

The Molecular Mechanism Underlying the Therapeutic Effect of Dihydromyricetin on Type 2 Diabetes Mellitus Based on Network Pharmacology, Molecular Docking, and Transcriptomics


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Network Pharmacology Analysis
2.2.1. Prediction of DHM Targets
2.2.2. Prediction of Targets of T2DM
2.2.3. PPI Network Construction and Its Visualization
2.2.4. GO Enrichment and KEGG Pathway Analysis
2.2.5. Molecular Docking
2.3. Animals and Models
2.4. Transcriptome Sequencing Analysis
2.5. qRT-PCR
2.6. Western Blotting
2.7. Statistical Analysis
3. Results
3.1. Prediction of DHM and T2DM Targets
3.2. PPI Network Construction and Its Visualization
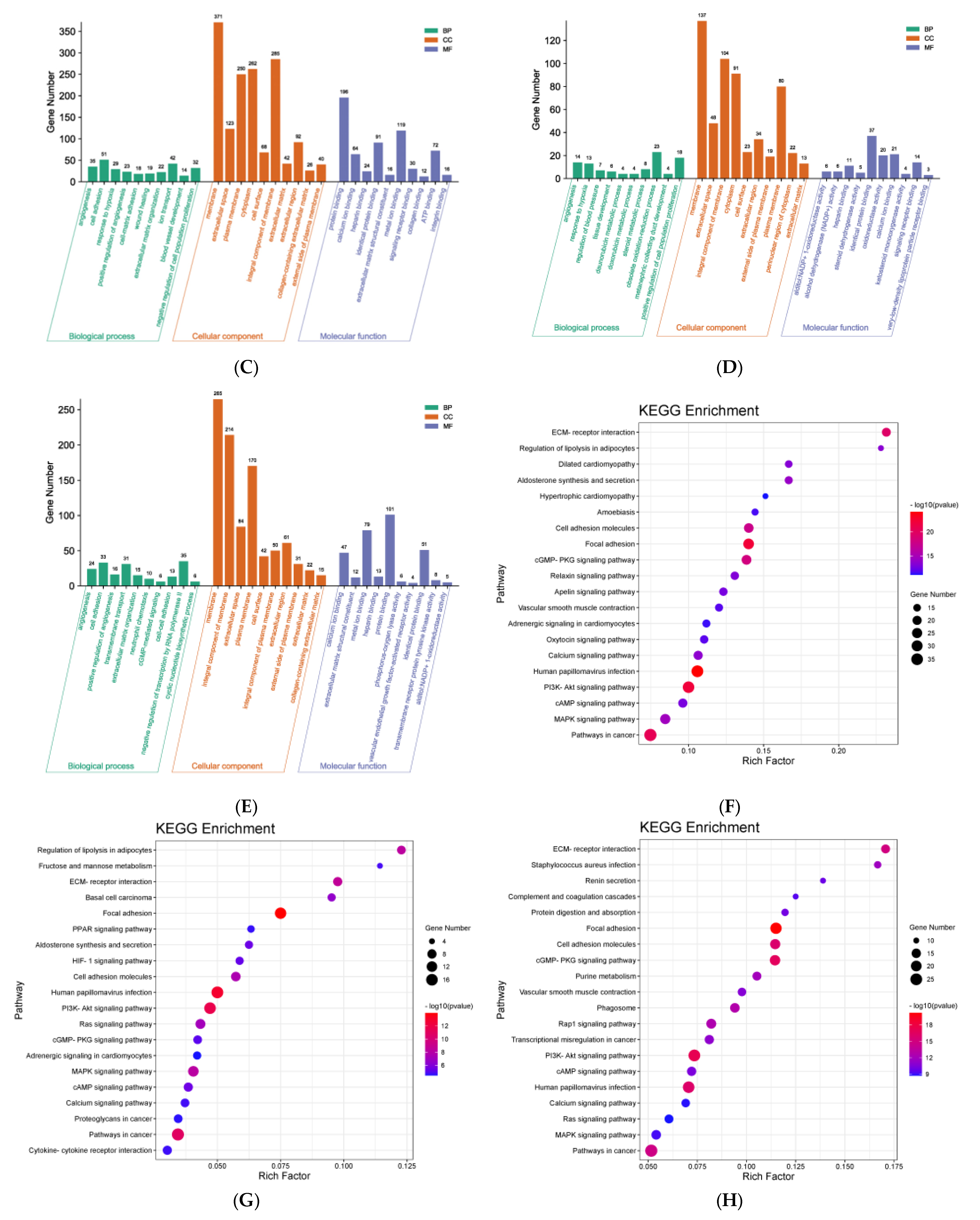
3.3. GO Enrichment and KEGG Pathway Analysis Based on Network Pharmacology
3.4. Construction and Analysis of “Composition-Target-Disease-Pathway” Network
3.5. Molecular Docking
3.6. Effects of DHM on Physiological and Biochemical Indicators of db/db Mice
3.7. Effect of DHM on Gene Expression of Liver in db/db Mice
3.8. GO Enrichment and KEGG Pathway Analysis Based on Liver Transcriptomes
3.9. qRT-PCR Analysis
3.10. Effect of DHM on PI3K/AKT/MAPK Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; Available online: https://www.diabetesatlas.org (accessed on 18 May 2023).
