
Dioxin-like Activity in Pregnant Women and Indices of Fetal Growth: The ACCEPT Birth Cohort


Abstract
:1. Introduction
2. Materials and Methods
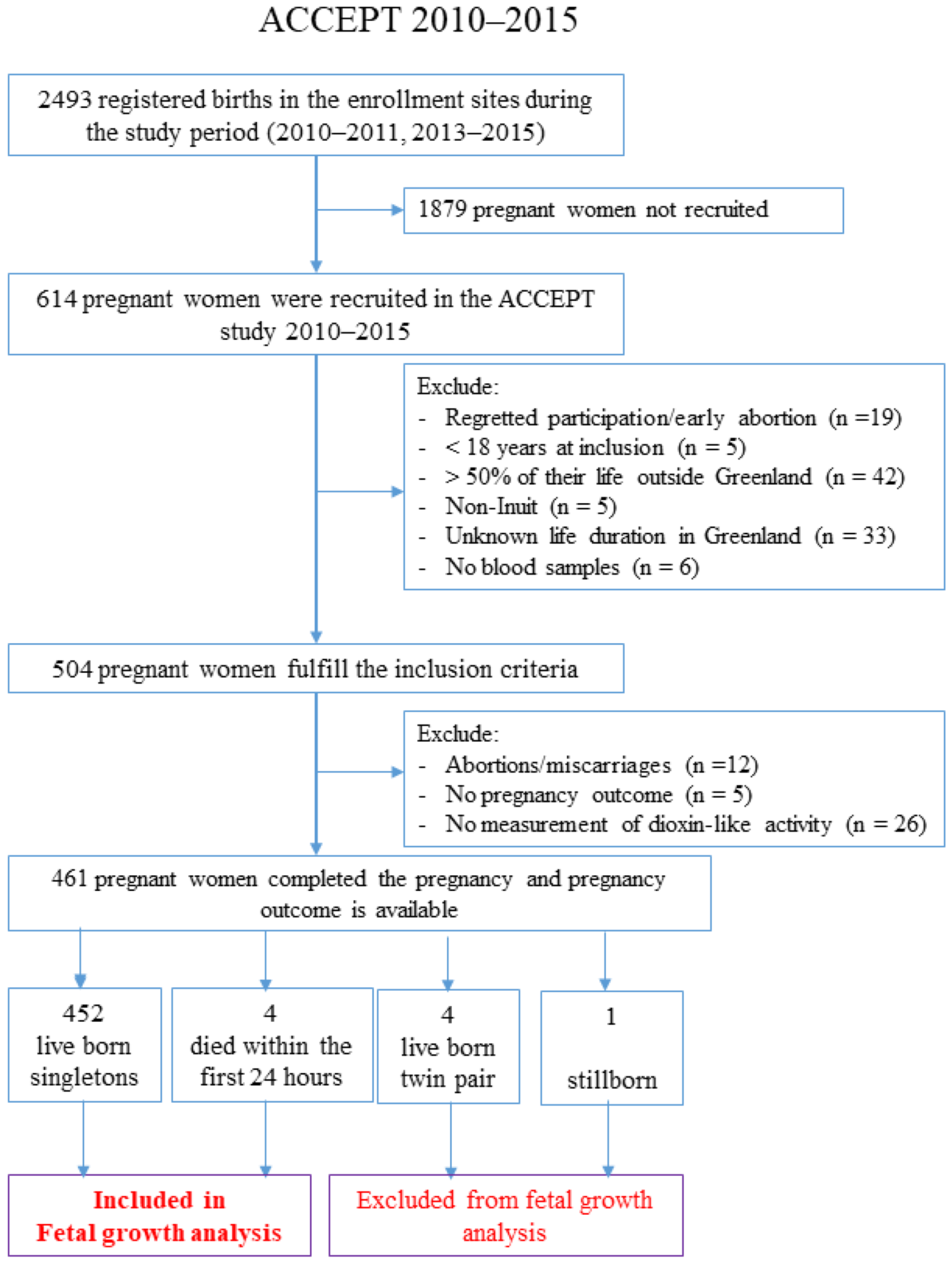
2.1. Study Population
2.2. Plasma Fatty Acids Easurement
2.3. Plasma Cotinine Levels Measurement
2.4. Measurement of Lipophilic POPs and Metals
2.5. Serum Sample Extractions for Determination of AhR Mediated Dioxin-like Activity
2.6. Measurement of Dioxin-like Activity Using AhR Transactivation Assay
2.7. Fetal Growth Indices
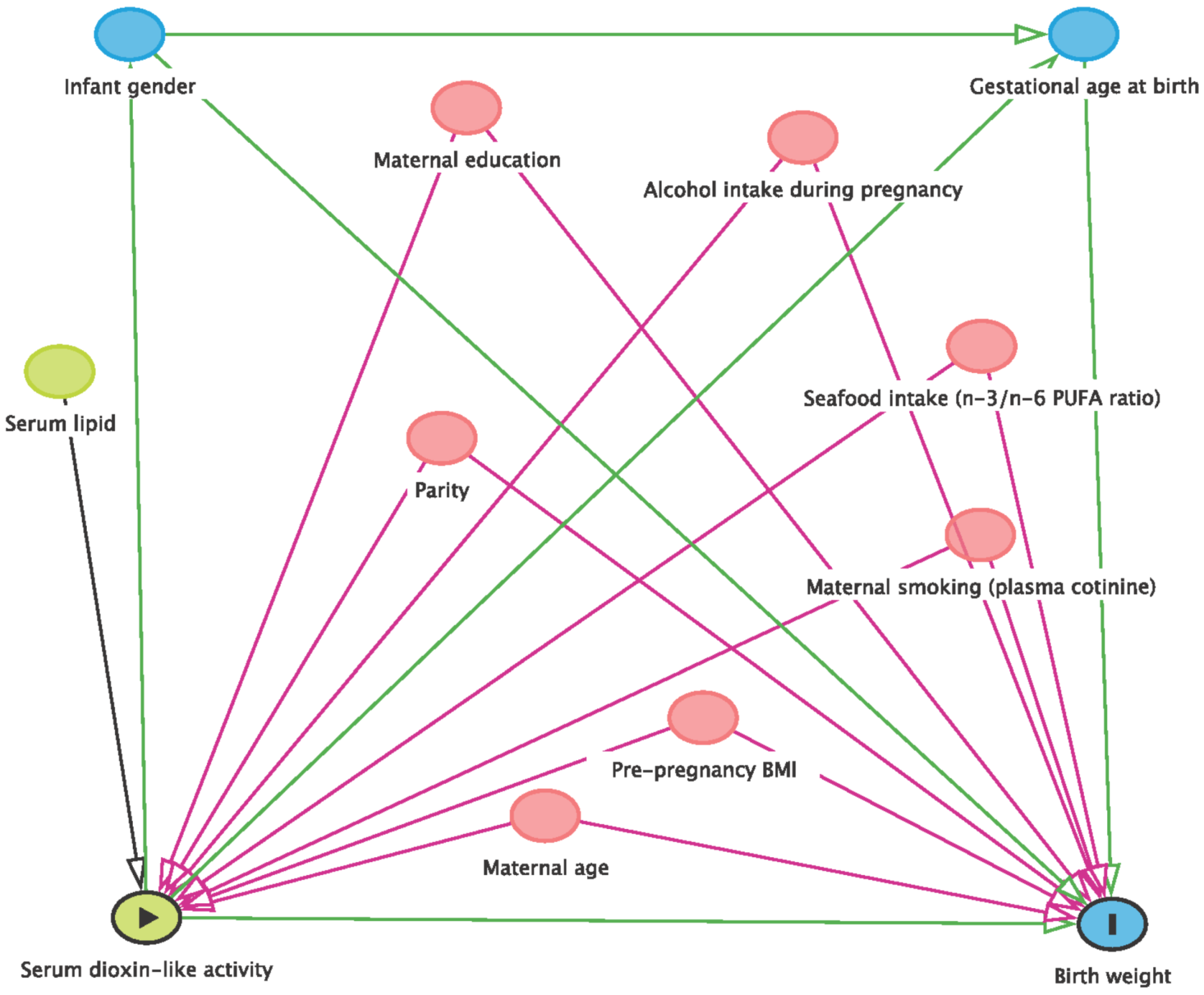
2.8. Statistical Analysis
3. Results
3.1. The Serum Levels of the Combined DL-PCBs and LipPOP Induced Dioxin-like Activity (AhR-TEQ)
3.2. Associations between Dioxin-like Activity and Lipophilic POPs
3.3. Correlation of Maternal Characteristics with Fetal Growth Indices and Serum Dioxin-like Activity
3.4. Association of the Combined Maternal Serum Dioxin-like Activity and Fetal Growth Indices
3.5. Sensitivity Analysis of Association between AhR-TEQ and Fetal Growth Indices
4. Discussion
4.1. Levels of Serum Dioxin-like Activity (AhR-TEQ)
4.2. Determinants of Serum Dioxin-like Activity (AhR-TEQ)
4.3. Associations of Maternal AhR-TEQ with Fetal Growth Indices
4.4. Strength and Limitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACCEPT | Adapting to Climate Change and Environmental Pollution and Dietary Transition |
| AhR | Aryl hydrocarbon receptor |
| AR | Androgen receptor |
| BMI | Body mass index |
| BL | Birth length |
| BW | Birth weight |
| ß-HCH | β-hexachlorocyclohexane |
| DL-PCB | Dioxin-like- polychlorinated biphenyl |
| DLCs | Dioxin-like compounds |
| DMSO | Dimethyl sulfoxide |
| EC50 | Half maximum effect concentration |
| EDCs | Endocrine disrupting chemicals |
| ER | Estrogen receptor |
| GA | Gestational age at birth |
| HC | Head circumference |
| HCB | Hexachlorobenzene |
| Hg | Mercury |
| lipPOPs | Lipophilic persistent organic pollutants |
| LOD | Limit of detection |
| n-3 | Omega-3 fatty acids |
| n-6 | Omega-6 fatty acids |
| OCP | Organochlorine pesticide |
| PAHs | Polycyclic aromatic hydrocarbons |
| PBB | Polybrominated biphenyl |
| PBDE | Polybrominated diphenyl ethers |
| PC | Principal component |
| PCA | Principal component analysis |
| PI | Ponderal index |
| PCB | Polychlorinated biphenyl |
| PCDDs | Polychlorinated dibenzo-p-dioxins |
| PCDFs | Polychlorinated dibenzofurans |
| POP | Persistent organic pollutant |
| p,p′-DDE | Dichlorodiphenyldichloroethylene |
| p,p′-DDT | Dichlorodiphenyltrichloroethane |
| PUFA | Polyunsaturated fatty acid |
| RLU | Relative light unit |
| SC | Solvent control |
| Se | Selenium |
| SPE | Solid phase extraction |
| TCDD | 2,3,7,8-tetrachlorodibenzo-p-dioxin |
| TEF | Toxic Equivalency Factor |
| TEQ | TCDD toxic equivalent |
| TR | Thyroid hormone receptor |
References
- AMAP. Human Health in the Arctic. Arctic Monitoring and Assessment Programme (AMAP). 2002. Available online: http://www.amap.no/documents/index.cfm?dirsub=/AMAP%20Assessment%202002%20-%20Human%20Health%20in%20the%20Arctic (accessed on 11 November 2020).
- Smith, D. Worldwide trends in DDT levels in human breast milk. Int. J. Epidemiol. 1999, 28, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, M.; Birnbaum, L.; Bosveld, A.T.; Brunstrom, B.; Cook, P.; Feeley, M.; Giesy, J.P.; Hanberg, A.; Hasegawa, R.; Kennedy, S.W.; et al. Toxic equivalency factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environ. Health Perspect. 1998, 106, 775–792. [Google Scholar] [CrossRef]
- Barrie, L.A.; Gregor, D.; Hargrave, B.; Lake, R.; Muir, D.; Shearer, R.; Tracey, B.; Bidleman, T. Arctic contaminants: Sources, occurrence and pathways. Sci. Total Environ. 1992, 122, 1–74. [Google Scholar] [CrossRef]
- AMAP. Human Health in the Arctic. Arctic Monitoring and Assessment Programme (AMAP). 2009. Available online: http://www.amap.no/assessment/scientificbackground.htm (accessed on 9 August 2021).
- Bjerregaard, P.; Dewailly, E.; Ayotte, P.; Pars, T.; Ferron, L.; Mulvad, G. Exposure of Inuit in Greenland to organochlorines through the marine diet. J. Toxicol. Environ. Health A 2001, 62, 69–81. [Google Scholar] [CrossRef]
- Deutch, B.; Pedersen, H.S.; Asmund, G.; Hansen, J.C. Contaminants, diet, plasma fatty acids and smoking in Greenland 1999–2005. Sci. Total Environ. 2007, 372, 486–496. [Google Scholar] [CrossRef]
- Deutch, B.; Pedersen, H.S.; Hansen, J.C. Dietary composition in Greenland 2000, plasma fatty acids and persistent organic pollutants. Sci. Total Environ. 2004, 331, 177–188. [Google Scholar] [CrossRef]
- Lauby-Secretan, B.; Loomis, D.; Grosse, Y.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Baan, R.; Mattock, H.; Straif, K. Carcinogenicity of polychlorinated biphenyls and polybrominated biphenyls. Lancet Oncol. 2013, 14, 287–288. [Google Scholar] [CrossRef] [Green Version]
