
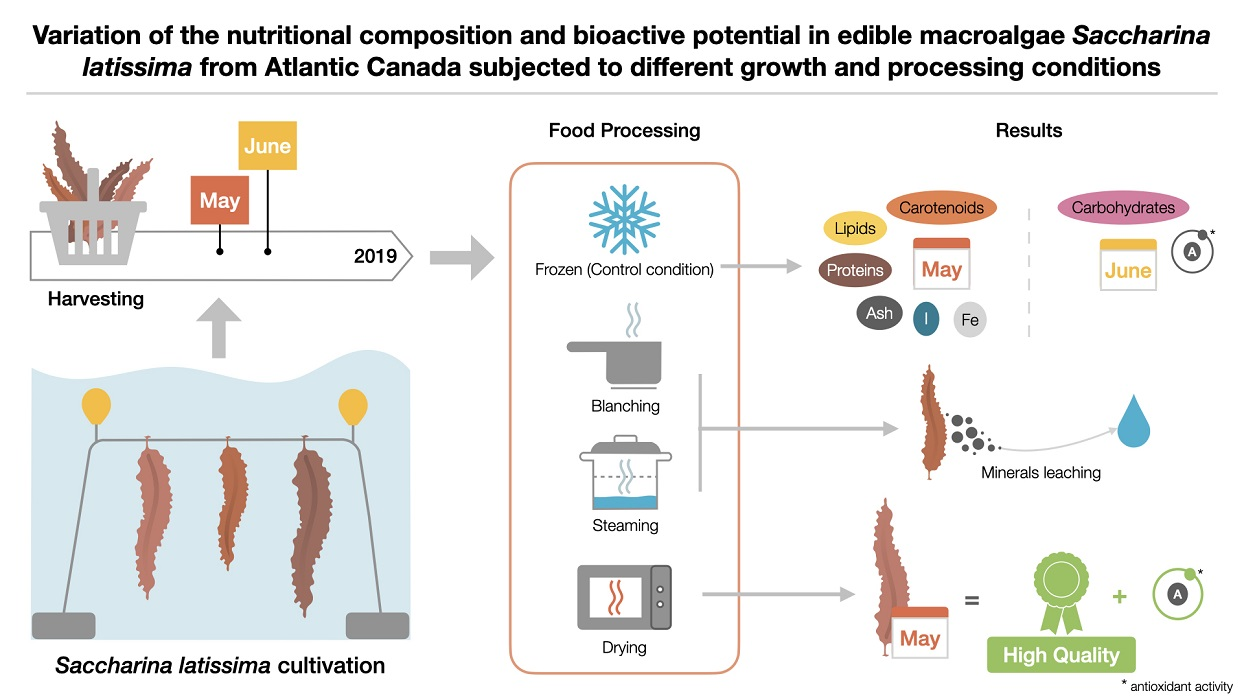
Variation of the Nutritional Composition and Bioactive Potential in Edible Macroalga Saccharina latissima Cultivated from Atlantic Canada Subjected to Different Growth and Processing Conditions

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
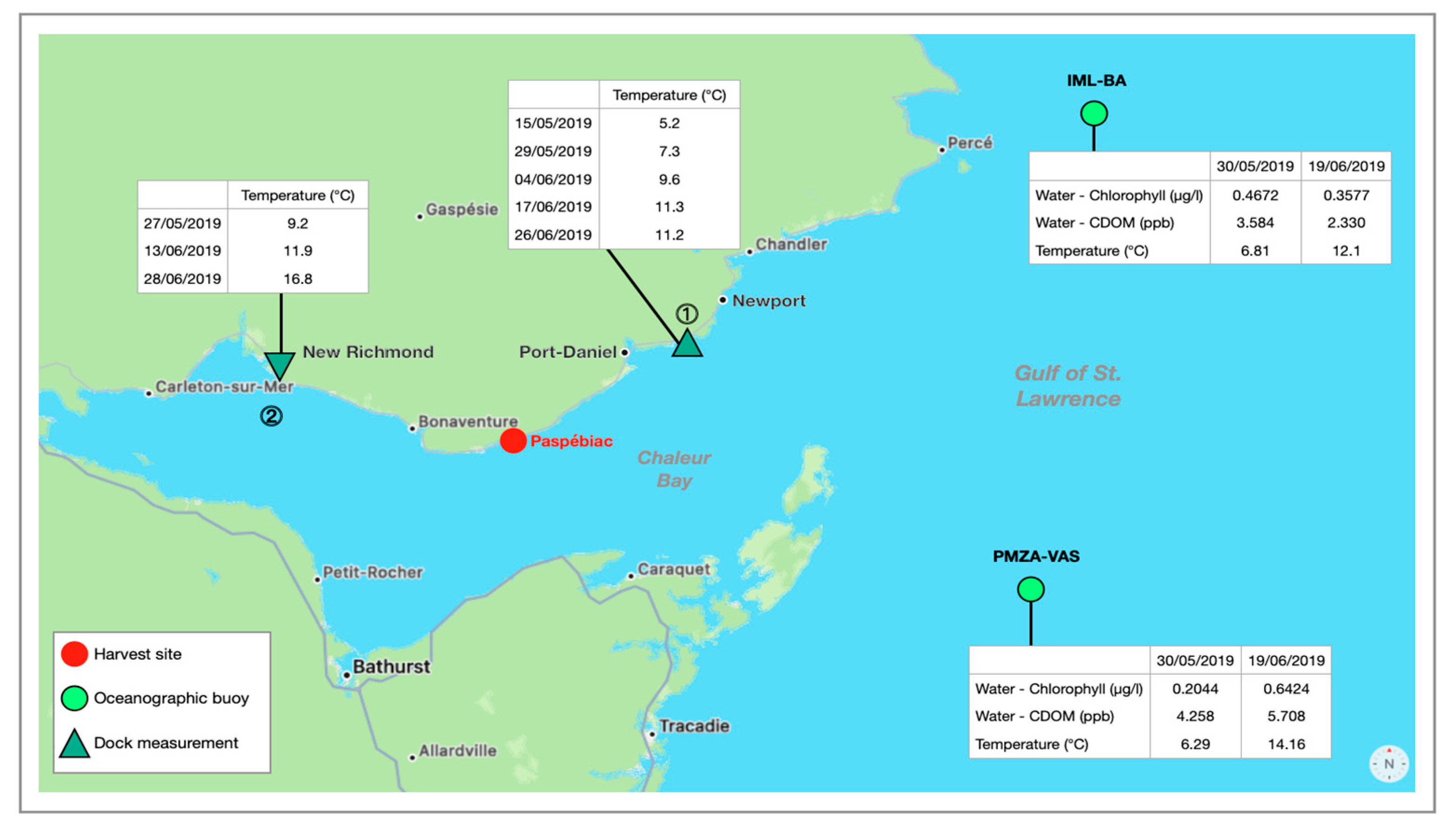
2.1. Macroalgal Biomass
2.2. Processing of the Blades
2.3. Nutritional Composition (Lipids, Ash, Proteins, Carbohydrates, Fibers) and Mineral Composition
2.4. Potential Bioactive Compound Determination
2.4.1. Total Phenolic Content
2.4.2. Carotenoids
2.4.3. Alginates, Fucoidans, and Laminarans
2.5. In Vitro Bioactive Potential
2.5.1. Water-Soluble S. latissima Extracts
2.5.2. Oxygen Radical Absorbance Capacity (ORAC) Assay
2.5.3. Ferric Ion Reducing Antioxidant Power (FRAP) Assay
2.6. Statistical Analysis
3. Results and Discussion
3.1. Chemical and Mineral Compositions
3.1.1. Variations through Growth Conditions of Crude S. latissima
3.1.2. Variation through Food Processing of S. latissima
- Blanching
- Steaming
- Drying
3.2. Bioactive Compound Contents and In Vitro Bioactive Potential
3.2.1. Variations through Growth Conditions of Crude S. latissima
- Bioactive compound content
- Protein content and extraction yields of soluble S. latissima extracts
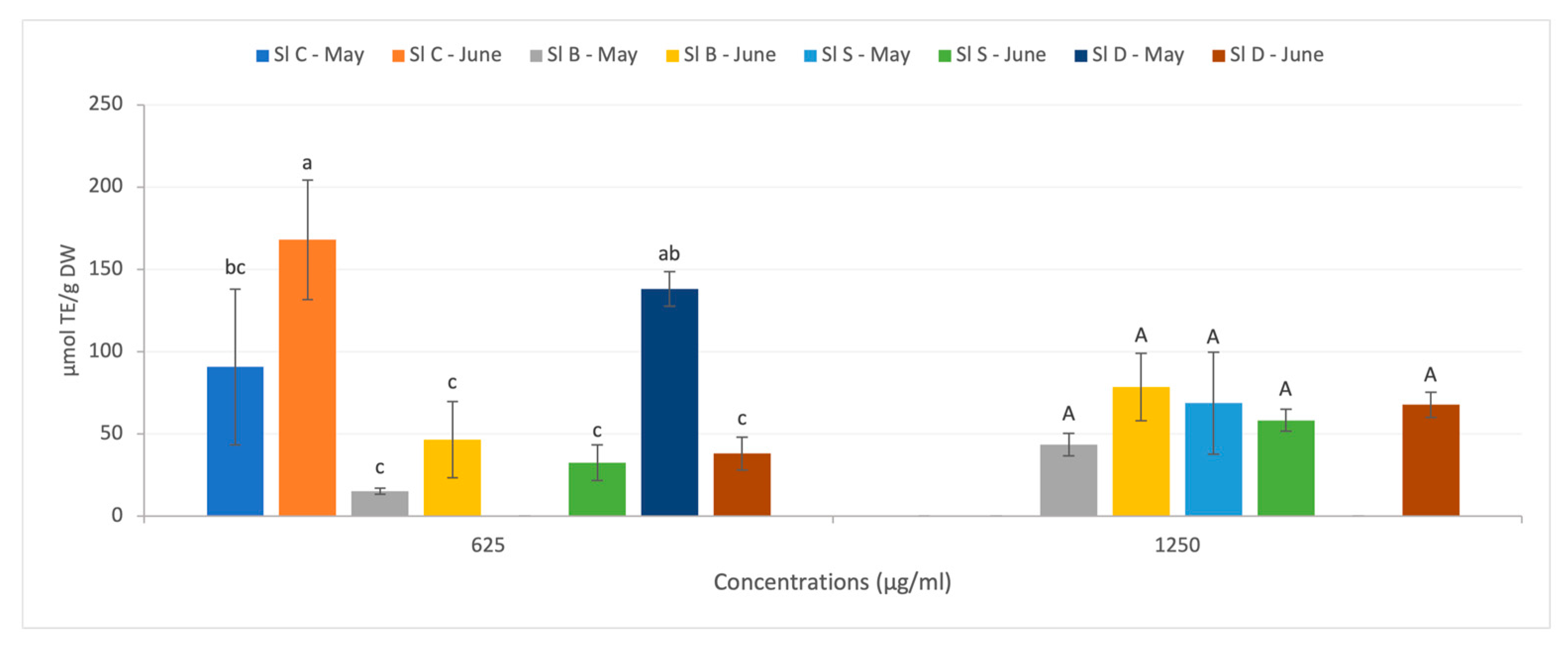
- Antioxidant capacity of soluble S. latissima extracts: ORAC and FRAP tests
3.2.2. Variation through Food Processing of S. latissima
- Blanching
- Steaming
- Drying
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsu, S.Y.; Wang, H.C.; Ho, J.L.; Chen, H.C. Exploring Consumers’ Interest in Choosing Sustainable Food. Front. Psychol. 2020, 11, 489. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Munir, I.U.; Hyder, S.; Nassani, A.A.; Qazi Abro, M.M.; Zaman, K. Sustainable food production, forest biodiversity and mineral pricing: Interconnected global issues. Resour. Policy 2020, 65, 101583. [Google Scholar] [CrossRef]
- Cai, J.; Lovatelli, A.; Aguilar-Manjarrez, J.; Cornish, L.; Dabbadie, L.; Desrochers, A.; Diffey, S.; Garrido Gamarro, E.; Geehan, J.; Hurtado, A.; et al. Seaweeds and microalgae: An overview for unlocking their potential in global aquaculture development. FAO Fish. Aquac. Circ. 2021, 1229, 48. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Rhatigan, P.; Pérez-Lloréns, J.-L. World cuisine of seaweeds: Science meets gastronomy. Int. J. Gastron. Food Sci. 2018, 14, 55–65. [Google Scholar] [CrossRef]
- Rioux, L.-E.; Beaulieu, L.; Turgeon, S.L. Seaweeds: A traditional ingredients for new gastronomic sensation. Food Hydrocoll. 2017, 68, 255–265. [Google Scholar] [CrossRef]
- Bourgougnon, N.; Gervois, A. Diversité des lignées et habitats. In Les Algues Marines: Biologie, Ecologie et Utilisation; Ellipses: Paris, France, 2021; pp. 12–30. [Google Scholar]
- Bleakley, S.; Hayes, M. Algal Proteins: Extraction, Application, and Challenges Concerning Production. Foods 2017, 6, 33. [Google Scholar] [CrossRef]
- Cho, G.; Yoon, H.; Boo, S.; Yarish, C. Atlantic kelp species Laminaria longicruris and L. saccharina (Laminariales) are conspecific. J. Phycol. 2000, 36, 12–13. [Google Scholar] [CrossRef]