- Li, H.; Chang, G.Y.; Jiang, Y.H.; Xu, L.; Shen, L.; Gu, Z.C.; Lin, H.W.; Shi, F.H. System Dynamic Model Simulates the Growth Trend of Diabetes Mellitus in Chinese Population: Implications for Future Urban Public Health Governance. Int. J. Public Health 2022, 67, 1605064. [Google Scholar] [CrossRef] [PubMed]
- Classification of Diabetes Mellitus; World Health Organization: Geneva, Switzerland, 2019.
- Li, Y.; Teng, D.; Shi, X.; Qin, G.; Qin, Y.; Quan, H.; Shi, B.; Sun, H.; Ba, J.; Chen, B.; et al. Prevalence of diabetes recorded in mainland China using 2018 diagnostic criteria from the American Diabetes Association: National cross-sectional study. BMJ 2020, 369, m997. [Google Scholar] [CrossRef] [PubMed]
- Association, A.D. Classification and diagnosis of diabetes: Standards of medical care in Diabetesd 2018. Diabetes Care 2018, 41, 13–27. [Google Scholar] [CrossRef]
- Ortiz-Martínez, M.; González-González, M.; Martagón, A.J.; Hlavinka, V.; Willson, R.C.; Rito-Palomares, M. Recent Developments in Biomarkers for Diagnosis and Screening of Type 2 Diabetes Mellitus. Curr. Diabetes Rep. 2022, 22, 95–115. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Rahu, N.; Saleem Kalhoro, M.; Hussain Kalhoro, D.; Adebowale, T.O.; Usman Mazhar, M.; Rehman, Z.U.; et al. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacol. Res. 2020, 152, 104629. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Bhat, J.; Wang, P.H. Cardiovascular effects of anti-diabetic medications in type 2 diabetes mellitus. Curr. Cardiol. Rep. 2013, 15, 327. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mittal, A.; Babu, D.; Mittal, A. Herbal Medicines for Diabetes Management and its Secondary Complications. Curr. Diabetes Rev. 2021, 17, 437–456. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Q.; Liu, Z.; Yang, K.; Chen, Z.; Cheng, Q.; Wu, L. The Versatile Effects of Dihydromyricetin in Health. Evid.-Based Complement. Altern. Med. 2017, 2017, 1053617. [Google Scholar] [CrossRef]
- He, J.; Zhang, J.; Dong, L.; Dang, X.; Wang, L.; Cheng, L.; Huang, Y. Dihydromyricetin Attenuates Metabolic Syndrome and Improves Insulin Sensitivity by Upregulating Insulin Receptor Substrate-1 (Y612) Tyrosine Phosphorylation in db/db Mice. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2237–2249. [Google Scholar] [CrossRef]
- Zhou, X.; Song, Y.; Zeng, C.; Zhang, H.; Lv, C.; Shi, M.; Qin, S. Molecular Mechanism Underlying the Regulatory Effect of Vine Tea on Metabolic Syndrome by Targeting Redox Balance and Gut Microbiota. Front. Nutr. 2022, 9, 802015. [Google Scholar] [CrossRef]
- Liu, L.; Sun, S.; Li, X. A network pharmacology-based approach to explore the effect of dihydromyricetin on non-alcoholic fatty liver rats via regulating PPARG and CASP3. Mol. Cell. Probes 2023, 71, 101926. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Chen, L.; Lin, S.; Cao, H.; Teng, H. A designed self-microemulsion delivery system for dihydromyricetin and its dietary intervention effect on high-fat-diet fed mice. Food Chem. 2022, 390, 132954. