Physicochemical Effects of Lactobacillus plantarum and Lactobacillus casei Cocultures on Soy–Wheat Flour Dough Fermentation




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Dough Preparation
2.3. Model Media and Dough Fermentation
2.4. Testing Methods
2.4.1. pH
2.4.2. Cell Viability
2.4.3. Rheological Properties
2.4.4. HPLC
2.4.5. Statistical Analysis
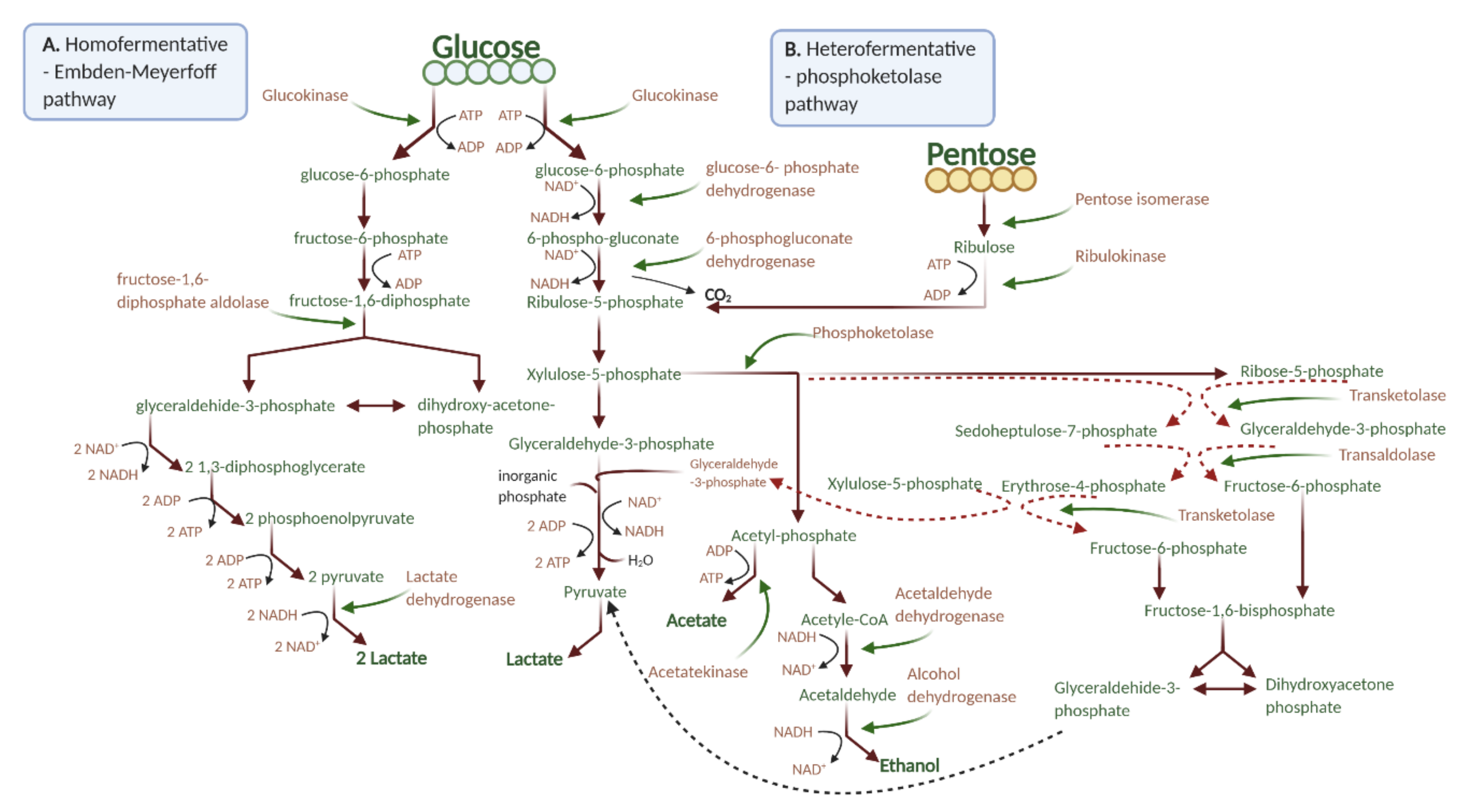
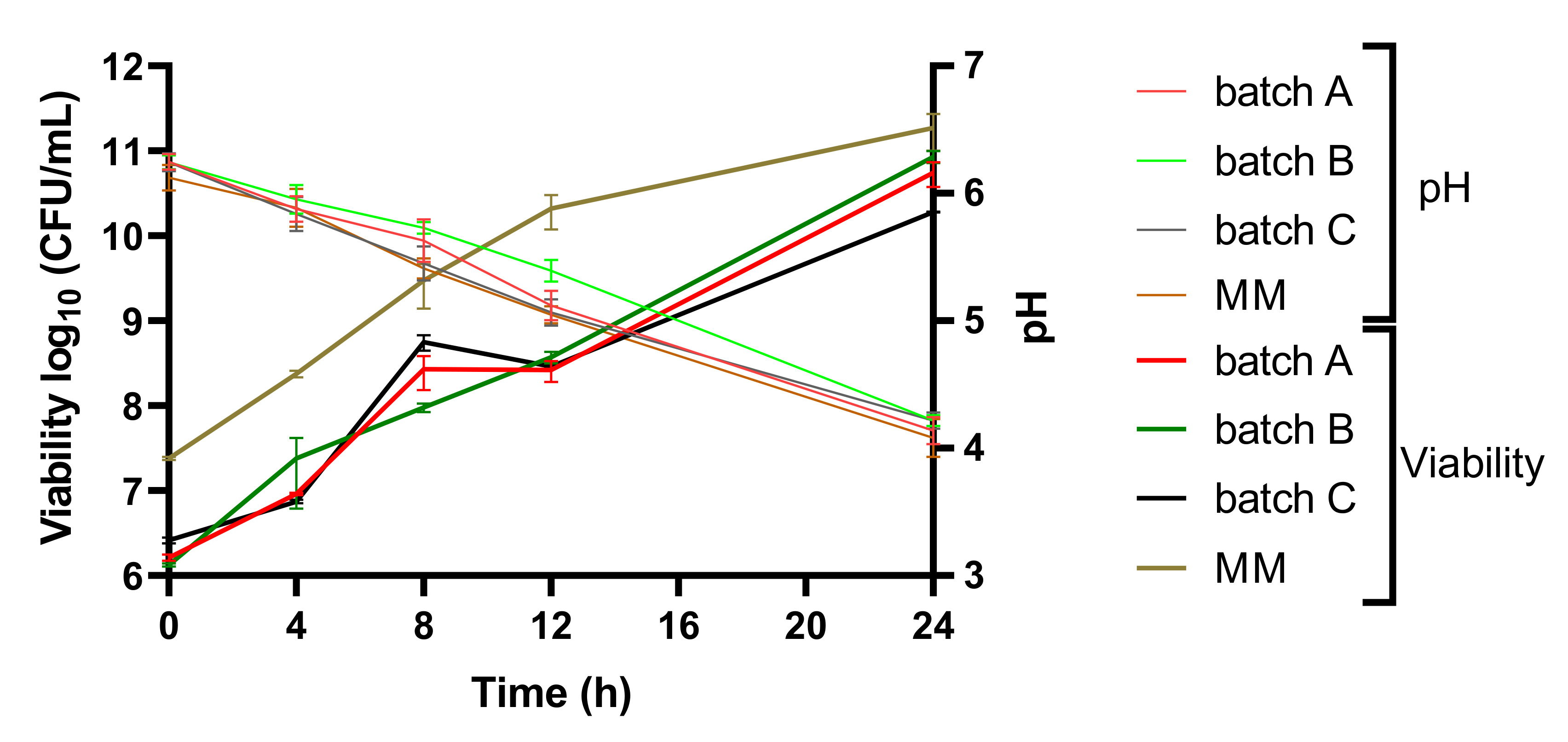
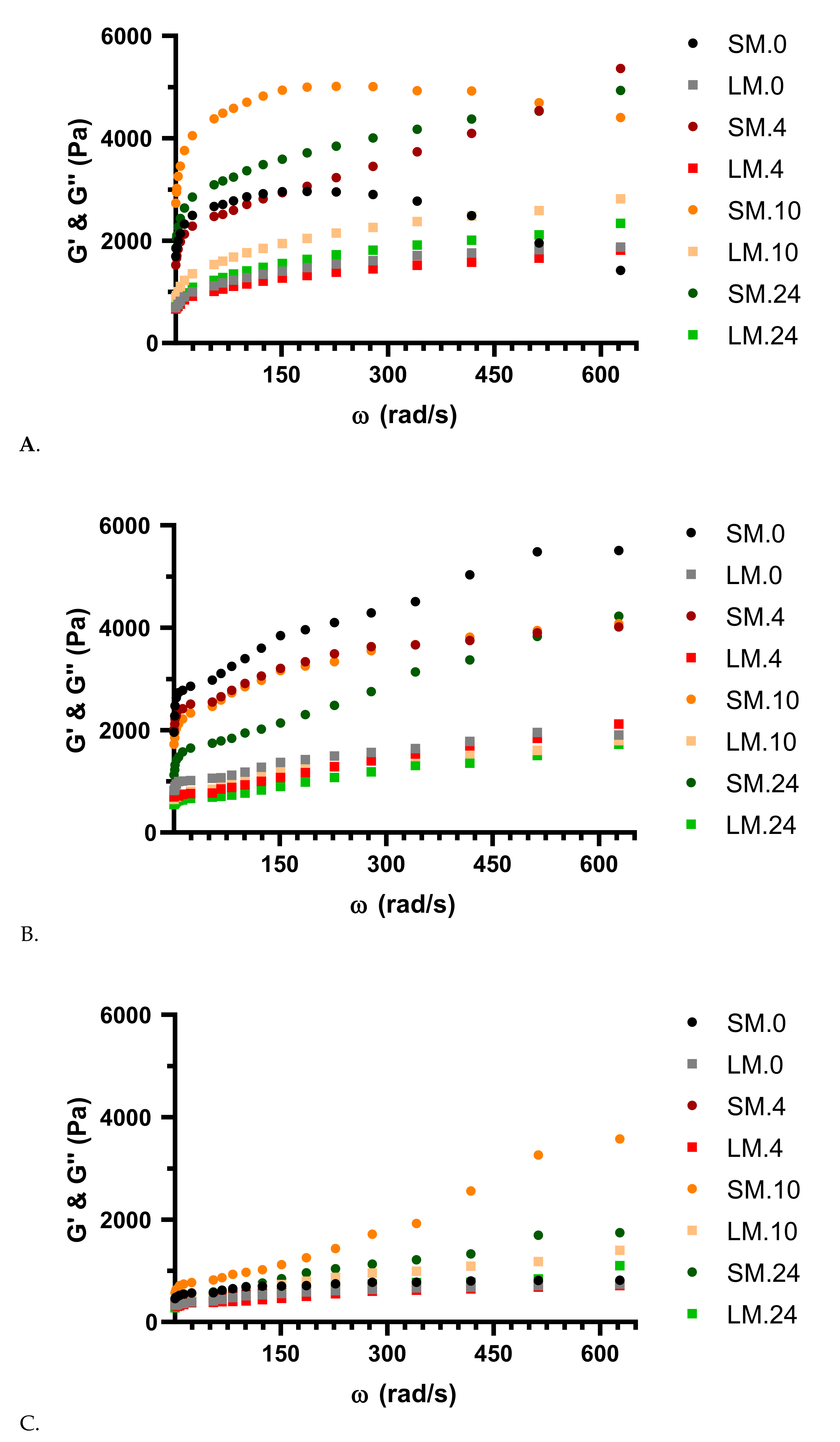
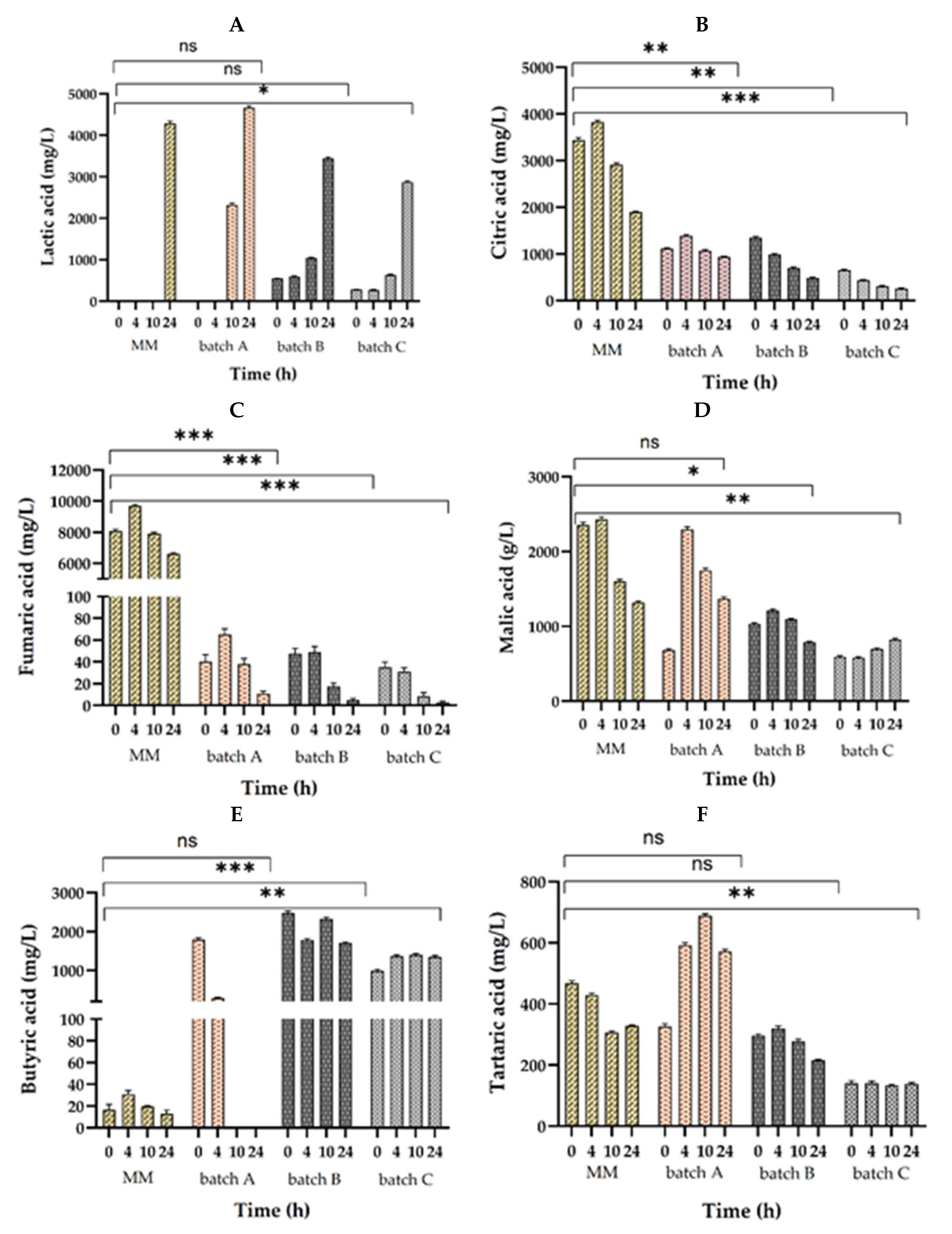
3. Results and Discussion
3.1. LAB Viability and pH
3.2. Viscoelastic Behavior of Dough
3.3. Organic Acid Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saget, S.; Costa, M.; Barilli, E.; Wilton, M.; Vasconcelos, D.; Sancho, C.; Styles, D.; Williams, M. Substituting wheat with chickpea flour in pasta production delivers more nutrition at a lower environmental cost. Sustain. Prod. Consum. 2020, 24, 26–38. [Google Scholar] [CrossRef]
- Chis, M.S.; Paucean, A.; Man, S.M.; Bonta, V.; Pop, A.; Stan, L.; Beldean, B.V.; Pop, C.R.; Mureșan, V.; Muste, S.; et al. Effect of rice flour fermentation with Lactobacillus spicheri DSM 15429 on the nutritional features of gluten-free muffins. Foods 2020, 9, 822. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole grains and phenolic acids: A review on bioactivity, functionality, health benefits and bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chatzidimitriou, E.; Wood, L.; Hasanalieva, G.; Markelou, E.; Iversen, P.O.; Seal, C.; Baranski, M.; Vigar, V.; Ernst, L.; et al. Effect of wheat species (Triticum aestivum vs. T. spelta), farming system (organic vs. conventional) and flour type (wholegrain vs. white) on composition of wheat flour—Results of a retail survey in the UK and Germany—2. Antioxidant activity, and phenoli. Food Chem. X 2020, 6, 100091. [Google Scholar] [CrossRef] [PubMed]
- Guardianelli, L.M.; Salinas, M.V.; Puppo, M.C. Hydration and rheological properties of amaranth-wheat flour dough: Influence of germination of amaranth seeds. Food Hydrocoll. 2019, 97, 105242. [Google Scholar] [CrossRef]