- Croci, D.O.; Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; Piccoli, A.; Totani, L.; Ustyuzhanina, N.E.; Bilan, M.I.; Usov, A.I.; Grachev, A.A.; et al. Fucans, but not fucomannoglucuronans, determine the biological activities of sulfated polysaccharides from Laminaria saccharina brown seaweed. PLoS ONE 2011, 6, e17283. [Google Scholar] [CrossRef] [PubMed]
- Sappati, P.K.; Nayak, B.; VanWalsum, G.P.; Mulrey, O.T. Combined effects of seasonal variation and drying methods on the physicochemical properties and antioxidant activity of sugar kelp (Saccharina latissima). J. Appl. Phycol. 2018, 31, 1311–1332. [Google Scholar] [CrossRef]
- Côté-Laurin, M.-C.; Berger, K.; Tamigneaux, É. Manuel pour la Récolte Commerciale des Macroalgues du Québec; Merinov: Grande-Rivère, QC, Canada, 2016; 89p. [Google Scholar]
- Boisvert, C.; Beaulieu, L.; Bonnet, C.; Pelletier, É. Assessment of the Antioxidant and Antibacterial Activities of Three Species of Edible Seaweeds. J. Food Biochem. 2015, 39, 377–387. [Google Scholar] [CrossRef]
- Hell, A.; Labrie, S.; Beaulieu, L. Effect of seaweed flakes addition on the development of bioactivities in functional Camembert-type cheese. Int. J. Food Sci. Technol. 2018, 53, 1054–1064. [Google Scholar] [CrossRef]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [PubMed]
- Roleda, M.Y.; Marfaing, H.; Desnica, N.; Jónsdóttir, R.; Skjermo, J.; Rebours, C.; Nitschke, U. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: Health risk assessment and implication for food applications. Food Control. 2019, 95, 121–134. [Google Scholar] [CrossRef]
- Tanna, B.; Mishra, A. Nutraceutical Potential of Seaweed Polysaccharides: Structure, Bioactivity, Safety, and Toxicity. Compr. Rev. Food Sci. Food Saf. 2019, 18, 817–831. [Google Scholar] [CrossRef]
- Bartsch, I.; Wiencke, C.; Bischof, K.; Buchholz, C.M.; Buck, B.H.; Eggert, A.; Feuerpfeil, P.; Hanelt, D.; Jacobsen, S.; Karez, R.; et al. The genus Laminaria sensu lato: Recent insights and developments. Eur. J. Phycol. 2008, 43, 1–86. [Google Scholar] [CrossRef]
- Tamigneaux, É.; Johnson, L.E. Les macroalgues du Saint-Laurent: Une composante essentielle d’un écosystème marin unique et une ressource naturelle précieuse dans un contexte de changement global. Nat. Can. 2016, 140, 62–73. [Google Scholar] [CrossRef]
- Charrier, B.A.; Abreu, M.H.; Araujo, R.; Bruhn, A.; Coates, J.C.; De Clerck, O.; Katsaros, C.; Robaina, R.R.; Wichard, T. Furthering knowledge of seaweed growth and development to facilitate sustainable aquaculture. New Phytol. 2017, 216, 967–975. [Google Scholar] [CrossRef]
- Forbord, S.; Matsson, S.; Brodahl, G.E.; Bluhm, B.A.; Broch, O.J.; Handå, A.; Metaxas, A.; Skjermo, J.; Steinhovden, K.B.; Olsen, Y. Latitudinal, seasonal and depth-dependent variation in growth, chemical composition and biofouling of cultivated Saccharina latissima (Phaeophyceae) along the Norwegian coast. J. Appl. Phycol. 2020, 32, 2215–2232. [Google Scholar] [CrossRef]
- Gendron, L.; Tamigneaux, É. Expériences de Culture de Ľalgue Brune Saccharina longicruris en 2007: Essais en Bassin et en Mer au Large de Paspébiac et de Grande-Rivière (Québec); Pêche et Océans Canada: Mont-Joli, QC, Canada, 2007. [Google Scholar]