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.P.; Wang, Q.H.; Zheng, H.J.; Wei, F. Research Progress on Non-Drug Treatment for Blood Glucose Control of Type 2 Diabetes Mellitus. Chin. J. Integr. Med. 2018, 24, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Egbuna, C.; Awuchi, C.G.; Kushwaha, G.; Rudrapal, M.; Patrick-Iwuanyanwu, K.C.; Singh, O.; Odoh, U.E.; Khan, J.; Jeevanandam, J.; Kumarasamy, S.; et al. Bioactive Compounds Effective Against Type 2 Diabetes Mellitus: A Systematic Review. Curr. Top. Med. Chem. 2021, 21, 1067–1095. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Xiang, H.; Ding, P.; Wu, T.; Ji, G. Recent update on application of dihydromyricetin in metabolic related diseases. Biomed. Pharmacother. 2022, 148, 112771. [Google Scholar] [CrossRef] [PubMed]
- White, A.L.; Bix, G.J. VEGFA Isoforms as Pro-Angiogenic Therapeutics for Cerebrovascular Diseases. Biomolecules 2023, 13, 702. [Google Scholar] [CrossRef]
- Alimam, H.Y.; Hussein, W.A.; Ibrahim, S.; Abdelgani, S.; Alharthi, N.; Eltayeb, L.B.; Elmahdi, S.A.; Abdrabo, A.A. Blood Glucose, HbA1c Level, and its Correlation with VEGF-A (+405G/C) Polymorphism as Biomarker Predicts the Risk of Retinopathy and Nephropathy in Type 2 Diabetic Patients. Rep. Biochem. Mol. Biol. 2022, 11, 421–429. [Google Scholar] [CrossRef]
- Cuadrado, A.; Nebreda, A.R. Mechanisms and functions of p38 MAPK signalling. Biochem. J. 2010, 429, 403–417. [Google Scholar] [CrossRef]
- Bengal, E.; Aviram, S.; Hayek, T. p38 MAPK in Glucose Metabolism of Skeletal Muscle: Beneficial or Harmful? Int. J. Mol. Sci. 2020, 21, 6480. [Google Scholar] [CrossRef]
- Zhao, Y.; Ye, W.; Wang, Y.D.; Chen, W.D. HGF/c-Met: A Key Promoter in Liver Regeneration. Front. Pharmacol. 2022, 13, 808855. [Google Scholar] [CrossRef]
- Ilegems, E.; Bryzgalova, G.; Correia, J.; Yesildag, B.; Berra, E.; Ruas, J.L.; Pereira, T.S.; Berggren, P.O. HIF-1α inhibitor PX-478 preserves pancreatic β cell function in diabetes. Sci. Transl. Med. 2022, 14, eaba9112. [Google Scholar] [CrossRef] [PubMed]
- Le, L.; Jiang, B.; Wan, W.; Zhai, W.; Xu, L.; Hu, K.; Xiao, P. Metabolomics reveals the protective of Dihydromyricetin on glucose homeostasis by enhancing insulin sensitivity. Sci. Rep. 2016, 6, 36184. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Qian, D.W.; Jiang, S.; Shang, E.X.; Zhu, Z.H.; Duan, J.A. Scutellariae Radix and Coptidis Rhizoma Improve Glucose and Lipid Metabolism in T2DM Rats via Regulation of the Metabolic Profiling and MAPK/PI3K/Akt Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 3634. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Zaib, S.; Rahman, M.M.; Jannat, S.; Iqbal, J.; Park, S.K.; Chang, M.S. Didymin, a dietary citrus flavonoid exhibits anti-diabetic complications and promotes glucose uptake through the activation of PI3K/Akt signaling pathway in insulin-resistant HepG2 cells. Chem. Biol. Interact. 2019, 305, 180–194. [Google Scholar] [CrossRef]