- Steenland, K.; Bertazzi, P.; Baccarelli, A.; Kogevinas, M. Dioxin revisited: Developments since the 1997 IARC classification of dioxin as a human carcinogen. Environ. Health Perspect. 2004, 112, 1265–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindstrom, G.; Hooper, K.; Petreas, M.; Stephens, R.; Gilman, A. Workshop on perinatal exposure to dioxin-like compounds. I. Summary. Environ. Health Perspect. 1995, 103, 135–142. [Google Scholar] [PubMed] [Green Version]
- Rowlands, J.C.; Gustafsson, J.A. Aryl hydrocarbon receptor-mediated signal transduction. Crit. Rev. Toxicol. 1997, 27, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Krishnan, V. Cellular and molecular biology of aryl hydrocarbon (Ah) receptor-mediated gene expression. Arch. Toxicol. Suppl. 1995, 17, 99–115. [Google Scholar] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef]
- Pocar, P.; Fischer, B.; Klonisch, T.; Hombach-Klonisch, S. Molecular interactions of the aryl hydrocarbon receptor and its biological and toxicological relevance for reproduction. Reproduction 2005, 129, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Bonefeld Jorgensen, E.C.; Autrup, H.; Hansen, J.C. Effect of toxaphene on estrogen receptor functions in human breast cancer cells. Carcinogenesis 1997, 18, 1651–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonefeld-Jorgensen, E.C.; Andersen, H.R.; Rasmussen, T.H.; Vinggaard, A.M. Effect of highly bioaccumulated polychlorinated biphenyl congeners on estrogen and androgen receptor activity. Toxicology 2001, 158, 141–153. [Google Scholar] [CrossRef]
- Amaral Mendes, J.J. The endocrine disrupters: A major medical challenge. Food Chem. Toxicol. 2002, 40, 781–788. [Google Scholar] [CrossRef]
- Bonefeld Jorgensen, E.C.; Ayotte, P. Toxicological properties of persistent organic pollutants and related health effects of concern for the arctic populations. In AMAP Assessment 2002: Human Health in the Arctic; Chapter 6; AMAP: Oslo, Norway, 2003; pp. 57–74. [Google Scholar]
- Van Overmeire, I.; Clark, G.C.; Brown, D.J.; Chu, M.D.; Cooke, W.M.; Denison, M.S.; Baeyene, W.; Srebrnik, S.; Goeyens, L. Trace contamination with dioxin-like chemicals: Evaluation of bioassay-based TEQ determination for hazard assessement and regulatory responses. Environ. Sci. Policy 2001, 4, 345–357. [Google Scholar] [CrossRef]
- Long, M.; Laier, P.; Vinggaard, A.M.; Andersen, H.R.; Lynggaard, J.; Bonefeld-Jørgensen, E.C. Effects of currently used pesticides in the AhR-CALUX assay: Comparison between the human TV101L and the rat H4IIE cell line. Toxicology 2003, 194, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Safe, S.H. Polychlorinated biphenyls (PCBs): Environmental impact, biochemical and toxic responses, and implications for risk assessment. Crit. Rev. Toxicol. 1994, 24, 87–149. [Google Scholar] [CrossRef]
- Aarts, J.M.; Denison, M.S.; Cox, M.A.; Schalk, M.A.; Garrison, P.M.; Tullis, K.; de Haan, L.H.; Brouwer, A. Species-specific antagonism of Ah receptor action by 2,2′,5,5′-tetrachloro- and 2,2′,3,3′4,4′-hexachlorobiphenyl. Eur. J. Pharmacol. 1995, 293, 463–474. [Google Scholar] [CrossRef]
- Garrison, P.M.; Tullis, K.; Aarts, J.M.; Brouwer, A.; Giesy, J.P.; Denison, M.S. Species-specific recombinant cell lines as bioassay systems for the detection of 2,3,7,8-tetrachlorodibenzo-p-dioxin-like chemicals. Fundam. Appl. Toxicol. 1996, 30, 194–203. [Google Scholar] [CrossRef]
- Windal, I.; Denison, M.S.; Birnbaum, L.S.; Van Wouwe, N.; Baeyens, W.; Goeyens, L. Chemically activated luciferase gene expression (CALUX) cell bioassay analysis for the estimation of dioxin-like activity: Critical parameters of the CALUX procedure that impact assay results. Environ. Sci. Technol. 2005, 39, 7357–7364. [Google Scholar] [CrossRef]
- Halldorsson, T.I.; Thorsdottir, I.; Meltzer, H.M.; Nielsen, F.; Olsen, S.F. Linking exposure to polychlorinated biphenyls with fatty fish consumption and reduced fetal growth among Danish pregnant women: A cause for concern? Am. J. Epidemiol. 2008, 168, 958–965. [Google Scholar] [CrossRef]
- McGraw, J.E.; Waller, D.P. Fish ingestion and congener specific polychlorinated biphenyl and p,p′-dichlorodiphenyldichloroethylene serum concentrations in a great lakes cohort of pregnant African American women. Environ. Int. 2009, 35, 557–565. [Google Scholar] [CrossRef]