- Yamul, D.K.; Navarro, A.S. Effect of hydrocolloids on structural and functional properties of wheat/potato (50/50) flour dough. Food Struct. 2020, 24, 100138. [Google Scholar] [CrossRef]
- Xu, M.; Hou, G.G.; Ma, F.; Ding, J.; Deng, L.; Kahraman, O.; Niu, M.; Trivettea, K.; Lee, B.; Wu, L.; et al. Evaluation of aleurone flour on dough, textural, and nutritional properties of instant fried noodles. LWT 2020, 126, 109294. [Google Scholar] [CrossRef]
- Paucean, A.; Man, S.M.; Chis, M.S.; Mureşan, V.; Pop, C.R.; Socaci, S.A.; Muresan, C.C.; Muste, S. Use of pseudocereals preferment made with aromatic yeast strains for enhancing wheat bread quality. Foods 2019, 8, 443. [Google Scholar] [CrossRef] [Green Version]
- Jayachandran, M.; Xu, B. An insight into the health benefits of fermented soy products. Food Chem. 2019, 271, 362–371. [Google Scholar] [CrossRef]
- Hu, C.; Wong, W.-T.; Wu, R.; Lai, W.-F. Biochemistry and use of soybean isoflavones in functional food development. Crit. Rev. Food Sci. Nutr. 2019, 1–15. [Google Scholar] [CrossRef]
- Simmons, A.L.; Smith, K.B.; Vodovotz, Y. Soy ingredients stabilize bread dough during frozen storage. J. Cereal Sci. 2015, 56, 232–238. [Google Scholar] [CrossRef]
- Bojňanská, T.; Šmitalová, J.; Vollmannová, A.; Tokár, M.; Vietoris, V. Bakery products with the addition of soybean flour and their quality after freezer storage of dough. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 18–22. [Google Scholar] [CrossRef]
- Angioloni, A.; Romani, S.; Pinnavaia, G.G.; Rosa, M.D. Characteristics of bread making doughs: Influence of sourdough fermentation on the fundamental rheological properties. Eur. Food Res. Technol. 2006, 222, 54–57. [Google Scholar] [CrossRef]
- Zhang, Y.; Hong, T.; Yu, W.; Yang, N.; Jin, Z.; Xu, X. Structural, thermal and rheological properties of gluten dough: Comparative changes by dextran, weak acidification and their combination. Food Chem. 2020, 330, 127154. [Google Scholar] [CrossRef] [PubMed]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Vodnar, D.C.; Teleky, B.E.; Pop, C.R.; Stan, L.; Borsai, O.; Kadar, C.B.; Urcan, A.C.; et al. Quinoa sourdough fermented with Lactobacillus plantarum ATCC 8014 designed for gluten-free muffins—A powerful tool to enhance bioactive compounds. Appl. Sci. 2020, 10, 7140. [Google Scholar] [CrossRef]
