Volatile Transference and Antimicrobial Activity of Cheeses Made with Ewes’ Milk Fortified with Essential Oils

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and EO Production
2.2. Milk Samples
2.3. Microbial Strains
2.4. Elaboration of Cheese Samples Fortified with EOs
2.5. Volatile Extractions and HS-SBSE/GC/MS Analyses
2.6. Cheese Microbial Content
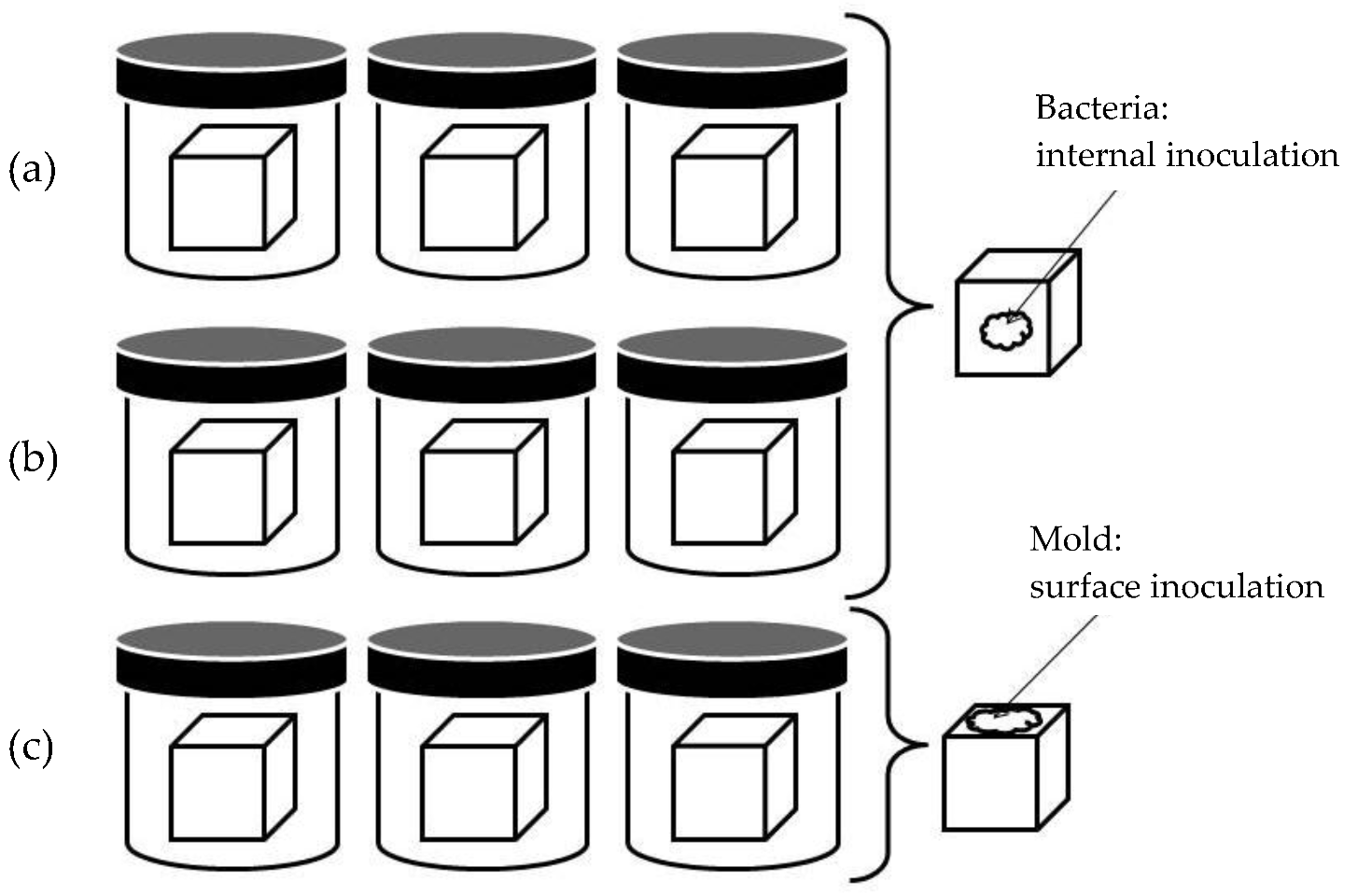
2.7. Antimicrobial Activity Test
2.8. Microorganism Inoculum Preparation
2.9. Statistical Analysis
3. Results and Discussion
3.1. Extraction and Composition Analysis of Essential Oils
3.2. Volatile Composition of Dairy Samples
3.3. Antimicrobial Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mato Rodriguez, L.; Alatossava, T. Effects of copper on germination, growth and sporulation of Clostridium tyrobutyricum. Food Microbiol. 2010, 27, 434–437. [Google Scholar] [CrossRef] [PubMed]
- Mullan, W.M.A. Causes and control of early gas production in cheddar cheese. Int. J. Dairy Technol. 2000, 53, 63–68. [Google Scholar] [CrossRef]
- Kure, C.F.; Skaar, I.; Brendehaug, J. Mould contamination in production of semi-hard cheese. Int. J. Food Microbiol. 2004, 93, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kure, C.F.; Wasteson, Y.; Brendehaug, J.; Skaar, I. Mould contaminants on Jarlsberg and Norvegia cheese blocks from four factories. Int. J. Food Microbiol. 2001, 70, 21–27. [Google Scholar] [CrossRef]
- Vázquez, B.I.; Fente, C.; Franco, C.M.; Vázquez, M.J.; Cepeda, A. Inhibitory effects of eugenol and thymol on Penicillium citrinum strains in culture media and cheese. Int. J. Food Microbiol. 2001, 67, 157–163. [Google Scholar] [CrossRef]