- Tsukimori, K.; Morokuma, S.; Hori, T.; Takahashi, K.; Hirata, T.; Otera, Y.; Fukushima, K.; Kawamoto, T.; Wake, N. Characterization of placental transfer of polychlorinated dibenzo-p-dioxins, dibenzofurans and polychlorinated biphenyls in normal pregnancy. J. Obstet. Gynaecol. Res. 2013, 39, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.R.; Clode, S.; Fan, M.Q.; Fernandes, A.; Foster, P.M.; Jiang, T.; Loizou, G.; MacNicoll, A.; Miller, B.G.; Rose, M.; et al. Toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin in the developing male Wistar(Han) rat. II: Chronic dosing causes developmental delay. Toxicol. Sci. 2007, 99, 224–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golub, M.S.; Donald, J.M.; Reyes, J.A. Reproductive toxicity of commercial PCB mixtures: LOAELs and NOAELs from animal studies. Environ. Health Perspect. 1991, 94, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Majidi, N.; Bouchard, M.; Gosselin, N.H.; Carrier, G. Relationship between prenatal exposure to polychlorinated biphenyls and birth weight: A systematic analysis of published epidemiological studies through a standardization of biomonitoring data. Regul. Toxicol. Pharmacol. 2012, 64, 161–176. [Google Scholar] [CrossRef]
- Konishi, K.; Sasaki, S.; Kato, S.; Ban, S.; Washino, N.; Kajiwara, J.; Todaka, T.; Hirakawa, H.; Hori, T.; Yasutake, D.; et al. Prenatal exposure to PCDDs/PCDFs and dioxin-like PCBs in relation to birth weight. Environ. Res. 2009, 109, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Lundqvist, C.; Zuurbier, M.; Leijs, M.; Johansson, C.; Ceccatelli, S.; Saunders, M.; Schoeters, G.; ten Tusscher, G.; Koppe, J.G. The effects of PCBs and dioxins on child health. Acta Paediatr. 2006, 95, 55–64. [Google Scholar] [CrossRef]
- Nishijo, M.; Tai, P.T.; Nakagawa, H.; Maruzeni, S.; Anh, N.T.; Luong, H.V.; Anh, T.H.; Honda, R.; Morikawa, Y.; Kido, T.; et al. Impact of perinatal dioxin exposure on infant growth: A cross-sectional and longitudinal studies in dioxin-contaminated areas in Vietnam. PLoS ONE 2012, 7, e40273. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Liu, X.; Li, X.; Niu, N.; Yin, X.; Li, N.; Yu, Z. Association between Environmental Dioxin-Related Toxicants Exposure and Adverse Pregnancy Outcome: Systematic Review and Meta-Analysis. Int. J. Fertil. Steril. 2015, 8, 351–366. [Google Scholar] [CrossRef]
- Patandin, S.; Koopman-Esseboom, C.; de Ridder, M.A.; Weisglas-Kuperus, N.; Sauer, P.J. Effects of environmental exposure to polychlorinated biphenyls and dioxins on birth size and growth in Dutch children. Pediatr. Res. 1998, 44, 538–545. [Google Scholar] [CrossRef] [Green Version]
- Tsukimori, K.; Uchi, H.; Mitoma, C.; Yasukawa, F.; Chiba, T.; Todaka, T.; Kajiwara, J.; Yoshimura, T.; Hirata, T.; Fukushima, K.; et al. Maternal exposure to high levels of dioxins in relation to birth weight in women affected by Yusho disease. Environ. Int. 2012, 38, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Hertz-Picciotto, I.; Charles, M.J.; James, R.A.; Keller, J.A.; Willman, E.; Teplin, S. In utero polychlorinated biphenyl exposures in relation to fetal and early childhood growth. Epidemiology 2005, 16, 648–656. [Google Scholar] [CrossRef]
- Sagiv, S.K.; Tolbert, P.E.; Altshul, L.M.; Korrick, S.A. Organochlorine exposures during pregnancy and infant size at birth. Epidemiology 2007, 18, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Sonneborn, D.; Park, H.Y.; Petrik, J.; Kocan, A.; Palkovicova, L.; Trnovec, T.; Nguyen, D.; Hertz-Picciotto, I. Prenatal polychlorinated biphenyl exposures in eastern Slovakia modify effects of social factors on birthweight. Paediatr. Perinat. Epidemiol. 2008, 22, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Tawara, K.; Nishijo, M.; Honda, R.; Maruzeni, S.; Seto, T.; Kido, T.; Saito, S.; Nakagawa, H. Effects of maternal dioxin exposure on newborn size at birth among Japanese mother-infant pairs. Environ. Health Prev. Med. 2009, 14, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Halldorsson, T.I.; Thorsdottir, I.; Meltzer, H.M.; Strøm, M.; Olsen, S.F. Dioxin-like activity in plasma among Danish pregnant women: Dietary predictors, birth weight and infant development. Environ. Res. 2009, 109, 22–28. [Google Scholar] [CrossRef]
- Vafeiadi, M.; Agramunt, S.; Pedersen, M.; Besselink, H.; Chatzi, L.; Fthenou, E.; Fleming, S.; Hardie, L.J.; Wright, J.; Knudsen, L.E.; et al. In utero exposure to compounds with dioxin-like activity and birth outcomes. Epidemiology 2014, 25, 215–224. [Google Scholar] [CrossRef]
- Knudsen, A.K.; Long, M.; Pedersen, H.S.; Bonefeld-Jorgensen, E.C. Lifestyle, reproductive factors and food intake in Greenlandic pregnant women: The ACCEPT—Sub study. Int. J. Circumpolar Health 2015, 74, 29469. [Google Scholar] [CrossRef]
- Long, M.; Knudsen, A.K.; Pedersen, H.S.; Bonefeld-Jorgensen, E.C. Food intake and serum persistent organic pollutants in the Greenlandic pregnant women: The ACCEPT sub-study. Sci. Total Environ. 2015, 529, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Terkelsen, A.S.; Long, M.; Hounsgaard, L.; Bonefeld-Jorgensen, E.C. Reproductive factors, lifestyle and dietary habits among pregnant women in Greenland: The ACCEPT sub-study 2013–2015. Scand. J. Public Health 2017, 252–261. [Google Scholar] [CrossRef] [PubMed]
- CTQ. Test Catalog. Available online: https://www.inspq.qc.ca/en/ctq/test-catalog (accessed on 4 September 2021).