- Forbord, S.; Skjermo, J.; Arff, J.; Handå, A.; Reitan, K.I.; Bjerregaard, R.; Lüning, K. Development of Saccharina latissima (Phaeophyceae) kelp hatcheries with year-round production of zoospores and juvenile sporophytes on culture ropes for kelp aquaculture. J. Appl. Phycol. 2012, 24, 393–399. [Google Scholar] [CrossRef]
- Tamigneaux, É.; Pedneault, E.; Gendron, L. Comparaison des Rendements de L’Algue brune Saccharina longicruris Cultivée en Milieu Ouvert en Gaspésie et en Lagune aux Îles-de-la-Madeleine; Merinov: Grande-Rivère, QC, Canada, 2014. [Google Scholar]
- Raven, J.A.; Johnston, A.M. Mechanisms of inorganic-carbon acquisition in marine phytoplankton and their implications for the use of other resources. Limnol. Oceanogr. 1991, 36, 1701–1714. [Google Scholar] [CrossRef]
- Beer, S.; Björk, M.; Beardall, J.; Bjork, M. Photosynthesis in the Marine Environment; John Wiley & Sons, Inc.: Hoboken, UK, 2014. [Google Scholar]
- Brody, S.R.; Lozier, M.S.; Dunne, J.P. A comparison of methods to determine phytoplankton bloom initiation. J. Geophysical. Res. Oceans 2013, 118, 2345–2357. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Effect of season on the composition of bioactive polysaccharides from the brown seaweed Saccharina longicruris. Phytochemistry 2009, 70, 1069–1075. [Google Scholar] [CrossRef]
- Vasconcelos, M.M.M.; Marson, G.V.; Turgeon, S.L.; Tamigneaux, É.; Beaulieu, L. Physicochemical properties of wild and cultivated Saccharina latissima macroalgae harvested in the Canadian boreal-subarctic transition zone. Foods 2022. submitted to foods. [Google Scholar] [CrossRef]
- Beaulieu, L.; Bondu, S.; Doiron, K.; Rioux, L.-E.; Turgeon, S.L. Characterization of antibacterial activity from protein hydrolysates of the macroalga Saccharina longicruris and identification of peptides implied in bioactivity. J. Funct. Foods 2015, 17, 685–697. [Google Scholar] [CrossRef]
- Stévant, P.; Indergård, E.; Ólafsdóttir, A.; Marfaing, H.; Larssen, W.E.; Fleurence, J.; Roleda, M.Y.; Rustad, T.; Slizyte, R.; Nordtvedt, T.S. Effects of drying on the nutrient content and physico-chemical and sensory characteristics of the edible kelp Saccharina latissima. J. Appl. Phycol. 2018, 30, 2587–2599. [Google Scholar] [CrossRef]
- Akomea-Frempong, S.; Skonberg, D.I.; Camire, M.E.; Perry, J.J. Impact of Blanching, Freezing, and Fermentation on Physicochemical, Microbial, and Sensory Quality of Sugar Kelp (Saccharina latissima). Foods 2021, 10, 2258. [Google Scholar] [CrossRef] [PubMed]
- Albers, E.; Malmhäll-Bah, E.; Olsson, J.; Sterner, M.; Mayers, J.J.; Nylund, G.M.; Rupar-Gadd, K.; Abdollahi, M.; Cvijetinovic, S.; Welander, U.; et al. Influence of preservation methods on biochemical composition and downstream processing of cultivated Saccharina latissima biomass. Algal. Res. 2021, 55, 102261. [Google Scholar] [CrossRef]
- Susanto, E.; Fahmi, A.S.; Agustini, T.W.; Rosyadi, S.; Wardani, A.D. Effects of Different Heat Processing on Fucoxanthin, Antioxidant Activity and Colour of Indonesian Brown. IOP Conf. Ser. Earth Environ. Sci. 2017, 55, 012063. [Google Scholar] [CrossRef]