- Belitz, H.D.; Grosh, W.; Schieberle, P. Food Contamination. In Food Chemistry; Springer: Berlin, Germany, 2009. [Google Scholar]
- Bassi, D.; Puglisi, E.; Cocconcelli, P.S. Understanding the bacterial communities of hard cheese with blowing defect. Food Microbiol. 2015, 52, 106–118. [Google Scholar] [CrossRef]
- Klijn, N.; Nieuwenhof, F.F.; Hoolwerf, J.D.; van der Waals, C.B.; Weerkamp, A.H. Identification of Clostridium tyrobutyricum as the causative agent of late blowing in cheese by species-specific PCR amplification. Appl. Environ. Microbiol. 1995, 61, 2919–2924. [Google Scholar]
- Garde, S.; Arias, R.; Gaya, P.; Nuñez, M. Occurrence of Clostridium spp. in ovine milk and Manchego cheese with late blowing defect: Identification and characterization of isolates. Int. Dairy J. 2011, 21, 272–278. [Google Scholar] [CrossRef]
- Garde, S.; Ávila, M.; Arias, R.; Gaya, P.; Nuñez, M. Outgrowth inhibition of Clostridium beijerinckii spores by a bacteriocin-producing lactic culture in ovine milk cheese. Int. J. Food Microbiol. 2011, 150, 59–65. [Google Scholar] [CrossRef]
- Garde, S.; Gaya, P.; Arias, R.; Nuñez, M. Enhanced PFGE protocol to study the genomic diversity of Clostridium spp. isolated from Manchego cheeses with late blowing defect. Food Control 2012, 28, 392–399. [Google Scholar] [CrossRef]
- Gómez-Torres, N.; Garde, S.; Peirotén, Á.; Ávila, M. Impact of Clostridium spp. on cheese characteristics: Microbiology, color, formation of volatile compounds and off-flavors. Food Control 2015, 56, 186–194. [Google Scholar] [CrossRef]
- EFSA. EFSA Confirms Safe Levels for Nitrites and Nitrates Added to Food. Available online: http://www.efsa.europa.eu/en/press/news/170615 (accessed on 13 July 2019).