- Masiá, C.; Jensen, P.E.; Buldo, P. Effect of Lactobacillus rhamnosus on physicochemical properties of fermented plant-based raw materials. Foods 2020, 9, 1182. [Google Scholar] [CrossRef] [PubMed]
- Shirotani, N.; Bygvraa, A.; Lametsch, R.; Agerlin, M.; Rattray, F.P.; Ipsen, R. Proteolytic activity of selected commercial Lactobacillus helveticus strains on soy protein isolates. Food Chem. 2020, 340, 128152. [Google Scholar] [CrossRef]
- Brizuela, N.; Tymczyszyn, E.E.; Semorile, L.C.; Valdes La Hens, D.; Delfederico, L.; Hollmann, A.; Bravo-Ferrada, B. Lactobacillus plantarum as a malolactic starter culture in winemaking: A new (old) player? Electron. J. Biotechnol. 2019, 38, 10–18. [Google Scholar] [CrossRef]
- Collado-Fernández, M. Bread and dough fermentation. Encycl. Food Sci. Nutr. 2003, 1997, 647–655. [Google Scholar] [CrossRef]
- Endo, A.; Dicks, L.M.T. Physiology of the LAB. In Lactic Acid Bacteria: Biodiversity and Taxonomy; Wiley Blackwell: Hoboken, NJ, USA, 2014; pp. 13–30. [Google Scholar]
- Paucean, A.; Vodnar, D.C.; Socaci, S.A.; Socaciu, C. Carbohydrate metabolic conversions to lactic acid and volatile derivatives, as influenced by Lactobacillus plantarum ATCC 8014 and Lactobacillus casei ATCC 393 efficiency during in vitro and sourdough fermentation. Eur. Food Res. Technol. 2013, 237, 679–689. [Google Scholar] [CrossRef]
- Saxami, G.; Ypsilantis, P.; Sidira, M.; Simopoulos, C.; Kourkoutas, Y.; Galanis, A. Distinct adhesion of probiotic strain Lactobacillus casei ATCC 393 to rat intestinal mucosa. Anaerobe 2012, 18, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Pop, O.L.; Dulf, F.V.; Cuibus, L.; Castro-Giráldez, M.; Fito, P.J.; Vodnar, D.C.; Coman, C.; Socaciu, C.; Suharoschi, R. Characterization of a sea buckthorn extract and its effect on free and encapsulated Lactobacillus casei. Int. J. Mol. Sci. 2017, 18, 2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Martău, G.A.; Coman, V.; Vodnar, D.C. Recent advances in the biotechnological production of erythritol and mannitol. Crit. Rev. Biotechnol. 2020, 40, 608–622. [Google Scholar] [CrossRef]