- Abdollahi, M.; Axelsson, J.; Carlsson, N.-G.; Nylund, G.M.; Albers, E.; Undeland, I. Effect of stabilization method and freeze/thaw-aided precipitation on structural and functional properties of proteins recovered from brown seaweed (Saccharina latissima). Food Hydrocoll. 2019, 96, 140–150. [Google Scholar] [CrossRef]
- Obluchinskaya, E.; Daurtseva, A. Effects of air drying and freezing and long-term storage on phytochemical composition of brown seaweeds. J. Appl. Phycol. 2020, 32, 4235–4249. [Google Scholar] [CrossRef]
- Silva, C.L.; Gonçalves, E.M.; Brandao, T.R. Freezing of fruits and vegetables. In Frozen Food Science and Technology; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 165–193. [Google Scholar]
- Davison, I.R.; Ruan, S.R.; Hang-Ming, R. Effect of freezing on seaweed photosynthesis. Mar. Ecol. Prog. Ser. 1989, 58, 123–131. [Google Scholar] [CrossRef]
- Zhu, X.; Healy, L.E.; Sevindik, O.; Sun, D.W.; Selli, S.; Kelebek, H.; Tiwari, B.K. Impacts of novel blanching treatments combined with commercial drying methods on the physicochemical properties of Irish brown seaweed Alaria esculenta. Food Chem. 2022, 369, 130949. [Google Scholar] [CrossRef] [PubMed]
- Akomea-Frempong, S.; Perry, J.J.; Skonberg, D.I. Effects of pre-freezing blanching procedures on the physicochemical properties and microbial quality of frozen sugar kelp. J. Appl. Phycol. 2021, 34, 609–624. [Google Scholar] [CrossRef]
- Blikra, M.J.; Lovdal, T.; Vaka, M.R.; Roiha, I.S.; Lunestad, B.T.; Lindseth, C.; Skipnes, D. Assessment of food quality and microbial safety of brown macroalgae (Alaria esculenta and Saccharina latissima). J. Sci. Food Agric. 2019, 99, 1198–1206. [Google Scholar] [CrossRef]
- Nielsen, C.W.; Holdt, S.L.; Sloth, J.J.; Marinho, G.S.; Saether, M.; Funderud, J.; Rustad, T. Reducing the High Iodine Content of Saccharina latissima and Improving the Profile of Other Valuable Compounds by Water Blanching. Foods 2020, 9, 569. [Google Scholar] [CrossRef]
- Andersson, J.; Garrido-Bañuelos, G.; Bergdoll, M.; Vilaplana, F.; Menzel, C.; Mihnea, M.; Lopez-Sanchez, P. Comparison of steaming and boiling of root vegetables for enhancing carbohydrate content and sensory profile. J. Food Eng. 2022, 312, 110754. [Google Scholar] [CrossRef]
- Nobosse, P.; Fombang, E.N.; Mbofung, C.M.F. The effect of steam blanching and drying method on nutrients, phytochemicals and antioxidant activity of Moringa (Moringa oleifera L.) leaves. Am. J. Food Sci. Technol. 2017, 5, 53–60. [Google Scholar] [CrossRef]
- Amorim-Carrilho, K.; Lage-Yusty, M.A.; López-Hernández, J. Variation of bioactive compounds in dried seaweedHimanthalia elongatasubjected to different culinary processes. CyTA J. Food 2014, 12, 336–339. [Google Scholar] [CrossRef]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2015, 28, 511–524. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jónsdóttir, R.; Kristinsson, H.G.; Hreggvidsson, G.O.; Jónsson, J.Ó.; Thorkelsson, G.; Ólafsdóttir, G. Enzyme-enhanced extraction of antioxidant ingredients from red algae Palmaria palmata. LWT 2010, 43, 1387–1393. [Google Scholar] [CrossRef]
- Ganesan, K.; Kumar, K.S.; Rao, P.V.S. Comparative assessment of antioxidant activity in three edible species of green seaweed, Enteromorpha from Okha, Northwest coast of India. Innov. Food Sci. Emerg. Technol. 2011, 12, 73–78. [Google Scholar] [CrossRef]