- Bjerregaard-Olesen, C.; Long, M.; Ghisari, M.; Bech, B.H.; Nohr, E.A.; Uldbjerg, N.; Henriksen, T.B.; Olsen, J.; Bonefeld-Jorgensen, E.C. Temporal trends of lipophilic persistent organic pollutants in serum from Danish nulliparous pregnant women 2011–2013. Environ. Sci. Pollut. Res. Int. 2017, 24, 16592–16603. [Google Scholar] [CrossRef]
- Butler Walker, J.; Seddon, L.; McMullen, E.; Houseman, J.; Tofflemire, K.; Corriveau, A.; Weber, J.P.; Mills, C.; Smith, S.; Van Oostdam, J. Organochlorine levels in maternal and umbilical cord blood plasma in Arctic Canada. Sci. Total Environ. 2003, 302, 27–52. [Google Scholar] [CrossRef]
- Bank-Nielsen, P.I.; Long, M.; Bonefeld-Jorgensen, E.C. Pregnant Inuit Women′s Exposure to Metals and Association with Fetal Growth Outcomes: ACCEPT 2010(-)2015. Int. J. Environ. Res. Public Health 2019, 16, 1171. [Google Scholar] [CrossRef] [Green Version]
- Kruger, T.; Long, M.; Ghisari, M.; Bonefeld-Jorgensen, E.C. The combined effect of persistent organic pollutants in the serum POP mixture in Greenlandic Inuit: Xenoestrogenic, xenoandrogenic and dioxin-like transactivities. Biomarkers 2012, 17, 692–705. [Google Scholar] [CrossRef]
- Long, M.; Andersen, B.S.; Lindh, C.H.; Hagmar, L.; Giwercman, A.; Manicardi, G.C.; Bizzaro, D.; Spano, M.; Toft, G.; Pedersen, H.S.; et al. Dioxin-like activities in serum across European and Inuit populations. Environ. Health 2006, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Long, M.; Deutch, B.; Bonefeld-Jorgensen, E.C. AhR transcriptional activity in serum of Inuits across Greenlandic districts. Environ. Health 2007, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Mørck, T.A.; Erdmann, S.E.; Long, M.; Mathiesen, L.; Nielsen, F.; Siersma, V.D.; Bonefeld-Jørgensen, E.C.; Knudsen, L.E. PCB concentrations and dioxin-like activity in blood samples from Danish school children and their mothers living in urban and rural areas. Basic Clin. Pharmacol. Toxicol. 2014, 115, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Rouse, D.J.; Owen, J.; Goldenberg, R.L.; Cliver, S.P. The effectiveness and costs of elective cesarean delivery for fetal macrosomia diagnosed by ultrasound. JAMA 1996, 276, 1480–1486. [Google Scholar] [CrossRef]
- Valvi, D.; Oulhote, Y.; Weihe, P.; Dalgård, C.; Bjerve, K.S.; Steuerwald, U.; Grandjean, P. Gestational diabetes and offspring birth size at elevated environmental pollutant exposures. Environ. Int. 2017, 107, 205–215. [Google Scholar] [CrossRef]
- Villar, J.; Cheikh Ismail, L.; Victora, C.G.; Ohuma, E.O.; Bertino, E.; Altman, D.G.; Lambert, A.; Papageorghiou, A.T.; Carvalho, M.; Jaffer, Y.A.; et al. International standards for newborn weight, length, and head circumference by gestational age and sex: The Newborn Cross-Sectional Study of the INTERGROWTH-21st Project. Lancet 2014, 384, 857–868. [Google Scholar] [CrossRef]
- Miller, H.C. Prenatal factors affecting intrauterine growth retardation. Clin. Perinatol. 1985, 12, 307–318. [Google Scholar] [CrossRef]
- Roje, D.; Banovic, I.; Tadin, I.; Vucinović, M.; Capkun, V.; Barisic, A.; Vulic, M.; Mestrovic, Z.; Mimica, M.; Miletic, T. Gestational age--the most important factor of neonatal ponderal index. Yonsei Med. J. 2004, 45, 273–280. [Google Scholar] [CrossRef] [PubMed]
- WHO. Preterm Birth. Available online: https://www.who.int/news-room/fact-sheets/detail/preterm-birth (accessed on 16 January 2021).
- Textor, J. Drawing and Analyzing Causal DAGs with DAGitty. Available online: https://arxiv.org/abs/1508.04633v1 (accessed on 5 March 2021).
- Albu, A.R.; Anca, A.F.; Horhoianu, V.V.; Horhoianu, I.A. Predictive factors for intrauterine growth restriction. J. Med. Life 2014, 7, 165–171. [Google Scholar] [PubMed]
- Wilcox, A.J.; Weinberg, C.R.; Basso, O. On the pitfalls of adjusting for gestational age at birth. Am. J. Epidemiol. 2011, 174, 1062–1068. [Google Scholar] [CrossRef]
- Carlson, S.E.; Colombo, J.; Gajewski, B.J.; Gustafson, K.M.; Mundy, D.; Yeast, J.; Georgieff, M.K.; Markley, L.A.; Kerling, E.H.; Shaddy, D.J. DHA supplementation and pregnancy outcomes. Am. J. Clin. Nutr. 2013, 97, 808–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koletzko, B.; Lien, E.; Agostoni, C.; Böhles, H.; Campoy, C.; Cetin, I.; Decsi, T.; Dudenhausen, J.W.; Dupont, C.; Forsyth, S.; et al. The roles of long-chain polyunsaturated fatty acids in pregnancy, lactation and infancy: Review of current knowledge and consensus recommendations. J. Perinat. Med. 2008, 36, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meher, A.; Randhir, K.; Mehendale, S.; Wagh, G.; Joshi, S. Maternal Fatty Acids and Their Association with Birth Outcome: A Prospective Study. PLoS ONE 2016, 11, e0147359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newberry, S.J.; Chung, M.; Booth, M.; Maglione, M.A.; Tang, A.M.; O′Hanlon, C.E.; Wang, D.D.; Okunogbe, A.; Huang, C.; Motala, A.; et al. Omega-3 Fatty Acids and Maternal and Child Health: An Updated Systematic Review. Evid. Rep. Technol. Assess 2016, 1–826. [Google Scholar] [CrossRef] [Green Version]
- Robledo, C.A.; Yeung, E.; Mendola, P.; Sundaram, R.; Maisog, J.; Sweeney, A.M.; Barr, D.B.; Louis, G.M. Preconception maternal and paternal exposure to persistent organic pollutants and birth size: The LIFE study. Environ. Health Perspect. 2015, 123, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Kiserud, T.; Benachi, A.; Hecher, K.; Perez, R.G.; Carvalho, J.; Piaggio, G.; Platt, L.D. The World Health Organization fetal growth charts: Concept, findings, interpretation, and application. Am. J. Obstet. Gynecol. 2018, 218, S619–S629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, M.; Bonefeld-Jorgensen, E.C. Dioxin-like activity in environmental and human samples from Greenland and Denmark. Chemosphere 2012, 89, 919–928. [Google Scholar] [CrossRef]