- D’Incecco, P.; Gatti, M.; Hogenboom, J.A.; Bottari, B.; Rosi, V.; Neviani, E.; Pellegrino, L. Lysozyme affects the microbial catabolism of free arginine in raw-milk hard cheeses. Food Microbiol. 2016, 57, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zantar, S.; Yedri, F.; Mrabet, R.; Laglaoui, A.; Bakkali, M.; Zerrouk, M.H. Effect of Thymus vulgaris and Origanum compactum essential oils on the shelf life of fresh goat cheese. J. Essent. Oil Res. 2014, 26, 76–84. [Google Scholar] [CrossRef]
- Carović-Stanko, K.; Orlić, S.; Politeo, O.; Strikić, F.; Kolak, I.; Milos, M.; Satovic, Z. Composition and antibacterial activities of essential oils of seven Ocimum taxa. Food Chem. 2010, 119, 196–201. [Google Scholar] [CrossRef]
- Hussain, A.I.; Anwar, F.; Hussain Sherazi, S.T.; Przybylski, R. Chemical composition, antioxidant and antimicrobial activities of basil (Ocimum basilicum) essential oils depends on seasonal variations. Food Chem. 2008, 108, 986–995. [Google Scholar] [CrossRef]
- Caleja, C.; Barros, L.; Antonio, A.L.; Ciric, A.; Barreira, J.C.M.; Sokovic, M.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Development of a functional dairy food: Exploring bioactive and preservation effects of chamomile (Matricaria recutita L.). J. Funct. Foods 2015, 16, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Govaris, A.; Botsoglou, E.; Sergelidis, D.; Chatzopoulou, P.S. Antibacterial activity of oregano and thyme essential oils against Listeria monocytogenes and Escherichia coli O157:H7 in feta cheese packaged under modified atmosphere. LWT Food Sci. Technol. 2011, 44, 1240–1244. [Google Scholar] [CrossRef]
- Dupas, C.; Métoyer, B.; Hatmi, H.E.; Adt, I.; Mahgoub, S.A.; Dumas, E. Plants: A natural solution to enhance raw milk cheese preservation? Food Res. Int. 2019, 108883. [Google Scholar] [CrossRef]
- Khorshidian, N.; Yousefi, M.; Khanniri, E.; Mortazavian, A.M. Potential application of essential oils as antimicrobial preservatives in cheese. Innov. Food Sci. Emerg. Tech. 2018, 45, 62–72. [Google Scholar] [CrossRef]
- Gutierrez, J.; Barry-Ryan, C.; Bourke, P. The antimicrobial efficacy of plant essential oil combinations and interactions with food ingredients. Int. J. Food Microbiol. 2008, 124, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Moreira, M.R.; Ponce, A.G.; Del Valle, C.E.; Roura, S.I. Effects of clove and tea tree oils on Escherichia coli O157:H7 in blanched spinach and minced cooked beef. J. Food Process. Preserv. 2007, 31, 379–391. [Google Scholar] [CrossRef]
- Smith-Palmer, A.; Stewart, J.; Fyfe, L. The potential application of plant essential oils as natural food preservatives in soft cheese. Food Microbiol. 2001, 18, 463–470. [Google Scholar] [CrossRef]
- Fernandes, R.V.D.B.; Guimarães, I.C.; Ferreira, C.L.R.; Botrel, D.A.; Borges, S.V.; de Souza, A.U. Microencapsulated Rosemary (Rosmarinus officinalis) Essential Oil as a Biopreservative in Minas Frescal Cheese. J. Food Process. Preserv. 2017, 41, e12759. [Google Scholar] [CrossRef]
- Hamedi, H.; Razavi-Rohani, S.M.; Gandomi, H. Combination Effect of Essential Oils of Some Herbs With Monolaurin on Growth and Survival of Listeria Monocytogenes in Culture Media and Cheese. J. Food Process. Preserv. 2014, 38, 304–310. [Google Scholar] [CrossRef]
- Moro, A.; Librán, C.M.; Berruga, M.I.; Zalacain, A.; Carmona, M. Mycotoxicogenic fungal inhibition by innovative cheese cover with aromatic plants. J. Sci. Food Agric. 2013, 93, 1112–1118. [Google Scholar] [CrossRef]
- Regulation (EC) No 1333/2008 of the European Parliament and of the Council of 16 December 2008 on Food Additives. Last Consolidated Version 02008R1333-EN-28.10.2019-041.001. Available online: http://data.europa.eu/eli/reg/2008/1333/2019-10-28 (accessed on December 2019).
- Licón, C.C.; Carmona, M.; Molina, A.; Berruga, M.I. Chemical, microbiological, textural, color, and sensory characteristics of pressed ewe milk cheeses with saffron (Crocus sativus L.) during ripening. J. Dairy Sci. 2012, 95, 4263–4274. [Google Scholar] [CrossRef] [Green Version]