- Li, J.S.; Ji, T.; Su, S.L.; Zhu, Y.; Chen, X.L.; Shang, E.X.; Guo, S.; Qian, D.W.; Duan, J.A. Mulberry leaves ameliorate diabetes via regulating metabolic profiling and AGEs/RAGE and p38 MAPK/NF-κB pathway. J. Ethnopharmacol. 2022, 283, 114713. [Google Scholar] [CrossRef]
- Lee, H.H.; Kim, K.J.; Lee, O.H.; Lee, B.Y. Effect of pycnogenol on glucose transport in mature 3T3-L1 adipocytes. Phytother. Res. 2010, 24, 1242–1249. [Google Scholar] [CrossRef]
- Ke, Y.Q.; Liu, C.; Hao, J.B.; Lu, L.; Lu, N.N.; Wu, Z.K.; Zhu, S.S.; Chen, X.L. Morin inhibits cell proliferation and fibronectin accumulation in rat glomerular mesangial cells cultured under high glucose condition. Biomed. Pharmacother. 2016, 84, 622–627. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef]
- Entezari, M.; Hashemi, D.; Taheriazam, A.; Zabolian, A.; Mohammadi, S.; Fakhri, F.; Hashemi, M.; Hushmandi, K.; Ashrafizadeh, M.; Zarrabi, A.; et al. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: A pre-clinical and clinical investigation. Biomed. Pharmacother. 2022, 146, 112563. [Google Scholar] [CrossRef]
- Solinas, G.; Becattini, B. PI3K and AKT at the Interface of Signaling and Metabolism. Curr. Top. Microbiol. Immunol. 2022, 436, 311–336. [Google Scholar] [CrossRef]
- Hu, X.; Wang, S.; Xu, J.; Wang, D.B.; Chen, Y.; Yang, G.Z. Triterpenoid saponins from Stauntonia chinensis ameliorate insulin resistance via the AMP-activated protein kinase and IR/IRS-1/PI3K/Akt pathways in insulin-resistant HepG2 cells. Int. J. Mol. Sci. 2014, 15, 10446–10458. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal. Transduct. Res. 2015, 35, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, N.; Barma, S.; Konwar, N.; Dewanjee, S.; Manna, P. Mechanistic insight of diabetic nephropathy and its pharmacotherapeutic targets: An update. Eur. J. Pharmacol. 2016, 791, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, T.; Zhou, Y.; Zeng, X.; Ran, L.; Zhang, Q.; Zhu, J.; Mi, M. Dihydromyricetin improves skeletal muscle insulin sensitivity by inducing autophagy via the AMPK-PGC-1α-Sirt3 signaling pathway. Endocrine 2015, 50, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Tan, K.; Luo, Q.; Bai, X. Dihydromyricetin promotes autophagy and attenuates renal interstitial fibrosis by regulating miR-155-5p/PTEN signaling in diabetic nephropathy. Bosn. J. Basic Med. Sci. 2020, 20, 372–380. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, M.; Lang, H.; Zhou, Y.; Mi, M. Dihydromyricetin enhances glucose uptake by inhibition of MEK/ERK pathway and consequent down-regulation of phosphorylation of PPARγ in 3T3-L1 cells. J. Cell. Mol. Med. 2018, 22, 1247–1256. [Google Scholar] [CrossRef]
- Hua, Y.Y.; Zhang, Y.; Gong, W.W.; Ding, Y.; Shen, J.R.; Li, H.; Chen, Y.; Meng, G.L. Dihydromyricetin Improves Endothelial Dysfunction in Diabetic Mice via Oxidative Stress Inhibition in a SIRT3-Dependent Manner. Int. J. Mol. Sci. 2020, 21, 6699. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG ID | Pathway | Genes | p-Value | Fold Enrichment |
|---|---|---|---|---|
| hsa01522 | Endocrine resistance | SRC, MMP2, BCL2, MAPK14, ESR1, MMP9, ESR2 | 1.21 × 10−06 | 18.79262673 |