- Abraham, M.; Alramadhan, S.; Iniguez, C.; Duijts, L.; Jaddoe, V.W.; Den Dekker, H.T.; Crozier, S.; Godfrey, K.M.; Hindmarsh, P.; Vik, T.; et al. A systematic review of maternal smoking during pregnancy and fetal measurements with meta-analysis. PLoS ONE 2017, 12, e0170946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Xu, G.; Sun, Y.; Qiu, X.; Ryckman, K.K.; Yu, Y.; Snetselaar, L.G.; Bao, W. Maternal cigarette smoking before and during pregnancy and the risk of preterm birth: A dose-response analysis of 25 million mother-infant pairs. PLoS Med. 2020, 17, e1003158. [Google Scholar] [CrossRef] [PubMed]
- Krüger, T.; Ghisari, M.; Hjelmborg, P.S.; Deutch, B.; Bonefeld-Jorgensen, E.C. Xenohormone transactivities are inversely associated to serum POPs in Inuit. Environ. Health 2008, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Brenerová, P.; Hamers, T.; Kamstra, J.H.; Vondráček, J.; Strapáčová, S.; Andersson, P.L.; Machala, M. Pure non-dioxin-like PCB congeners suppress induction of AhR-dependent endpoints in rat liver cells. Environ. Sci. Pollut. Res. Int. 2016, 23, 2099–2107. [Google Scholar] [CrossRef]
- Doan, T.Q.; Berntsen, H.F.; Verhaegen, S.; Ropstad, E.; Connolly, L.; Igout, A.; Muller, M.; Scippo, M.L. A mixture of persistent organic pollutants relevant for human exposure inhibits the transactivation activity of the aryl hydrocarbon receptor in vitro. Environ. Pollut. 2019, 254, 113098. [Google Scholar] [CrossRef] [Green Version]
- Billionnet, C.; Sherrill, D.; Annesi-Maesano, I. Estimating the health effects of exposure to multi-pollutant mixture. Ann. Epidemiol. 2012, 22, 126–141. [Google Scholar] [CrossRef]
- Newby, P.K.; Tucker, K.L. Empirically derived eating patterns using factor or cluster analysis: A review. Nutr. Rev. 2004, 62, 177–203. [Google Scholar] [CrossRef]
- Donders, A.R.; van der Heijden, G.J.; Stijnen, T.; Moons, K.G. Review: A gentle introduction to imputation of missing values. J. Clin. Epidemiol. 2006, 59, 1087–1091. [Google Scholar] [CrossRef]
- Sterne, J.A.; White, I.R.; Carlin, J.B.; Spratt, M.; Royston, P.; Kenward, M.G.; Wood, A.M.; Carpenter, J.R. Multiple imputation for missing data in epidemiological and clinical research: Potential and pitfalls. BMJ 2009, 338, b2393. [Google Scholar] [CrossRef]
- Bonefeld-Jorgensen, E.C.; Long, M. Dioxin-like activity in the blood of Greenlandic Inuit and Danish women: A pilot study. Int. J. Circumpolar Health 2010, 69, 181–194. [Google Scholar] [CrossRef] [Green Version]
- Bonefeld-Jorgensen, E.C. Biomonitoring in Greenland: Human biomarkers of exposure and effects—A short review. Rural Remote Health 2010, 10, 1362. [Google Scholar] [CrossRef]
- Deutch, B.; Dyerberg, J.; Pedersen, H.S.; Aschlund, E.; Hansen, J.C. Traditional and modern Greenlandic food dietary composition, nutrients and contaminants. Sci. Total Environ. 2007, 384, 106–119. [Google Scholar] [CrossRef]
- Medehouenou, T.C.; Larochelle, C.; Dumas, P.; Dewailly, E.; Ayotte, P. Determinants of AhR-mediated transcriptional activity induced by plasma extracts from Nunavik Inuit adults. Chemosphere 2010, 80, 75–82. [Google Scholar] [CrossRef] [PubMed]
- AMAP. AMAP Assessment 2015: Human Health in the Arctic. Available online: https://www.amap.no/documents/doc/amap-assessment-2015-human-health-in-the-arctic/1346 (accessed on 2 October 2021).
- van den Berg, M.; Kypke, K.; Kotz, A.; Tritscher, A.; Lee, S.Y.; Magulova, K.; Fiedler, H.; Malisch, R. WHO/UNEP global surveys of PCDDs, PCDFs, PCBs and DDTs in human milk and benefit-risk evaluation of breastfeeding. Arch. Toxicol. 2017, 91, 83–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, M.; Halldorsson, T.I.; Mathiesen, L.; Mose, T.; Brouwer, A.; Hedegaard, M.; Loft, S.; Kleinjans, J.C.S.; Besselink, H.; Knudsen, L.E. Dioxin-like exposures and effects on estrogenic and androgenic exposures and micronuclei frequency in mother-newborn pairs. Environ. Int. 2010, 36, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, J.A.; Bazinet, T.M.; Arnason, T.T.; Kimpe, L.E.; Blais, J.M.; White, P.A. Polychlorinated biphenyls (PCBs) contamination and aryl hydrocarbon receptor (AhR) agonist activity of Omega-3 polyunsaturated fatty acid supplements: Implications for daily intake of dioxins and PCBs. Food Chem. Toxicol. 2010, 48, 3093–3097. [Google Scholar] [CrossRef] [PubMed]
- La Merrill, M.; Emond, C.; Kim, M.J.; Antignac, J.P.; Le Bizec, B.; Clément, K.; Birnbaum, L.S.; Barouki, R. Toxicological function of adipose tissue: Focus on persistent organic pollutants. Environ. Health Perspect. 2013, 121, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Lignell, S.; Winkvist, A.; Bertz, F.; Rasmussen, K.M.; Glynn, A.; Aune, M.; Brekke, H.K. Environmental organic pollutants in human milk before and after weight loss. Chemosphere 2016, 159, 96–102. [Google Scholar] [CrossRef]
- Malarvannan, G.; Van Hoorenbeeck, K.; Deguchtenaere, A.; Verhulst, S.L.; Dirinck, E.; Van Gaal, L.; Jorens, P.G.; Covaci, A. Dynamics of persistent organic pollutants in obese adolescents during weight loss. Environ. Int. 2018, 110, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Mocarelli, P.; Warner, M.; Chee, W.Y.; Gerthoux, P.M.; Samuels, S.; Needham, L.L.; Patterson, D.G., Jr. Maternal serum dioxin levels and birth outcomes in women of Seveso, Italy. Environ. Health Perspect. 2003, 111, 947–953. [Google Scholar] [CrossRef]