- Licón, C.C.; Hurtado de Mendoza, J.; Maggi, L.; Berruga, M.I.; Martín Aranda, R.M.; Carmona, M. Optimization of headspace sorptive extraction for the analysis of volatiles in pressed ewes’ milk cheese. Int. Dairy J. 2012, 23, 53–61. [Google Scholar] [CrossRef]
- Moro, A.; Librán, C.M.; Berruga, M.I.; Carmona, M.; Zalacain, A. Dairy matrix effect on the transference of rosemary (Rosmarinus officinalis) essential oil compounds during cheese making. J. Sci. Food Agric. 2015, 95, 1507–1513. [Google Scholar] [CrossRef]
- Yang, W.-W.; Crow-Willard, E.N.; Ponce, A. Production and characterization of pure Clostridium spore suspensions. J. Appl. Microbiol. 2008, 106, 26–33. [Google Scholar] [CrossRef]
- Anastasiou, R.; Aktypis, A.; Georgalaki, M.; Papadelli, M.; De Vuyst, L.; Tsakalidou, E. Inhibition of Clostridium tyrobutyricum by Streptococcus macedonicus ACA-DC 198 under conditions mimicking Kasseri cheese production and ripening. Int. Dairy J. 2009, 19, 330–335. [Google Scholar] [CrossRef]
- Baratta, M.T.; Dorman, H.J.D.; Deans, S.G.; Figueiredo, A.C.; Barroso, J.G. Antimicrobial and antioxidant properties of some commercial essential oils. Flavour Fragr. J. 1998, 13, 235–244. [Google Scholar] [CrossRef]
- Cassel, E.; Vargas, R.M.F.; Martinez, N.; Lorenzo, D.; Dellacassa, E. Steam distillation modeling for essential oil extraction process. Ind. Crops Prod. 2009, 29, 171–176. [Google Scholar] [CrossRef]
- Singh Chouhan, K.B.; Tandey, R.; Sen, K.K.; Mehta, R.; Mandal, V. Critical analysis of microwave hydrodiffusion and gravity as a green tool for extraction of essential oils: Time to replace traditional distillation. Trends Food Sci. Technol. 2019, 92, 12–21. [Google Scholar] [CrossRef]
- De Carvalho, R.J.; de Souza, G.T.; Honório, V.G.; de Sousa, J.P.; da Conceição, M.L.; Maganani, M.; de Souza, E.L. Comparative inhibitory effects of Thymus vulgaris L. essential oil against Staphylococcus aureus, Listeria monocytogenes and mesophilic starter co-culture in cheese-mimicking models. Food Microbiol. 2015, 52, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Turhan, H. 23-Lemon balm. In Handbook of Herbs and Spices; Peter, K.V., Ed.; Woodhead Publishing: Cambridge, UK, 2006; pp. 390–399. [Google Scholar] [CrossRef]
- Carnat, A.P.; Carnat, A.; Fraisse, D.; Lamaison, J.L. The aromatic and polyphenolic composition of lemon balm (Melissa officinalis L. subsp. officinalis) tea. Pharm. Acta Hel. 1998, 72, 301–305. [Google Scholar] [CrossRef]
- Stahl-Biskup, E.; Venskutonis, R.P. 19-Thyme. In Handbook of Herbs and Spices; Peter, K.V., Ed.; Woodhead Publishing: Cambridge, UK, 2004; pp. 297–321. [Google Scholar] [CrossRef]
- Satyal, P.; Murray, B.L.; McFeetersand, R.L.; Setze, W.N. Essential Oil Characterization of Thymus vulgaris from Various Geographical Locations. Foods 2016, 5, 70. [Google Scholar] [CrossRef]
- Handa, S.S.; Khanuja, S.; Longo, G.; Rakesh, D.D. Extraction Technologies for Medicinal and Aromatic Plants; International Centre for Science and High Technology: Trieste, Italy, 2008; pp. 21–25. [Google Scholar]
- Gulluce, M.; Sahin, F.; Sokmen, M.; Ozer, H.; Daferera, D.; Sokmen, A.; Polissiou, M.; Adiguzel, A.; Ozkan, H. Antimicrobial and antioxidant properties of the essential oils and methanol extract from Mentha longifolia L. ssp. longifolia. Food Chem. 2007, 103, 1449–1456. [Google Scholar] [CrossRef]
- Okoh, O.O.; Sadimenko, A.P.; Afolayan, A.J. Comparative evaluation of the antibacterial activities of the essential oils of Rosmarinus officinalis L. obtained by hydrodistillation and solvent free microwave extraction methods. Food Chem. 2010, 120, 308–312. [Google Scholar] [CrossRef]
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Tornambé, G.; Cornu, A.; Verdier-Metz, I.; Pradel, P.; Kondjoyan, N.; Figueredo, G.; Hulin, S.; Martin, B. Addition of Pasture Plant Essential Oil in Milk: Influence on Chemical and Sensory Properties of Milk and Cheese. J. Dairy Sci. 2008, 91, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.S.; Frisvad, J.C.; Nielsen, P.V. Protection by fungal starters against growth and secondary metabolite production of fungal spoilers of cheese. Int. J. Food Microbiol. 1998, 42, 91–99. [Google Scholar] [CrossRef]