- Singleton, V.L.R.; Joseph, A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Abdelhady, M.I.S.; Motaal, A.A.; Beerhues, L. Total Phenolic Content and Antioxidant Activity of Standardized Extracts from Leaves and Cell Cultures of Three Callistemon Species. Am. J. Plant Sci. 2011, 02, 847–850. [Google Scholar] [CrossRef]
- Quan, C.; Turner, C. Extraction of Astaxanthin from Shrimp Waste Using Pressurized Hot Ethanol. Chromatographia 2009, 70, 247–251. [Google Scholar] [CrossRef]
- Fletcher, H.R.; Biller, P.; Ross, A.B.; Adams, J.M.M. The seasonal variation of fucoidan within three species of brown macroalgae. Algal. Res. 2017, 22, 79–86. [Google Scholar] [CrossRef]
- Honya, M.; Kinoshita, T.; Ishikawa, M.; Mori, H.; Nisizawa, K. Monthly Determination of Alginate, M/G Ratio, Mannitol, and Minerals in Cultivated Laminaria japonica. Nippon. Suisan Gakkaishi 1993, 59, 295–299. [Google Scholar] [CrossRef]
- Bondu, S.; Bonnet, C.; Gaubert, J.; Deslandes, É.; Turgeon, S.L.; Beaulieu, L. Bioassay-guided fractionation approach for determination of protein precursors of proteolytic bioactive metabolites from macroalgae. J. Appl. Phycol. 2014, 27, 2059–2074. [Google Scholar] [CrossRef]
- Beaulieu, L.; Sirois, M.; Tamigneaux, É. Evaluation of the in vitro biological activity of protein hydrolysates of the edible red alga, Palmaria palmata (dulse) harvested from the Gaspe coast and cultivated in tanks. J. Appl. Phycol. 2016, 28, 3101–3115. [Google Scholar] [CrossRef]
- Simonne, A.H.; Simonne, E.H.; Eitenmiller, R.R.; Mills, H.A.; Cresman, C.P. Could the Dumas Method replace the Kjeldahl digestion for nitrogen and crude protein determinations in foods? J. Sci. Food Agric. 1997, 73, 39–45. [Google Scholar] [CrossRef]
- Cao, G.A.; Helaine, M.; Cutler, R.G. Oxygen-radical absorbance capacity assay for antioxidants. Free Radic. Biol. Med. 1993, 14, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Kelman, D.; Posner, E.K.; McDermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant activity of Hawaiian marine algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef]
- Reissiger, S.M. Characterization of Three Macroalgae: Saccharina Latissima, Alaria Esculenta and Palmaria Palmata. Master’s Thesis, Norwegian University of Science and Technology, Trondheim, Norway, 2016. [Google Scholar]
- Handå, A.; Forbord, S.; Wang, X.; Broch, O.J.; Dahle, S.W.; Størseth, T.R.; Reitan, K.I.; Olsen, Y.; Skjermo, J. Seasonal- and depth-dependent growth of cultivated kelp (Saccharina latissima) in close proximity to salmon (Salmo salar) aquaculture in Norway. Aquaculture 2013, 414–415, 191–201. [Google Scholar] [CrossRef]
- Nielsen, M.M.; Krause-Jensen, D.; Olesen, B.; Thinggaard, R.; Christensen, P.B.; Bruhn, A. Growth dynamics of Saccharina latissima (Laminariales, Phaeophyceae) in Aarhus Bay, Denmark, and along the species’ distribution range. Mar. Biol. 2014, 161, 2011–2022. [Google Scholar] [CrossRef]
- Jordbrekk Blikra, M.; Wang, X.; James, P.; Skipnes, D. Saccharina latissima Cultivated in Northern Norway: Reduction of Potentially Toxic Elements during Processing in Relation to Cultivation Depth. Foods 2021, 10, 1290. [Google Scholar] [CrossRef] [PubMed]