- Voinea, A.; Stroe, S.-G.; Codină, G.G. The effect of sodium reduction by sea salt and dry sourdough addition on the wheat flour dough rheological properties. Foods 2020, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Yildirim-Mavis, C.; Yilmaz, M.T.; Dertli, E.; Arici, M.; Ozmen, D. Non-linear rheological (LAOS) behavior of sourdough-based dough. Food Hydrocoll. 2019, 96, 481–492. [Google Scholar] [CrossRef]
- Autio, K.; Flander, L.; Kinnunen, A.; Heinonen, R. Bread quality relationship with rheological measurements of wheat flour dough. Cereal Chem. 2001, 78, 654–657. [Google Scholar] [CrossRef]
- Oh, I.; Lee, S. Rheological, microstructural, and tomographical studies on the rehydration improvement of hot air-dried noodles with oleogel. J. Food Eng. 2020, 268. [Google Scholar] [CrossRef]
- Marco, C.; Rosell, C.M. Functional and rheological properties of protein enriched gluten free composite flours. J. Food Eng. 2008, 88, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Sozer, N. Rheological properties of rice pasta dough supplemented with proteins and gums. Food Hydrocoll. 2009, 23, 849–855. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, J.; Tang, X. Effects of whey and soy protein addition on bread rheological property of wheat flour. J. Texture Stud. 2018, 49, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Psimouli, V.; Oreopoulou, V. The effect of alternative sweeteners on batter rheology and cake properties. J. Sci. Food Agric. 2012, 92, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Asghar, A.; Anjum, F.M.; Butt, M.S.; Randhawa, M.A.; Akhtar, S. Effect of polyols on the rheological and sensory parameters of frozen dough pizza. Food Sci. Technol. Res. 2012, 18, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Omedi, J.O.; Huang, W.; Su, X.; Liu, R.; Tang, X.; Xu, Y.; Rayas-Duarte, P. Effect of five lactic acid bacteria starter type on angiotensin-I converting enzyme inhibitory activity and emulsifying properties of soy flour sourdoughs with and without wheat bran supplementation. J. Cereal Sci. 2016, 69, 57–63. [Google Scholar] [CrossRef]