- Schlesser, J.E.; Gerdes, R.; Ravishankar, S.; Madsen, K.; Mowbray, J.; Teo, A.Y.-L. Survival of a Five-Strain Cocktail of Escherichia coli O157:H7 during the 60-Day Aging Period of Cheddar Cheese Made from Unpasteurized Milk. J. Food Protec. 2006, 69, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, A.; Hashemi, M.; Naghibi, S.S.; Mohammadi, S.; Khalili Sadaghiani, S. Properties of Bunium Persicum Essential Oil and its Application in Iranian White Cheese Against Listeria Monocytogenes and Escherichia Coli O157:H7. J. Food Safety 2016, 36, 563–570. [Google Scholar] [CrossRef]
- Le Bourhis, A.G.; Saunier, K.; Dore, J.; Carlier, J.P.; Chamba, J.F.; Popoff, M.R.; Tholozan, J.L. Development and validation of PCR primers to assess the diversity of Clostridium spp. in cheese by temporal temperature gradient gel electrophoresis. App. Environ. Microbiol. 2005, 71, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, P.F.; Cogan, T.M. Factors that Affect the Quality of Cheese. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T., Eds.; Elsevier Academic Press: London, UK, 2003; Volume 1, pp. 583–608. [Google Scholar]
- Walstra, P.; Wouters, J.T.; Geurts, T.J. Dairy Science and Technology; Taylor & Francis Group: London, UK, 2006. [Google Scholar]
- Sheehan, J. What causes the development of gas during ripening? In Cheese Problems Solved: The Microbiology of Cheese Ripening; PHL, M., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2007; pp. 131–133. [Google Scholar]
- Cogan, T.M. Cheese|Microbiology of cheese. In Encyclopedia of Dairy Sciences; John, W.F., Ed.; Academic Press: San Diego, CA, USA, 2011. [Google Scholar]
- Librán, C.M.; Moro, A.; Molina, A.; Carmona, M.; Berruga, M.I. Empleo potencial de plantas aromáticas para combatir defectos en quesos de oveja. In Parte I: Estudios In Vitro Frente a Penicillium Verrucosum. Proceedings of the XXXVII Congreso SEOC; Ciudad Real, Spain, 2012; pp. 169–173. [Google Scholar]
- Da Silva Dannenberg, G.; Funck, G.D.; Mattei, F.J.; da Silva, W.P.; Fiorentini, Â.M. Antimicrobial and antioxidant activity of essential oil from pink pepper tree (Schinus terebinthifolius Raddi) in vitro and in cheese experimentally contaminated with Listeria monocytogenes. Innov. Food Sci. Emerg. Technol. 2016, 36, 120–127. [Google Scholar] [CrossRef]
- Deans, S.G.; Ritchie, G. Antibacterial properties of plant essential oils. Int. J. Food Microbiol. 1987, 5, 165–180. [Google Scholar] [CrossRef]
- Librán, C.M.; Moro, A.; Carmona, M.; Molina, A.; Berruga, M.I. Empleo potencial de plantas aromáticas para combatir defectos en quesos de oveja. In Parte II: Estudios In Vitro Frente a Microorganismos Causantes de Hinchazones. Proceedings of the XXXVII Congreso SEOC; Ciudad Real, Spain, 2012; pp. 174–178. [Google Scholar]
- Razzaghi-Abyaneh, M.; Shams-Ghahfarokhi, M.; Rezaee, M.-B.; Jaimand, K.; Alinezhad, S.; Saberi, R.; Yoshinari, T. Chemical composition and antiaflatoxigenic activity of Carum carvi L., Thymus vulgaris and Citrus aurantifolia essential oils. Food Control 2009, 20, 1018–1024. [Google Scholar] [CrossRef]
- A, B.T.W. The Microbiology of cheese ripening. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Academic Press: Cambridge, UK, 2004. [Google Scholar]
- Goñi, P.; López, P.; Sánchez, C.; Gómez-Lus, R.; Becerril, R.; Nerín, C. Antimicrobial activity in the vapour phase of a combination of cinnamon and clove essential oils. Food Chem. 2009, 116, 982–989. [Google Scholar] [CrossRef]
- Reyes-Jurado, F.; Cervantes-Rincón, T.; Bach, H.; López-Malo, A.; Palou, E. Antimicrobial activity of Mexican oregano (Lippia berlandieri), thyme (Thymus vulgaris), and mustard (Brassica nigra) essential oils in gaseous phase. Ind. Crops Prod. 2019, 131, 90–95. [Google Scholar] [CrossRef]

 ; Lactic Acid Bacteria:
; Lactic Acid Bacteria:  ; Clostridium spp.:
; Clostridium spp.:  ).
; Lactic Acid Bacteria: ; Clostridium spp.: ).
).
; Lactic Acid Bacteria: ; Clostridium spp.: ). ; Lactic Acid Bacteria: ; Target microorganism: ). ***, *, NS: Significance level p < 0.001, p < 0.05 and non-significant, respectively. a, b, c: Different values among the same microbial group are significantly different between essential oils applications (p < 0.05).