- Yen, Y.Y.; Lan, S.J.; Yang, C.Y.; Wang, H.H.; Chen, C.N.; Hsieh, C.C. Follow-up study of intrauterine growth of transplacental Yu-Cheng babies in Taiwan. Bull. Environ. Contam. Toxicol. 1994, 53, 633–641. [Google Scholar] [CrossRef]
- Eskenazi, B.; Warner, M.; Brambilla, P.; Signorini, S.; Ames, J.; Mocarelli, P. The Seveso accident: A look at 40 years of health research and beyond. Environ. Int. 2018, 121, 71–84. [Google Scholar] [CrossRef]
- Vartiainen, T.; Jaakkola, J.J.; Saarikoski, S.; Tuomisto, J. Birth weight and sex of children and the correlation to the body burden of PCDDs/PCDFs and PCBs of the mother. Environ. Health Perspect. 1998, 106, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Nishijo, M.; Tawara, K.; Nakagawa, H.; Honda, R.; Kido, T.; Nishijo, H.; Saito, S. 2,3,7,8-Tetrachlorodibenzo-p-dioxin in maternal breast milk and newborn head circumference. J. Exp. Sci. Environ. Epidemiol. 2008, 18, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Main, K.M.; Skakkebaek, N.E.; Virtanen, H.E.; Toppari, J. Genital anomalies in boys and the environment. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Vafeiadi, M.; Agramunt, S.; Papadopoulou, E.; Besselink, H.; Mathianaki, K.; Karakosta, P.; Spanaki, A.; Koutis, A.; Chatzi, L.; Vrijheid, M.; et al. In utero exposure to dioxins and dioxin-like compounds and anogenital distance in newborns and infants. Environ. Health Perspect. 2013, 121, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukimori, K.; Tokunaga, S.; Shibata, S.; Uchi, H.; Nakayama, D.; Ishimaru, T.; Nakano, H.; Wake, N.; Yoshimura, T.; Furue, M. Long-term effects of polychlorinated biphenyls and dioxins on pregnancy outcomes in women affected by the Yusho incident. Environ. Health Perspect. 2008, 116, 626–630. [Google Scholar] [CrossRef] [Green Version]
- Wesselink, A.; Warner, M.; Samuels, S.; Parigi, A.; Brambilla, P.; Mocarelli, P.; Eskenazi, B. Maternal dioxin exposure and pregnancy outcomes over 30 years of follow-up in Seveso. Environ. Int. 2014, 63, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, F.P.; Jedrychowski, W.; Rauh, V.; Whyatt, R.M. Molecular epidemiologic research on the effects of environmental pollutants on the fetus. Environ. Health Perspect. 1999, 107, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowska, M.; Więckowska, B.; Sztorc, L.; Sajdak, S. Smoking and Smoking Cessation in the Risk for Fetal Growth Restriction and Low Birth Weight and Additive Effect of Maternal Obesity. J. Clin. Med. 2020, 9, 3504. [Google Scholar] [CrossRef] [PubMed]
- Kasai, A.; Hiramatsu, N.; Hayakawa, K.; Yao, J.; Maeda, S.; Kitamura, M. High levels of dioxin-like potential in cigarette smoke evidenced by in vitro and in vivo biosensing. Cancer Res. 2006, 66, 7143–7150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtake, F.; Takeyama, K.; Matsumoto, T.; Kitagawa, H.; Yamamoto, Y.; Nohara, K.; Tohyama, C.; Krust, A.; Mimura, J.; Chambon, P.; et al. Modulation of oestrogen receptor signalling by association with the activated dioxin receptor. Nature 2003, 423, 545–550. [Google Scholar] [CrossRef]
- Fierens, S.; Eppe, G.; De Pauw, E.; Bernard, A. Gender dependent accumulation of dioxins in smokers. Occup. Environ. Med. 2005, 62, 61–62. [Google Scholar] [CrossRef] [Green Version]
- Lampa, E.; Eguchi, A.; Todaka, E.; Mori, C. Fetal exposure markers of dioxins and dioxin-like PCBs. Environ. Sci. Pollut. Res. Int. 2018, 25, 11940–11947. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Yin, S.; Zhang, J.; Chen, K.; Jin, M.; Liu, W. Prenatal exposure to polychlorinated biphenyl and umbilical cord hormones and birth outcomes in an island population. Environ. Pollut. 2018, 237, 581–591. [Google Scholar] [CrossRef]
- Ouidir, M.; Buck Louis, G.M.; Kanner, J.; Grantz, K.L.; Zhang, C.; Sundaram, R.; Rahman, M.L.; Lee, S.; Kannan, K.; Tekola-Ayele, F.; et al. Association of Maternal Exposure to Persistent Organic Pollutants in Early Pregnancy With Fetal Growth. JAMA Pediatrics 2019, 174, 149–161. [Google Scholar] [CrossRef]
- Silkworth, J.B.; Antrim, L.; Kaminsky, L.S. Correlations between polychlorinated biphenyl immunotoxicity, the aromatic hydrocarbon locus, and liver microsomal enzyme induction in C57BL/6 and DBA/2 mice. Toxicol. Appl. Pharmacol. 1984, 75, 156–165. [Google Scholar] [CrossRef]
- Heyborne, K.D.; McGregor, J.A.; Henry, G.; Witkin, S.S.; Abrams, J.S. Interleukin-10 in amniotic fluid at midtrimester: Immune activation and suppression in relation to fetal growth. Am. J. Obstet. Gynecol. 1994, 171, 55–59. [Google Scholar] [CrossRef]
- Ohtake, F.; Baba, A.; Fujii-Kuriyama, Y.; Kato, S. Intrinsic AhR function underlies cross-talk of dioxins with sex hormone signalings. Biochem. Biophys. Res. Commun. 2008, 370, 541–546. [Google Scholar] [CrossRef]
- Porterfield, S.P. Thyroidal dysfunction and environmental chemicals--potential impact on brain development. Environ. Health Perspect. 2000, 108 (Suppl. S3), 433–438. [Google Scholar] [PubMed] [Green Version]
- Brouwer, A.; Morse, D.C.; Lans, M.C.; Schuur, A.G.; Murk, A.J.; Klasson-Wehler, E.; Bergman, A.; Visser, T.J. Interactions of persistent environmental organohalogens with the thyroid hormone system: Mechanisms and possible consequences for animal and human health. Toxicol. Ind. Health 1998, 14, 59–84. [Google Scholar] [CrossRef]
- Blazer, S.; Moreh-Waterman, Y.; Miller-Lotan, R.; Tamir, A.; Hochberg, Z. Maternal hypothyroidism may affect fetal growth and neonatal thyroid function. Obstet. Gynecol. 2003, 102, 232–241. [Google Scholar] [CrossRef]