- Ammar, A.S.; Salem, S.A.; Badr, F.H. Rheological properties of wheat flour dough as affected by addition of whey and soy proteins. Pakistan J. Nutr. 2011, 10, 302–306. [Google Scholar] [CrossRef]
- Crockett, R.; Ie, P.; Vodovotz, Y. Effects of soy protein isolate and egg white solids on the physicochemical properties of gluten-free bread. Food Chem. 2015, 129, 84–91. [Google Scholar] [CrossRef]
- Yang, S.; Jeong, S.; Lee, S. Elucidation of rheological properties and baking performance of frozen doughs under different thawing conditions. J. Food Eng. 2020, 284, 110084. [Google Scholar] [CrossRef]
- Teleky, B.E.; Martău, A.G.; Ranga, F.; Chețan, F.; Vodnar, D.C.; Gheorghe, A.; Chet, F. Exploitation of lactic acid bacteria and baker’s yeast as single or multiple starter cultures of wheat flour dough enriched with soy flour. Biomolecules 2020, 10, 778. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Rizzello, C.G.; Alfonsi, G.; Arnault, P.; Cappelle, S.; Cagno, R.D.; Gobbetti, M. Use of sourdough lactobacilli and oat fibre to decrease the glycaemic index of white wheat bread. Br. J. Nutr. 2007, 98, 1196–1205. [Google Scholar] [CrossRef] [Green Version]
- Bottani, M.; Brasca, M.; Ferraretto, A.; Cardone, G.; Casiraghi, M.C.; Lombardi, G.; De Noni, I.; Cattaneo, S.; Silvetti, T. Chemical and nutritional properties of white bread leavened by lactic acid bacteria. J. Funct. Foods 2018, 45, 330–338. [Google Scholar] [CrossRef]
- Mitrea, L.; Trif, M.; Vodnar, D.-C. The effect of crude glycerol impurities on 1,3-propanediol biosynthesis by Klebsiella pneumoniae DSMZ 2026. Renew. Energy 2020, 153, 1418–1427. [Google Scholar] [CrossRef]
- Călinoiu, L.-F.; Catoi, A.-F.; Vodnar, D.C. Solid-state yeast fermented wheat and oat bran as a route for delivery of antioxidants. Antioxidants 2019, 8, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, K.; Teleky, B.E.; Mitrea, L.; Călinoiu, L.F.; Martău, G.A.; Simon, E.; Varvara, R.A.; Vodnar, D.C. Active packaging-poly (vinyl alcohol) films enriched with tomato by-products extract. Coatings 2020, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Mitrea, L.; Călinoiu, L.-F.F.; Martău, G.-A.; Szabo, K.; Teleky, B.-E.E.; Mureșan, V.; Rusu, A.-V.V.; Socol, C.-T.T.; Vodnar, D.-C.C.; Mărtau, G.A.; et al. Poly (vinyl alcohol)-based biofilms plasticized with polyols and colored with pigments extracted from tomato by-products. Polymers 2020, 12, 532. [Google Scholar] [CrossRef] [Green Version]