; Lactic Acid Bacteria: ; Target microorganism: ). ***, *, NS: Significance level p < 0.001, p < 0.05 and non-significant, respectively. a, b, c: Different values among the same microbial group are significantly different between essential oils applications (p < 0.05).
; Lactic Acid Bacteria: ; Target microorganism: ). ***, *, NS: Significance level p < 0.001, p < 0.05 and non-significant, respectively. a, b, c: Different values among the same microbial group are significantly different between essential oils applications (p < 0.05).
; Lactic Acid Bacteria: ; Target microorganism: ). ***, *, NS: Significance level p < 0.001, p < 0.05 and non-significant, respectively. a, b, c: Different values among the same microbial group are significantly different between essential oils applications (p < 0.05).

{kind=link}
{kind=link}
{kind=link}
| Compounds | RT (min) | KI exp. * | m/z Pattern ** | Melissa officinalis | Ocimum basilicum | Thymus vulgaris |
|---|---|---|---|---|---|---|
| Number of compounds | 18 | 14 | 27 | |||
| Monoterpenes family | ||||||
| α-thujene | 28.34 | 924 | 77/93/136 | - | - | 0.43 |
| α-pinene | 28.86 | 937 | 93/136 | 0.51 | 0.27 | 4.74 |
| camphene | 29.56 | 946 | 93/121/136 | 2.71 | - | 1.69 |
| sabinene | 30.51 | 970 | 93/77/41 | 0.02 | 0.23 | 1.27 |
| β-pinene | 30.77 | 975 | 41/93/107/121 | 2.22 | 0.52 | 8.13 |
| myrcene | 30.83 | 988 | 41/93/69 | - | 0.31 | - |
| α-phelandrene | 31.74 | 1002 | 93/77/136 | - | - | 0.8 |
| α-terpinene | 32.22 | 1014 | 93/121/136 | - | - | 5.12 |
| β-ocimene (Z) | 32.55 | 1017 | 79/93/136 | 0.43 | - | - |
| p-cymene | 32.64 | 1020 | 91/119 | - | - | 6.96 |
| sylvestrene | 32.66 | 1024 | 41/68/93/136 | 0.48 | - | - |
| 1,8 cineole | 32.80 | 1026 | 43/108/139/154 | - | 5.85 | - |
| β-phelandrene | 32.90 | 1025 | 77/93/136 | - | - | 1.52 |
| β-ocimene (E) | 33.06 | 1032 | 79/93/136 | 0.31 | 0.84 | 0.11 |
| γ-terpinene | 33.74 | 1054 | 77/93/121/136 | - | - | 7.5 |
| 4-thujanol (Z) | 34.16 | 1065 | 43/71/93/139/154 | - | - | 1.44 |
| terpinolene | 34.83 | 1086 | 43/93/121/136 | 0.3 | - | 2.47 |
| linalool | 35.09 | 1089 | 43/71/154 | 1.71 | 11.21 | 34.54 |
| perillene | 35.14 | 1093 | 41/69/81/150 | 0.12 | - | - |
| 4-thujanol (E) | 35.37 | 1098 | 43/71/93/139/154 | - | - | 0.18 |
| citronellal | 37.04 | 1148 | 41/69/95/121/154 | 0.65 | - | - |
| camphor | 37.64 | 1141 | 41/95/152 | - | 0.54 | 0.3 |
| borneol | 38.01 | 1165 | 95/154 | - | - | 2.77 |
| terpinen-4-ol | 38.30 | 1174 | 43/71/154 | 0.28 | 0.33 | 10.54 |
| α-terpineol | 38.75 | 1186 | 43/59/93/136 | 0.56 | - | 2.14 |
| 4-allyl-anisole | 38.84 | 1189 | 77/121/148 | - | 58.21 | - |
| dihydro carvone (E) | 39.09 | 1194 | 41/67/95/152 | - | - | 0.51 |
| dihydro carvone (Z) | 39.41 | 1200 | 41/67/95/152 | - | - | 0.56 |
| linalyl acetate | 40.25 | 1210 | 43/93/121 | - | - | 1.55 |
| nerol | 40.38 | 1227 | 41/69/154 | 35.85 | - | - |
| carvone | 40.747 | 1235 | 54/82/93/150 | - | - | 0.13 |
| neral | 41.31 | 1239 | 41/69/109/152 | 35.34 | - | - |