- McKinney, J.D.; Waller, C.L. Molecular determinants of hormone mimicry: Halogenated aromatic hydrocarbon environmental agents. J. Toxicol. Environ. Health B Crit. Rev. 1998, 1, 27–58. [Google Scholar] [CrossRef]
- Bogazzi, F.; Raggi, F.; Ultimieri, F.; Russo, D.; Campomori, A.; McKinney, J.D.; Pinchera, A.; Bartalena, L.; Martino, E. Effects of a mixture of polychlorinated biphenyls (Aroclor 1254) on the transcriptional activity of thyroid hormone receptor. J. Endocrinol. Invest. 2003, 26, 972–978. [Google Scholar] [CrossRef]
- Vintzileos, A.M.; Campbell, W.A.; Neckles, S.; Pike, C.L.; Nochimson, D.J. The ultrasound femur length as a predictor of fetal length. Obstet. Gynecol. 1984, 64, 779–782. [Google Scholar]
- Harvald, B. Genetic epidemiology of Greenland. Clin. Genet. 1989, 36, 364–367. [Google Scholar] [CrossRef]
- Albrecht, E.D.; Pepe, G.J. Estrogen regulation of placental angiogenesis and fetal ovarian development during primate pregnancy. Int. J. Dev. Biol. 2010, 54, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepe, G.J.; Albrecht, E.D. Regulation of functional differentiation of the placental villous syncytiotrophoblast by estrogen during primate pregnancy. Steroids 1999, 64, 624–627. [Google Scholar] [CrossRef]
- Lagiou, P.; Samoli, E.; Hsieh, C.-C.; Lagiou, A.; Xu, B.; Yu, G.-P.; Onoyama, S.; Chie, L.; Adami, H.-O.; Vatten, L.J.; et al. Maternal and cord blood hormones in relation to birth size. Eur. J. Epidemiol. 2014, 29, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Nagata, C.; Iwasa, S.; Shiraki, M.; Shimizu, H. Estrogen and α-Fetoprotein Levels in Maternal and Umbilical Cord Blood Samples in Relation to Birth Weight. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1469–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safe, S.; Wang, F.; Porter, W.; Duan, R.; McDougal, A. Ah receptor agonists as endocrine disruptors: Antiestrogenic activity and mechanisms. Toxicol. Lett. 1998, 102–103, 343–347. [Google Scholar] [CrossRef]
- Suzuki, G.; Nakano, M.; Nakano, S. Distribution of PCDDs/PCDFs and Co-PCBs in human maternal blood, cord blood, placenta, milk, and adipose tissue: Dioxins showing high toxic equivalency factor accumulate in the placenta. Biosci. Biotechnol. Biochem. 2005, 69, 1836–1847. [Google Scholar] [CrossRef] [Green Version]
- Ayotte, P.; Dewailly, E.; Ryan, J.J.; Bruneau, S.; Lebel, G. PCBs and dioxin-like compounds in plasma of adult Inuit living in Nunavik (Arctic Quebec). Chemosphere 1997, 34, 1459–1468. [Google Scholar] [CrossRef]
- Murk, A.J.; Leonards, P.E.G.; Bulder, A.S.; Jonas, A.S.; Rozemeijer, M.J.C.; Denison, M.S.; Koeman, J.H.; Brouwer, A. The CALUX (Chemical-Activated Luciferase Expression) assay adapted and validated for measuring TCDD equivalents in blood plasma. Environ. Toxicol. Chem. 1997, 16, 1583–1589. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
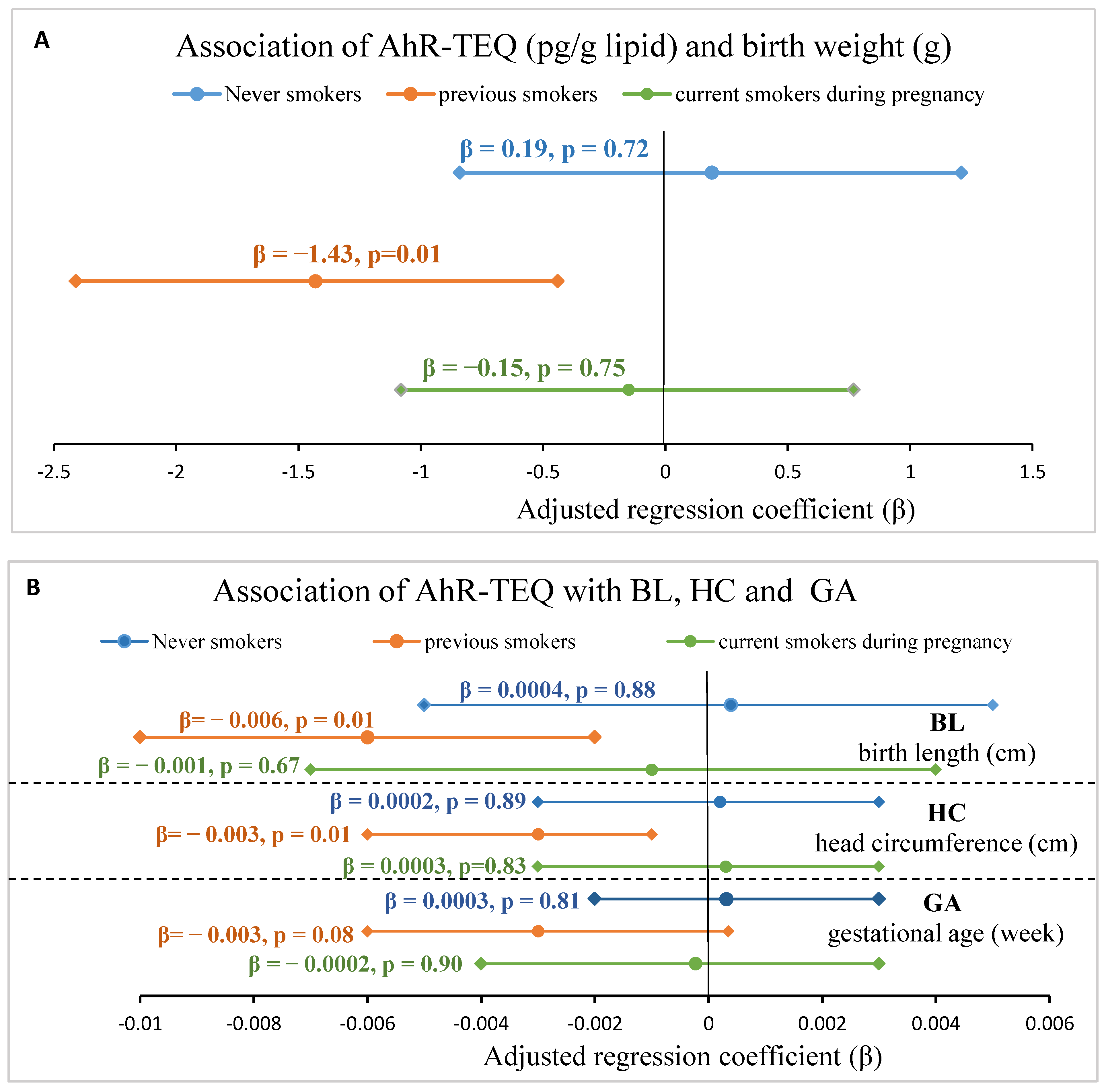{kind=link}
| North | Disko Bay | West | South | East | p | All Greenland Regions | |||
|---|---|---|---|---|---|---|---|---|---|
| POPs (µg/kg serum lipid) | % over LOD | n | 29 | 104 | 255 | 41 | 18 | 447 | |
| PCB105 | 44.4% | Mean ± SD | 3.89 ± 3.34 | 2.13 ± 1.52 | 1.75 ± 1.77 | 1.54 ± 1.02 | 11.9 ± 9.78 | <0.0001 | 2.37 ± 3.29 |
| Median (p25–p75) | 3.00 (1.95–4.95) | 1.85 (1.95–3.28) | 1.00 (1.00–2.20) | 1.00 (0.50–2.10) | 9.30 (4.38–16.5) | 1.60 (1.00–2.60) | |||
| PCB118 | 98.6% | Mean ± SD | 18.9 ± 15.5 | 12.0 ± 7.55 | 9.93 ± 9.87 | 9.01 ± 4.84 | 68.2 ± 66.8 | <0.0001 | 13.2 ± 19.7 |
| Median (p25–p75) | 14.0 (8.90–25.5) | 9.60(6.03–17.0) | 7.30 (4.80–12.0) | 8.80 (4.15–12.0) | 45.0 (23.0–82.8) | 9.00 (5.40–14.0) | |||
| PCB156 | 80.4% | Mean ± SD | 7.05 ± 13.5 | 3.72 ± 2.74 | 4.06 ± 4.04 | 3.90 ± 2.52 | 16.7 ± 15.1 | <0.0001 | 4.67 ± 6.20 |
| Median (p25–p75) | 4.60 (2.30–6.35) | 3.00 (1.90–5.00) | 3.00 (1.80–4.90) | 3.10 (1.95–5.45) | 11.5 (7.15–19.5) | 3.20 (2.00–5.40) | |||
| ∑DL–PCB | Mean ± SD | 29.8 ± 28.9 | 17.8 ± 11.1 | 15.7 ± 14.4 | 14.4 ± 7.20 | 96.8 ± 90.2 | <0.0001 | 20.3 ± 27.8 | |
| Median (p25–p75) | 20.1 (14.2–36.7) | 15.4 (8.85–25.0) | 11.9 (7.60–18.9) | 14.3 (7.24–18.6) | 69.1 (34.1–117) | 13.9 (8.40–21.9) | |||
| ∑PCB | Mean ± SD | 378 ± 489 | 218 ± 125 | 212 ± 219 | 225 ± 108 | 1821 ± 1981 | <0.0001 | 290 ± 545 | |
| Median (p25–p75) | 257 (168–397) | 188 (116–283) | 157 (104–237) | 199 (149–313) | 1181 (603–1857) | 181 (116–282) | |||
| ∑DL–PCB/∑PCB (%) | Mean ± SD | 8.33 ± 1.75 | 8.05 ± 1.72 | 7.56 ± 1.77 | 6.55 ± 1.74 | 6.01 ± 1.45 | <0.0001 | 7.57 ± 1.82 | |
| Median (p25–p75) | 8.53 (7.01–9.28) | 8.19 (6.47–9.17) | 7.42 (6.38–8.76) | 6.48 (5.38–7.32) | 5.96 (4.70–7.46) | 7.41 (6.35–8.88) | |||
| ∑OCP | Mean ± SD | 525 ± 416 | 315 ± 196 | 267 ± 307 | 279 ± 153 | 2781 ± 3493 | <0.0001 | 397 ± 885 | |
| Median (p25–p75) | 416 (266–642) | 271 (161–434) | 198 (127–304) | 256 (160–390) | 1501 (827–2779) | 232 (143–401) | |||
| ∑Legacy POPs | Mean ± SD | 903 ± 857 | 533 ± 314 | 480 ± 520 | 504 ± 253 | 4624 ± 5473 | <0.0001 | 688 ± 1422 | |
| Median (p25–p75) | 674 (452–1024) | 449 (283–713) | 361 (238–529) | 480 (307–709) | 2764 (1390–4652) | 422 (265–686) | |||
| ∑Lip POPs | Mean ± SD | 920 ± 855. | 551 ± 314 | 497 ± 519 | 522 ± 253 | 4552 ± 5328 | <0.0001 | 705 ± 1423 | |
| Median (p25–p75) | 693 (465–1049) | 462 (294–725) | 373 (256–549) | 504 (334–732) | 2866 (1403–4676) | 434 (280–695) | |||
| Dioxin–like activity | |||||||||
| AhR–TEQ (pg/mL serum) | 87.7% | n | 29 | 107 | 259 | 43 | 18 | 0.65 | 456 |