- Trigo, J.P.; Stedt, K.; Schmidt, A.E.M.; Kollander, B.; Edlund, U.; Nylund, G.; Pavia, H.; Abdollahi, M.; Undeland, I. Mild blanching prior to pH-shift processing of Saccharina latissima retains protein extraction yields and amino acid levels of extracts while minimizing iodine content. Food Chem. 2023, 404, 134576. [Google Scholar] [CrossRef] [PubMed]
- Venturi, S. Evolutionary Significance of Iodine. Curr. Chem. Biol. 2011, 5, 155–162. [Google Scholar]
- Municipality-Canada Paspébiac. Available online: https://www.municipality-canada.com/en/ville-paspebiac.html (accessed on 11 March 2023).
- OGSL. Conditions Maritimes. Available online: https://ogsl.ca/conditions/?lg=fr (accessed on 11 March 2023).
- Para, J.; Coble, P.G.; Charrière, B.; Tedetti, M.; Fontana, C.; Sempéré, R. Fluorescence and absorption properties of chromophoric dissolved organic matter (CDOM) in coastal surface waters of the northwestern Mediterranean Sea, influence of the Rhône River. Biogeosciences 2010, 7, 4083–4103. [Google Scholar] [CrossRef]
- Chen, J.; Ye, W.; Guo, J.; Luo, Z.; Li, Y. Diurnal Variability in Chlorophyll-a, Carotenoids, CDOM and SO42− Intensity of Offshore Seawater Detected by an Underwater Fluorescence-Raman Spectral System. Sensors 2016, 16, 1082. [Google Scholar] [CrossRef]
- Ji, R.; Edwards, M.; Mackas, D.L.; Runge, J.A.; Thomas, A.C. Marine plankton phenology and life history in a changing climate: Current research and future directions. J. Plankton Res. 2010, 32, 1355–1368. [Google Scholar] [CrossRef]
- Fichier Canadien sur les Éléments Nutritifs, Spinach, Boiled, Drained, with Salt, food code: 6544. Available online: https://aliments-nutrition.canada.ca/cnf-fce/report-rapport (accessed on 11 March 2023).
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef] [PubMed]
- Chupeerach, C.; Aursalung, A.; Watcharachaisoponsiri, T.; Whanmek, K.; Thiyajai, P.; Yosphan, K.; Sritalahareuthai, V.; Sahasakul, Y.; Santivarangkna, C.; Suttisansanee, U. The Effect of Steaming and Fermentation on Nutritive Values, Antioxidant Activities, and Inhibitory Properties of Tea Leaves. Foods 2021, 10, 117. [Google Scholar] [CrossRef] [PubMed]
- Ciqual. Califlower Steamed. Available online: https://ciqual.anses.fr/#/aliments/20312/cauliflower-steamed (accessed on 11 March 2023).
- Lytou, A.E.; Schoina, E.; Liu, Y.; Michalek, K.; Stanley, M.S.; Panagou, E.Z.; Nychas, G.E. Quality and Safety Assessment of Edible Seaweeds Alaria esculenta and Saccharina latissima Cultivated in Scotland. Foods 2021, 10, 2210. [Google Scholar] [CrossRef] [PubMed]
- Lafeuille, B.; Francezon, N.; Goulet, C.; Perreault, V.; Turgeon, S.L.; Beaulieu, L. Impact of temperature and cooking time on the physicochemical properties and sensory potential of seaweed water extracts of Palmaria palmata and Saccharina longicruris. J. Appl. Phycol. 2022, 34, 1731–1747. [Google Scholar] [CrossRef]
- Ciqual. Lentils Dried. Available online: https://ciqual.anses.fr/#/aliments/20504/lentille-seche (accessed on 11 March 2023).
- Ciqual. Wheat Bran. Available online: https://ciqual.anses.fr/#/aliments/9621/son-de-ble (accessed on 11 March 2023).