| hsa05418 | Fluid shear stress and atherosclerosis | SRC, MMP2, BCL2, KDR, MAPK14, MMP9, VEGFA | 9.33 × 10−06 | 13.24947784 |
| hsa04014 | Ras signaling pathway | PLA2G1B, KIT, PLA2G2A, KDR, MET, FGFR1, VEGFA | 1.78 × 10−04 | 7.836925189 |
| hsa04010 | MAPK signaling pathway | KIT, KDR, MAPT, MAPK14, MET, FGFR1, VEGFA | 5.96 × 10−04 | 6.264208909 |
| hsa04933 | AGE-RAGE signaling pathway in diabetic complications | STAT1, MMP2, SERPINE1, BCL2, MAPK14, VEGFA | 2.80 × 10−05 | 15.78580645 |
| hsa04151 | PI3K-Akt signaling pathway | KIT, BCL2, KDR, MET, FGFR1, VEGFA | 0.008711778 | 4.459267359 |
| hsa05417 | Lipid and atherosclerosis | SRC, BCL2, PPARG, MAPK14, MMP9 | 0.007515789 | 6.118529632 |
| hsa05208 | Chemical carcinogenesis—reactive oxygen species | SRC, MAPK14, HIF1A, MET, VEGFA | 0.008531633 | 5.899030812 |
| hsa04370 | VEGF signaling pathway | SRC, KDR, MAPK14, VEGFA | 0.001270708 | 17.83706944 |
| hsa04066 | HIF-1 signaling pathway | SERPINE1, BCL2, HIF1A, VEGFA | 0.007251828 | 9.654927493 |
| hsa05415 | Diabetic cardiomyopathy | MMP2, MAPK14, PPARA, MMP9 | 0.037668717 | 5.184172891 |
| hsa00590 | Arachidonic acid metabolism | PLA2G1B, PLA2G2A, PTGS1 | 0.020925862 | 12.93918562 |
| hsa04657 | IL-17 signaling pathway | MMP13, MAPK14, MMP9 | 0.046394807 | 8.396705559 |
| hsa00564 | Glycerophospholipid metabolism | ACHE, PLA2G1B, PLA2G2A | 0.049997898 | 8.053982883 |
| hsa04668 | TNF signaling pathway | MMP14, MAPK14, MMP9 | 0.063365979 | 7.047235023 |
| Composition | Targets | PBDID | Affinity (kcal/mol) |
|---|---|---|---|
| DHM | VEGFA | 1VPF | −7.0 |
| SRC | 4M4Z | −6.4 | |
| MAPK14 | 5ETI | −9.0 | |
| MMP9 | 5TH9 | −10.2 | |
| MET | 6I04 | −7.9 | |
| HIF1A | 1h2k | −7.5 | |
| KDR | 1ywn | −7.5 | |
| PPARG | 2VV2 | −8.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, X.; Lv, C.; Zhou, R.; Wang, Y.; Zhou, X.; Qin, S. The Molecular Mechanism Underlying the Therapeutic Effect of Dihydromyricetin on Type 2 Diabetes Mellitus Based on Network Pharmacology, Molecular Docking, and Transcriptomics. Foods 2024, 13, 344. https://doi.org/10.3390/foods13020344
Wen X, Lv C, Zhou R, Wang Y, Zhou X, Qin S. The Molecular Mechanism Underlying the Therapeutic Effect of Dihydromyricetin on Type 2 Diabetes Mellitus Based on Network Pharmacology, Molecular Docking, and Transcriptomics. Foods. 2024; 13(2):344. https://doi.org/10.3390/foods13020344
Chicago/Turabian StyleWen, Xinnian, Chenghao Lv, Runze Zhou, Yixue Wang, Xixin Zhou, and Si Qin. 2024. "The Molecular Mechanism Underlying the Therapeutic Effect of Dihydromyricetin on Type 2 Diabetes Mellitus Based on Network Pharmacology, Molecular Docking, and Transcriptomics" Foods 13, no. 2: 344. https://doi.org/10.3390/foods13020344
APA StyleWen, X., Lv, C., Zhou, R., Wang, Y., Zhou, X., & Qin, S. (2024). The Molecular Mechanism Underlying the Therapeutic Effect of Dihydromyricetin on Type 2 Diabetes Mellitus Based on Network Pharmacology, Molecular Docking, and Transcriptomics. Foods, 13(2), 344. https://doi.org/10.3390/foods13020344