- Mitrea, L.; Vodnar, D.C. Klebsiella pneumoniae—A useful pathogenic strain for biotechnological purposes: Diols biosynthesis under controlled and uncontrolled pH levels. Pathogens 2019, 8, 293. [Google Scholar] [CrossRef] [Green Version]
- Mitrea, L.; Leopold, L.F.; Bouari, C.; Vodnar, D.C. Separation and Purification of Biogenic 1,3-Propanediol from Fermented Glycerol through Flocculation and Strong Acidic Ion-Exchange Resin. Biomolecules 2020, 10, 1601. [Google Scholar] [CrossRef]
- Szabo, K.; Diaconeasa, Z.; Catoi, A.; Vodnar, D.C. Screening of Ten Tomato Varieties Processing Waste for Bioactive Components and Their Related Antioxidant and Antimicrobial Activities. Antioxidants 2019, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Yang, Z.; Huang, W.; Omedi, J.O.; Wang, F.; Zou, Q.; Zheng, J. Isoflavone aglycones enrichment in soybean sourdough bread fermented by lactic acid bacteria strains isolated from traditional Qu starters: Effects on in-vitro gastrointestinal digestion, nutritional, and baking properties. Cereal Chem. 2018, 96, 129–141. [Google Scholar] [CrossRef]
- Aguirre, L.; Hebert, E.M.; Garro, M.S.; Savoy de Giori, G. Proteolytic activity of Lactobacillus strains on soybean proteins. LWT Food Sci. Technol. 2014, 59, 780–785. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Gholamhosseinpour, A.; Mousavi Khaneghah, A. Fermentation of acorn dough by lactobacilli strains: Phytic acid degradation and antioxidant activity. LWT 2019, 100, 144–149. [Google Scholar] [CrossRef]