| thymol | 41.43 | 1281 | 135/150/65 | - | - | 0.66 |
| bornyl acetate | 41.91 | 1288 | 41/95/121 | - | 1.18 | - |
| carvacrol | 41.98 | 1298 | 41/135/150 | - | - | 0.51 |
| eugenol | 44.05 | 1356 | 43/131/149/164 | - | 3.03 | - |
| neryl acetate | 44.06 | 1359 | 41/69/93/154 | 7.54 | - | - |
| methyl eugenol | 45.03 | 1403 | 41/107/163/178 | - | 1.28 | - |
| Sesquiterpenes | ||||||
| α-bergamotene (E)(Z) | 46.33 | 1432 | 41/93/119/204 | - | 3.2 | - |
| β-caryophyllene (E) | 46.48 | 1417 | 41/93/103/161 | 2.94 | - | 1.76 |
| Others | ||||||
| 1-octen-3-ol | 30.19 | 974 | 43/57 | - | - | 0.06 |
| Total area of all identified compounds (%) | 91.97 | 87 | 98.39 | |||
| Total Monoterpenes (%) | 89.03 | 83.8 | 96.57 | |||
| Total Sesquiterpenes (%) | 2.94 | 3.2 | 1.76 | |||
| Total Others (%) | - | - | 0.06 |
| Melissa officinalis | Ocimum basilicum | Thymus vulgaris | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Conc. (mg/kg) † | Transf. (%) ‡ | Conc. (mg/kg) | Transf. (%) | Conc. (mg/kg) | Transf. (%) | ||||||||||
| M§ | C | W | C | W | M | C | W | C | W | M | C | W | C | W | |
| Number of compounds | 11 | 9 | 10 | 19 | 18 | 19 | 22 | 22 | 18 | ||||||
| Monoterpene family | |||||||||||||||
| α-thujene | - | - | - | - | - | 0.04 | 0.03 | 0.01 | 75.00 | 25.00 | 2.19 | 0.28 | 0.22 | 12.79 | 10.05 |
| α-pinene | 1.53 | 0.18 | 0.23 | 11.76 | 15.03 | 0.47 | 0.33 | 0.09 | 70.21 | 19.15 | 21.10 | 2.58 | 2.19 | 12.23 | 10.38 |
| camphene | 8.60 | 0.99 | 1.36 | 11.51 | 15.81 | 0.13 | 0.08 | 0.03 | 61.54 | 23.08 | 7.79 | 0.92 | 0.77 | 11.81 | 9.88 |
| sabinene | - | - | - | - | - | 0.45 | 0.21 | 0.08 | 46.67 | 17.78 | 5.64 | 0.64 | 0.64 | 11.35 | 11.35 |
| β-pinene | 0.22 | 0.03 | 0.05 | 13.64 | 22.73 | 0.67 | 0.32 | 0.14 | 47.76 | 20.90 | 34.08 | 3.72 | 4.25 | 10.31 | 11.78 |
| α-phelandrene | - | - | - | - | - | - | - | - | - | - | 3.26 | 0.40 | 0.40 | 12.27 | 12.27 |
| α-terpinene | - | - | - | - | - | - | - | - | - | - | 22.52 | 2.50 | 2.80 | 11.10 | 12.43 |
| p-cymene | - | - | - | - | - | 0.27 | 0.12 | 0.05 | 44.44 | 18.52 | 13.04 | 1.48 | 1.65 | 11.35 | 12.65 |
| sylvestrene | - | - | - | - | - | - | - | - | - | - | 16.54 | 1.56 | - | 9.43 | - |
| 1.8 cineole | - | - | - | - | - | 15.49 | 4.55 | 4.37 | 29.37 | 28.21 | 5.73 | 0.61 | 2.70 | 10.65 | 47.12 |
| β-ocimene (E) | 0.48 | 0.06 | 0.09 | 12.50 | 18.75 | 1.28 | 0.47 | 0.26 | 36.72 | 20.31 | 0.45 | 0.06 | - | 13.33 | - |
| γ-terpinene | - | - | - | - | - | 0.10 | 0.04 | 0.03 | 40.00 | 30.00 | 27.88 | 3.10 | 3.95 | 11.12 | 14.17 |
| 4-thujanol (Z) | - | - | - | - | - | - | - | - | - | - | 0.52 | 0.03 | - | 5.77 | - |
| terpinolene | 0.45 | 0.04 | 0.06 | 8.89 | 13.33 | 0.26 | 0.34 | 0.06 | 130.77 | 23.08 | 7.19 | 0.88 | 1.17 | 12.24 | 16.27 |
| linalool | 1.19 | 0.17 | 0.36 | 14.29 | 30.25 | 13.99 | 3.85 | 3.62 | 27.52 | 25.88 | 61.83 | 6.57 | 12.98 | 10.63 | 20.99 |
| 4-thujanol (E) | - | - | - | - | - | - | - | - | - | - | 1.99 | 0.13 | - | 6.53 | - |
