Structure, enzymatic transformation, anticancer activity of fucoidan and sulphated fucooligosaccharides from Sargassum horneri. Carbohydr. Polym. 2017, 175, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Kato, K. Isolation and Characterization of a Fucoidan-Degrading Marine Bacterial Strain and Its Fucoidanase. Mar. Biotechnol. 2004, 6, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Linhardt, R.J.; Cooney, C.L.; Galliher, P.M. Polysaccharide lyases. Appl. Biochem. Biotechnol. 1986, 12, 135–177. [Google Scholar] [CrossRef]
- Linhardt, R.J. Analysis of glycosaminoglycans with polysaccharide lyases. Curr. Protoc. Mol. Biol. 2001, 17, t13–t17. [Google Scholar]
- Cao, H.T.T.; Mikkelsen, M.D.; Lezyk, M.J.; Bui, L.M.; Tran, V.T.T.; Silchenko, A.S.; Kusaykin, M.I.; Pham, T.D.; Truong, B.H.; Holck, J.; et al. Novel Enzyme Actions for Sulphated Galactofucan Depolymerisation and a New Engineering Strategy for Molecular Stabilisation of Fucoidan Degrading Enzymes. Mar. Drugs 2018, 16, 422. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Enzymolysis Liquid: Acetone (V:V) | Total Activity (U) | Specific Activity (U mg−1) | Purification (Fold) | Yield (100%) |
|---|---|---|---|---|---|
| 1 | 1:0 | 6.60 | 0.41 | 1.00 | 100 |
| 2 | 1:1.0 | 2.49 | 0.16 | 0.39 | 38 |
| 3 | 1:1.5 | 3.48 | 0.25 | 0.60 | 53 |
| 4 | 1:2.0 | 6.44 | 0.43 | 1.21 | 98 |
| 5 | 1:2.5 | 4.44 | 0.38 | 0.93 | 67 |
| 6 | 1:3.0 | 5.56 | 0.38 | 0.92 | 84 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Wang, Q.; Shao, Z.; Liu, Q.; He, Y.; Ren, D.; Yang, H.; Li, X. Purification and Characterization of the Enzyme Fucoidanase from Cobetia amphilecti Utilizing Fucoidan from Undaria pinnatifida. Foods 2023, 12, 1555. https://doi.org/10.3390/foods12071555
Liu S, Wang Q, Shao Z, Liu Q, He Y, Ren D, Yang H, Li X. Purification and Characterization of the Enzyme Fucoidanase from Cobetia amphilecti Utilizing Fucoidan from Undaria pinnatifida. Foods. 2023; 12(7):1555. https://doi.org/10.3390/foods12071555
Chicago/Turabian StyleLiu, Shu, Qiukuan Wang, Zhenwen Shao, Qi Liu, Yunhai He, Dandan Ren, Hong Yang, and Xiang Li. 2023. "Purification and Characterization of the Enzyme Fucoidanase from Cobetia amphilecti Utilizing Fucoidan from Undaria pinnatifida" Foods 12, no. 7: 1555. https://doi.org/10.3390/foods12071555
APA StyleLiu, S., Wang, Q., Shao, Z., Liu, Q., He, Y., Ren, D., Yang, H., & Li, X. (2023). Purification and Characterization of the Enzyme Fucoidanase from Cobetia amphilecti Utilizing Fucoidan from Undaria pinnatifida. Foods, 12(7), 1555. https://doi.org/10.3390/foods12071555