- Fonseca, I.; Guarda, I.; Mourato, M.; Martins, L.L.; Gomes, R.; Matos, J.; Gomes-Bispo, A.; Bandarra, N.M.; Cardoso, C.; Afonso, C. Undervalued Atlantic brown seaweed species (Cystoseira abies-marina and Zonaria tournefortii): Influence of treatment on their nutritional and bioactive potential and bioaccessibility. Euro. Food Res. Technol. 2020, 247, 221–232. [Google Scholar] [CrossRef]
- Parjikolaei, B.R.; Bruhn, A.; Eybye, K.L.; Larsen, M.M.; Rasmussen, M.B.; Christensen, K.V.; Fretté, X.C. Valuable Biomolecules from Nine North Atlantic Red Macroalgae: Amino Acids, Fatty Acids, Carotenoids, Minerals and Metals. Nat. Resour. 2016, 7, 157–183. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Frediansyah, A.; Martyasari, N.W.R.; Ilhami, B.K.; Abidin, A.S.; Padmi, H.; Fahrurrozi; Juanssilfero, A.; Widyastuti, S.; Sunarwidhi, A.L. Effect of particle size on phytochemical composition and antioxidant properties of Sargassum cristaefolium ethanol extract. Sci. Rep. 2021, 11, 17876. [Google Scholar] [CrossRef]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Galland-Irmouli, A.; Fleurence, J.; Lamghari, R.; Luçon, L.; Rouxel, C.B.O.; Bronowicki, J.V.C.; Guéant, J. Nutritional value of proteins from edible seaweed Palmaria palmata (dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Jimenez-Escrig, A.; Gomez-Ordonez, E.; Ruperez, P. Seaweed as a source of novel nutraceuticals: Sulfated polysaccharides and peptides. Adv. Food Nutr. Res. 2011, 64, 325–337. [Google Scholar] [CrossRef]
- Stefaniak, M.M.; Gudjónsdóttir, M.; Marteinsdottir, G.; Omarsdottir, S.; Bravo, E.; Sigurjónsson, O.E.; Kristbergsson, K. Determination of bioactive properties of food grade extracts from Icelandic edible brown seaweed sugar kelp (Saccharina latissima) with in vitro human cell cultures (THP-1). Funct. Foods Health Dis. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, N.; Ghosh, T.; Sinha, S.; Chattopadhyay, K.; Karmakar, P.; Ray, B. Polysaccharides from Turbinaria conoides: Structural features and antioxidant capacity. Food Chem. 2010, 118, 823–829. [Google Scholar] [CrossRef]
- Dudonné, S.; Vitrac, X.; Coutière, P.; Woillez, M.; Mérillon, J. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agr. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Maehre, H.K.; Edvinsen, G.K.; Eilertsen, K.-E.; Elvevoll, E.O. Heat treatment increases the protein bioaccessibility in the red seaweed dulse (Palmaria palmata), but not in the brown seaweed winged kelp (Alaria esculenta). J. Appl. Phycol. 2015, 28, 581–590. [Google Scholar] [CrossRef]
- Rajauria, G.; Jaiswal, A.K.; Abu-Ghannam, N.; Gupta, S. Effect of hydrothermal processing on colour, antioxidant and free radical scavenging capacities of edible Irish brown seaweeds. Int. J. Food Sci. Technol. 2010, 45, 2485–2493. [Google Scholar] [CrossRef]
- Ghaliaoui, N.M.; Hazzit, H.; Hadjadj, M.; Otmani, M.; Touati, F.S.; Halima, S.S. Impact of Freezing and Drying Preprocessing on Pigments Extraction from the Brown Seaweed« Phyllaria Reniformis» Collected in Algerian Coast. Carpathian J. Food Sci. Technol. 2020, 12, 81–94. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl Treatment and Harvest Time | Lipids (%) | Proteins (%) | Ash (%) | Carbohydrates (%) | Fibers (%) |
|---|---|---|---|---|---|
| Crudes–May | 1.88 ± 0.02 ab | 11.00 ± 0.14 d | 43.06 ± 0.07 a | 44.05 ± 0.29 e | 34.57 ± 0.18 a |
| Blanched–May | 1.59 ± 0.22 ab | 15.46 ± 0.21 a | 21.67 ± 0.01 f | 61.28 ± 0.26 b | 54.77 ± 1.13 a |
| Steamed–May | 1.40 ± 0.19 b | 14.66 ± 0.25 b | 21.88 ± 0.18 f | 62.06 ± 0.27 b | 52.24 ± 0.50 a |
| Dried–May | 1.80 ± 0.24 ab | 10.38 ± 0.39 d | 42.96 ± 0.15 a | 44.86 ± 0.44 e | 34.78 ± 0.30 a |
| Crudes–June | 1.63 ± 0.16 ab | 9.00 ± 0.32 e | 41.55 ± 0.01 b | 47.82 ± 0.52 d | 37.72 ± 0.36 a |
| Blanched–June | 0.69 ± 0.04 c | 13.16 ± 0.47 c | 27.53 ± 0.03 d | 58.62 ± 0.67 c | 53.50 ± 1.17 a |
| Steamed–June | 1.30 ± 0.26 b | 13.13 ± 0.11 c | 22.24 ± 0.17 e | 63.34 ± 0.09 a | 57.12 ± 0.90 a |
| Dried-June | 1.98 ± 0.22 a | 9.42 ± 0.11 e | 40.09 ± 0.11 c | 48.51 ± 0.42 d | 35.94 ± 0.32 a |