| camphor | - | - | - | - | - | 0.61 | 0.18 | 0.15 | 29.51 | 24.59 | 0.85 | 0.09 | 0.17 | 10.59 | 20.00 |
| borneol | - | - | - | - | - | 0.15 | 0.04 | 0.03 | 26.67 | 20.00 | 2.14 | 0.30 | 0.49 | 14.02 | 22.90 |
| terpinen-4-ol | 0.15 | - | 0.02 | - | 13.33 | 0.20 | 0.05 | 0.04 | 25.00 | 20.00 | 13.03 | 1.45 | 3.31 | 11.13 | 25.40 |
| α-terpineol | - | - | - | - | - | - | - | - | - | - | 1.26 | 0.18 | 0.40 | 14.29 | 31.75 |
| 4-allyl-anisole | - | - | - | - | - | 47.02 | 12.70 | 9.97 | 27.01 | 21.20 | - | - | - | - | - |
| linalyl acetate | - | - | - | - | - | - | - | - | - | - | 0.61 | 0.08 | - | 13.11 | - |
| nerol | 17.56 | 0.86 | 3.57 | 4.90 | 20.33 | - | - | - | - | - | - | - | - | - | - |
| neral | 16.30 | 0.86 | 3.38 | 5.28 | 20.74 | - | - | - | - | - | - | - | - | - | - |
| eugenol | - | - | - | - | - | 0.77 | - | 0.07 | - | 9.09 | - | - | - | - | - |
| bornyl acetate | - | - | - | - | - | 0.64 | 0.18 | 0.12 | 28.13 | 18.75 | 0.17 | 0.02 | 0.08 | 11.76 | 47.06 |
| neryl acetate | 1.28 | 0.18 | 0.34 | 14.06 | 26.56 | - | - | - | - | - | - | - | - | - | - |
| methyl eugenol | - | - | - | - | - | 0.17 | 0.03 | 0.03 | 17.65 | 17.65 | - | - | - | - | - |
| Sesquiterpene family | |||||||||||||||
| α-bergamotene (E)(Z) | - | - | - | - | - | 0.68 | 0.18 | 0.14 | 26.47 | 20.59 | - | - | - | - | - |
| β-caryophyllene (E) | 0.49 | 0.06 | 0.15 | 12.24 | 30.61 | - | - | - | - | - | 0.48 | 0.08 | 0.19 | 16.67 | 39.58 |
| Total identified compounds | 48.25 | 3.33 | 9.61 | 7.11 | 19.92 | 83.39 | 23.70 | 19.29 | 28.42 | 23.13 | 250.29 | 27.66 | 38.36 | 11.05 | 15.33 |
| Total monoterpenes | 47.76 | 3.37 | 9.46 | 7.06 | 19.81 | 82.71 | 23.52 | 19.15 | 28.44 | 23.15 | 249.81 | 27.58 | 38.17 | 11.04 | 15.28 |
| Total sesquiterpenes | 0.49 | 0.06 | 0.15 | 12.24 | 30.61 | 0.68 | 0.18 | 0.14 | 26.47 | 20.59 | 0.48 | 0.08 | 0.19 | 16.67 | 39.58 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Licon, C.C.; Moro, A.; Librán, C.M.; Molina, A.M.; Zalacain, A.; Berruga, M.I.; Carmona, M. Volatile Transference and Antimicrobial Activity of Cheeses Made with Ewes’ Milk Fortified with Essential Oils. Foods 2020, 9, 35. https://doi.org/10.3390/foods9010035
Licon CC, Moro A, Librán CM, Molina AM, Zalacain A, Berruga MI, Carmona M. Volatile Transference and Antimicrobial Activity of Cheeses Made with Ewes’ Milk Fortified with Essential Oils. Foods. 2020; 9(1):35. https://doi.org/10.3390/foods9010035
Chicago/Turabian StyleLicon, Carmen C., Armando Moro, Celia M. Librán, Ana M. Molina, Amaya Zalacain, M. Isabel Berruga, and Manuel Carmona. 2020. "Volatile Transference and Antimicrobial Activity of Cheeses Made with Ewes’ Milk Fortified with Essential Oils" Foods 9, no. 1: 35. https://doi.org/10.3390/foods9010035
APA StyleLicon, C. C., Moro, A., Librán, C. M., Molina, A. M., Zalacain, A., Berruga, M. I., & Carmona, M. (2020). Volatile Transference and Antimicrobial Activity of Cheeses Made with Ewes’ Milk Fortified with Essential Oils. Foods, 9(1), 35. https://doi.org/10.3390/foods9010035