- Gerez, C.L.; Dallagnol, A.; Rollán, G.; Font de Valdez, G. A combination of two lactic acid bacteria improves the hydrolysis of gliadin during wheat dough fermentation. Food Microbiol. 2012, 32, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Gaglio, R.; Alfonzo, A.; Barbera, M.; Franciosi, E.; Francesca, N.; Moschetti, G.; Settanni, L. Persistence of a mixed lactic acid bacterial starter culture during lysine fortification of sourdough breads by addition of pistachio powder. Food Microbiol. 2020, 86, 103349. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Campanella, O. Impulse viscoelastic characterization of wheat flour dough during fermentation. J. Food Eng. 2013, 118, 266–270. [Google Scholar] [CrossRef]
- Edwards, N.M.; Peressini, D.; Dexter, J.E.; Mulvaney, S.J. Viscoelastic properties of durum wheat and common wheat dough of different strengths. Rheol. Acta 2001, 40, 142–153. [Google Scholar] [CrossRef]
- Mariotti, M.; Lucisano, M.; Ambrogina Pagani, M.; Perry, K.W.N. The role of corn starch, amaranth flour, pea isolate, and psyllium flour on the rheological properties and the ultrastructure of gluten-free doughs. Food Res. Int. 2009, 42, 963–975. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef]
- Hansen, A.; Schieberle, P. Generation of aroma compounds during sourdough fermentation: Applied and fundamental aspects. Trends Food Sci. Technol. 2005, 16, 85–94. [Google Scholar] [CrossRef]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Sterr, Y.; Weiss, A.; Schmidt, H. Evaluation of lactic acid bacteria for sourdough fermentation of amaranth. Int. J. Food Microbiol. 2009, 136, 75–82. [Google Scholar] [CrossRef]
- Debonne, E.; Van Schoors, F.; Maene, P.; Van Bockstaele, F.; Vermeir, P.; Verwaeren, J.; Eeckhout, M.; Devlieghere, F. Comparison of the antifungal effect of undissociated lactic and acetic acid in sourdough bread and in chemically acidified wheat bread. Int. J. Food Microbiol. 2020, 321, 1–19. [Google Scholar] [CrossRef]
- Robert, H.; Gabriel, V.; Lefebvre, D.; Rabier, P.; Vayssier, Y.; Fontagné-Faucher, C. Study of the behaviour of Lactobacillus plantarum and Leuconostoc starters during a complete wheat sourdough breadmaking process. LWT Food Sci. Technol. 2006, 39, 256–265. [Google Scholar] [CrossRef]
- Kunkee, R.E. Some roles of malic acid in the malolactic fermentation in wine making. FEMS Microbiol. Rev. 1991, 88, 55–72. [Google Scholar]