| Sl Treatment and Harvest Time | I (g/100 g) | K (g/100 g) | Na (g/100 g) | Ca (mg/100 g) | Mg (mg/100 g) | Fe (mg/100 g) |
|---|---|---|---|---|---|---|
| Crudes–May | 0.20 ± 0.00 ab | 15.74 ± 1.28 a | 2.33 ± 0.06 c | 614.00 ± 22.00 ab | 623.00 ± 23.00 bc | 41.67 ± 2.89 a |
| Blanched–May | 0.01 ± 0.00 e | 0.95 ± 0.01 c | 6.20 ± 0.20 b | 744.00 ± 6.00 ab | 207.00 ± 4.00 f | 19.33 ± 1.53 c |
| Steamed–May | 0.06 ± 0.00 d | 5.51 ± 0.09 abc | 1.03 ± 0.12 d | 976.00 ± 79.00 a | 634.00 ± 46.00 bc | 31.67 ± 1.53 b |
| Dried–May | 0.19 ± 0.00 b | 12.70 ± 0.43 abc | 2.23 ± 0.06 c | 543.00 ± 15.00 bc | 532.00 ± 14.00 e | 34.33 ± 2.52 b |
| Crudes–June | 0.17 ± 0.01 c | 11.15 ± 0.13 abc | 2.57 ± 0.06 c | 638.00 ± 16.00 ab | 608.00 ± 2.00 cd | 12.33 ± 0.58 e |
| Blanched–June | 0.02 ± 0.00 e | 1.77 ± 0.4 bc | 6.97 ± 0.38 a | 626.00 ± 19.00 ab | 181.00 ± 7.00 g | 12.00 ± 0.00 e |
| Steamed–June | 0.07 ± 0.00 d | 5.20 ± 0.09 abc | 0.97 ± 0.06 d | 922.00 ± 4.00 a | 505.00 ± 27.00 e | 13.67 ± 0.58 de |
| Dried-June | 0.21 ± 0.01 a | 10.95 ± 0.21 abc | 2.27 ± 0.29 d | 641.00 ± 9.00 ab | 547.00 ± 15.00 de | 15.33 ± 0.58 d |
| Sl Treatment and Harvest Time | Alginates (g/100 g) | Fucoidans (g/100 g) | Laminarans (g/100 g) | Carotenoids (μg/g) | Polyphenols (mg GA/g) |
|---|---|---|---|---|---|
| Crudes–May | 12.53 ± 2.07 b | 5.47 ± 0.49 de | 3.60 ± 0.44 bc | 558.00 ± 21.00 a | 3.77 ± 0.06 ab |
| Blanched–May | 30.73 ± 1.50 a | 8.13 ± 0.76 ab | 3.73 ± 0.29 bc | 323.00 ± 1.00 de | 1.73 ± 0.12 e |
| Steamed–May | 27.77 ± 1.92 a | 8.13 ± 0.80 ab | 2.80 ± 0.53 cd | 555.00 ± 24.00 a | 2.85 + 0.07 cd |
| Dried–May | 12.57 ± 2.11 b | 6.07 ± 0.75 cd | 4.47 ± 0.21 ab | 405.00 ± 11.00 c | 2.60 ± 0.20 cd |
| Crudes–June | 15.97 ± 1.41 b | 3.83 ± 0.25 e | 4.00 ± 0.10 b | 466.00 ± 14.00 b | 3.80 ± 0.20 ab |
| Blanched–June | 28.30 ± 1.74 a | 7.30 ± 0.44 bc | 1.87 ± 0.67 de | 281.00 ± 16.00 e | 3.20 ± 0.35 bc |
| Steamed–June | 28.17 ± 0.12 a | 9.50 ± 0.61 a | 1.07 ± 0.15 e | 337.00 ± 17.00 d | 4.33 ± 0.32 a |
| Dried-June | 15.47 ± 2.14 b | 4.30 ± 0.40 e | 3.90 ± 2.27 b | 330.00 ± 28.00 de | 2.33 ± 0.32 de |
| Sl Extracts | Crudes–May | Blanched–May | Steamed–May | Dried–May | Crudes–June | Blanched–June | Steamed–June | Dried–June |
|---|---|---|---|---|---|---|---|---|
| Proteins (%) | 40.68 ± 1.43 ab | 32.76 ± 1.56 d | 37.57 ± 0.89 bc | 42.91 ± 0.15 a | 36.87 ± 1.20 c | 35.78 ± 1.63 cd | 38.50 ± 1.04 bc | 38.27 ± 1.57 bc |
| Extraction yield (%) | 1.68 ± 0.28 a | 1.33 ± 0.16 ab | 1.40 ± 0.24 ab | 0.84 ± 0.10 bc | 0.91 ± 0.18 bd | 1.51 ± 0.22 a | 0.69 ± 0.24 c | 1.59 ± 0.06 a |
| Protein extraction yield (%) | 6.55 ± 1.11 a | 2.98 ± 0.23 bc | 3.76 ± 0.69 bc | 3.78 ± 0.32 bc | 4.09 ± 0.95 bc | 4.40 ± 0.73 b | 2.19 ± 0.71 c | 6.82 ± 0.48 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lafeuille, B.; Tamigneaux, É.; Berger, K.; Provencher, V.; Beaulieu, L. Variation of the Nutritional Composition and Bioactive Potential in Edible Macroalga Saccharina latissima Cultivated from Atlantic Canada Subjected to Different Growth and Processing Conditions. Foods 2023, 12, 1736. https://doi.org/10.3390/foods12081736
Lafeuille B, Tamigneaux É, Berger K, Provencher V, Beaulieu L. Variation of the Nutritional Composition and Bioactive Potential in Edible Macroalga Saccharina latissima Cultivated from Atlantic Canada Subjected to Different Growth and Processing Conditions. Foods. 2023; 12(8):1736. https://doi.org/10.3390/foods12081736
Chicago/Turabian StyleLafeuille, Bétina, Éric Tamigneaux, Karine Berger, Véronique Provencher, and Lucie Beaulieu. 2023. "Variation of the Nutritional Composition and Bioactive Potential in Edible Macroalga Saccharina latissima Cultivated from Atlantic Canada Subjected to Different Growth and Processing Conditions" Foods 12, no. 8: 1736. https://doi.org/10.3390/foods12081736
APA StyleLafeuille, B., Tamigneaux, É., Berger, K., Provencher, V., & Beaulieu, L. (2023). Variation of the Nutritional Composition and Bioactive Potential in Edible Macroalga Saccharina latissima Cultivated from Atlantic Canada Subjected to Different Growth and Processing Conditions. Foods, 12(8), 1736. https://doi.org/10.3390/foods12081736