- Filipcev, B.; Simurina, O.; Bodroza-Solarov, M. Combined effect of xylanase, ascorbic and citric acid in regulating the quality of bread made from organically grown spelt cultivars. J. Food Qual. 2014, 1–11. [Google Scholar] [CrossRef]
- Su, X.; Wu, F.; Zhang, Y.; Yang, N.; Chen, F.; Jin, Z.; Xu, X. Effect of organic acids on bread quality improvement. Food Chem. 2019, 278, 267–275. [Google Scholar] [CrossRef]
- Huda-Faujan, N.; Abdulamir, A.S.; Fatimah, A.B.; Muhammad Anas, O.; Shuhaimi, M.; Yazid, A.M.; Loong, Y.Y. The impact of the level of the intestinal short chain fatty acids in inflammatory bowel disease patients versus healthy subjects. Open Biochem. J. 2010, 4, 53–58. [Google Scholar] [CrossRef]
- Stevens, M.J.A.; Wiersma, A.; de Vos, W.M.; Kuipers, O.P.; Smid, E.J. Improvement of Lactobacillus plantarum Aerobic Growth as Directed by Comprehensive Transcriptome Analysis. Appl. Environ. Microbiol. 2008, 74, 4776–4778. [Google Scholar] [CrossRef] [Green Version]
- Sieuwerts, S.; Bron, P.A.; Smid, E.J. Mutually stimulating interactions between lactic acid bacteria and Saccharomyces cerevisiae in sourdough fermentation. LWT Food Sci. Technol. 2018, 90, 201–206. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teleky, B.-E.; Martău, G.A.; Vodnar, D.C. Physicochemical Effects of Lactobacillus plantarum and Lactobacillus casei Cocultures on Soy–Wheat Flour Dough Fermentation. Foods 2020, 9, 1894. https://doi.org/10.3390/foods9121894
Teleky B-E, Martău GA, Vodnar DC. Physicochemical Effects of Lactobacillus plantarum and Lactobacillus casei Cocultures on Soy–Wheat Flour Dough Fermentation. Foods. 2020; 9(12):1894. https://doi.org/10.3390/foods9121894
Chicago/Turabian StyleTeleky, Bernadette-Emőke, Gheorghe Adrian Martău, and Dan Cristian Vodnar. 2020. "Physicochemical Effects of Lactobacillus plantarum and Lactobacillus casei Cocultures on Soy–Wheat Flour Dough Fermentation" Foods 9, no. 12: 1894. https://doi.org/10.3390/foods9121894
APA StyleTeleky, B. -E., Martău, G. A., & Vodnar, D. C. (2020). Physicochemical Effects of Lactobacillus plantarum and Lactobacillus casei Cocultures on Soy–Wheat Flour Dough Fermentation. Foods, 9(12), 1894. https://doi.org/10.3390/foods9121894