| Mean ± SD | 2.05 ± 2.00 | 1.53 ± 1.29 | 1.79 ± 1.72 | 1.64 ± 1.28 | 1.49 ± 1.52 | 1.72 ± 1.60 | |||
| Median (p25–p75) | 1.67 (0.52–2.90) | 1.29 (0.42–2.22) | 1.32 (0.65–2.41) | 1.32 (0.60–2.62) | 1.38 (0.21–2.17) | 1.32 (0.56–2.40) | |||
| AhR–TEQ (pg/g serum lipid) | 87.7% | n | 29 | 104 | 255 | 41 | 18 | 0.30 | 447 |
| Mean ± SD | 154 ± 194 | 105 ± 101 | 125 ± 123 | 118 ± 101 | 71.9 ± 65.8 | 119 ± 121 | |||
| Median (p25–p75) | 98.2 (36.1–197) | 76.8 (24.2–147) | 90.4 (42.5–163) | 80.1 (35.7–180) | 58.8 (15.8–101) | 86.2 (35.1–160) | |||
| Serum lipid (g/L serum) | n | 29 | 104 | 255 | 41 | 18 | 0.05 | 447 | |
| Mean ± SD | 7.44 ± 1.80 | 7.47 ± 1.92 | 7.40 ± 1.79 | 7.37 ± 1.67 | 8.76 ± 1.66 | 7.47 ± 1.82 | |||
| Median (p25–p75) | 7.60 (5.85–9.15) | 7.10 (6.20–8.10) | 7.30 (6.00–8.50) | 6.90 (6.10–8.50) | 8.70 (7.75–10.0) | 7.30 (6.10–8.60) |
| AhR–TEQ (pg/mL) | AhR–TEQ (pg/g lipid) | ||||||
|---|---|---|---|---|---|---|---|
| n (%) | Mean ± SD | Median (p25–p75) | n | Mean ± SD | Median (p25–p75) | ||
| All | 456 (100) | 1.72 ± 1.60 | 1.32 (0.56–2.40) | 447 (100) | 119 ± 121 | 86.2 (35.1–160) | |
| Maternal age (years) | ≤27 | 239 (52.4) | 1.72 ± 1.63 | 1.35 (0.52–2.33) | 232 (51.9) | 123 ± 130 | 86.6 (32.9–163) |
| >27 | 217 (47.6) | 1.71 ± 1.57 | 1.30 (0.56–2.51) | 215 (48.1) | 115 ± 110 | 85.9 (35.8–157) | |
| Missing | 0 (0.00) | 0 (0.00) | |||||
| p value | 0.99 | 0.84 | |||||
| Adjusted p value a | 0.55 | 0.53 | |||||
| Pre–pregnancy BMI (kg/m2) | Underweight (<18.5) | 12 (2.63) | 2.00 ± 2.30 | 1.37 (0.45–2.47) | 12 (2.68) | 181 ± 261 | 80.7 (27.9–254) |
| Normal weight (18.5–24.9) | 246 (53.9) | 1.87 ± 1.75 | 1.38 (0.68–2.56) | 238 (53.2) | 133 ± 130 | 91.6 (43.2–178) | |
| Overweight (25–30) | 111 (24.3) | 1.48 ± 1.30 | 1.17 (0.44–1.99) | 110 (24.6) | 99.9 ± 95.3 | 70.3 (25.8–134) | |
| Obese (>30) | 73 (16.1) | 1.48 ± 1.31 | 1.17 (0.27–2.13) | 73 (16.3) | 95.9 ± 81.4 | 85.0 (20.7–136) | |
| Missing | 14 (3.07) | 14 (3.13) | |||||
| p value | 0.10 | 0.06 # | |||||
| Adjusted p value b | 0.35 | 0.31 | |||||
| Maternal education | Primary school | 173 (37.9) | 1.63 ± 1.47 | 1.24 (0.56–2.35) | 166 (37.1) | 113 ± 121 | 79.7 (34.5–157) |
| High school | 74 (16.2) | 1.79 ± 1.76 | 1.36 (0.32–2.54) | 73 (16.3) | 127 ± 141 | 90.5 (23.6–159) | |
| Technical college | 130 (28.5) | 1.84 ± 1.85 | 1.37 (0.59–2.47) | 129 (28.9) | 124 ± 122 | 90.4 (39.9–165) | |
| University | 78 (17.1) | 1.67 ± 1.26 | 1.56 (0.59–2.52) | 78 (17.4) | 118 ± 97.4 | 105 (38.7–165) | |
| Missing | 1 (0.22) | 1(0.22) | |||||
| p value | 0.95 | 0.95 | |||||
| Adjusted p value c | 0.85 | 0.91 | |||||
| Maternal smoking history | Never smokers | 114 (25.0) | 1.74 ± 1.60 | 1.28 (0.51–2.39) | 111 (24.8) | 112 ± 122 | 74.4 (25.8–139) |
| Previous smokers | 173 (37.9) | 1.56 ± 1.38 | 1.14 (0.48–2.33) | 172 (38.5) | 120 ± 115 | 89.3 (31.5–163) | |
| Current smokers during pregnancy | 168 (36.8) | 1.86 ± 1.79 | 1.38 (0.65–2.50) | 163 (36.5) | 123 ± 127 | 88.8 (40.6–165) | |
| Missing | 1 (0.22) | 1 (0.22) | |||||
| p value | 0.28 | 0.33 | |||||
| Adjusted p value d | 0.51 | 0.33 | |||||
| Maternal alcohol intake before pregnancy | <1 time a month | 205 (45.0) | 1.65 ± 1.39 | 1.30 (0.61–2.39) | 201 (45.0) | 115 ± 107 | 84.7 (40.9–161) |
| 1 time a month | 71 (15.6) | 1.70 ± 1.52 | 1.35 (0.63–2.51) | 71 (15.9) | 115 ± 116 | 92.5 (37.6–144) | |
| 2–3 times a month | 95 (20.8) | 1.98 ± 2.02 | 1.55 (0.52–2.64) | 93 (20.8) | 132 ± 134 | 92.5 (32.9–175) | |
| ≥1 time a week | 51 (11.2) | 1.63 ± 1.75 | 1.14 (0.55–1.88) | 48 (10.7) | 115 ± 162 | 65.7 (23.2–133) | |
| Missing | 34 (7.46) | 34 (8.50) | |||||
| p value | 0.73 | 0.68 | |||||
| Adjusted p value e | 0.66 | 0.58 | |||||
| Maternal alcohol intake during pregnancy | <1 time a month | 284 (62.3) | 1.75 ± 1.68 | 1.31 (0.64–2.35) | 275 (61.5) | 112 ± 122 | 79.6 (37.5–135) |
| ≥1 time a month | 9 (1.97) | 1.83 ± 1.94 | 0.89 (0.37–2.50) | 9 (2.01) | 98.5 ± 105 | 53.5 (23.7–107) | |
| Missing | 163 (35.7) | 163 (36.5) | |||||
| p value | 0.83 | 0.71 | |||||
| Adjusted p value f | 0.85 | 0.77 | |||||
| Parity | 0 | 176 (38.6) | 2.00 ± 1.93 | 1.53 (0.76–2.62) | 172 (38.5) | 146 ± 147 | 98.6 (52.1–193) |
| 1–2 | 217 (47.6) | 1.52 ± 1.30 | 1.25 (0.48–2.14) | 214 (47.9) | 103 ± 99.2 | 75.6 (25.8–137) | |
| ≥3 | 48 (10.5) | 1.68 ± 1.39 | 1.01 (0.53–3.03) | 46 (10.3) | 100 ± 86.6 | 78.7 (34.5–143) | |
| Missing | 15 (3.29) | 15 (3.36) | |||||
| p value | 0.10 | 0.02 | |||||
| Adjusted p value g | 0.02 | 0.005 | |||||
| AhR–TEQ (pg/mL) | AhR–TEQ (pg/g lipid) | ||||||
|---|---|---|---|---|---|---|---|
| n (%) | Mean ± SD | Median (p25–p75) | n | Mean ± SD | Median (p25–p75) | ||
| All | 456 (100) | 1.72 ± 1.60 | 1.32 (0.56–2.40) | 447 (100) | 119 ± 121 | 86.5 (35.1–160) | |
| Birth weight (g) | Low (<2500) | 16 (3.51) | 1.77 ± 1.45 | 1.83 (0.24–3.09) | 16 (3.58) | 124 ± 107 | 118 (17.7–221) |
| Normal (2500–4500) | 423 (92.8) | 1.72 ± 1.63 | 1.30 (0.56–2.37) | 415 (92.8) | 119 ± 123 | 85.7 (35.0–158) | |
| Macrosomia (≥4500) | 17 (3.73) | 1.59 ± 0.79 | 1.52 (1.08–2.37) | 16 (3.58) | 114 ± 66.0 | 113 (61.5–170) | |
| Missing | 0 (0.00) | 0 (0.00) | |||||
| p value | 0.70 | 0.64 | |||||
| Adjusted p value * | 0.19 | 0.07 # | |||||
| Birth length (cm) | <50 | 81 (17.8) | 2.05 ± 1.95 | 1.67 (0.51–3.03) | 80 (17.9) | 141 ± 153 | 95.5 (31.1–213) |
| ≥50 | 375 (82.2) | 1.65 ± 1.51 | 1.29 (0.56–2.29) | 367 (82.1) | 114 ± 112 | 85.2 (36.9–156) | |
| Missing | 0 (0.00) | ||||||
| p value | 0.33 | 0.44 | |||||
| Adjusted p value * | 0.05 | 0.05 | |||||
| Ponderal Index (g/cm3) | Normal (2.2–3.0) | 407 (89.3) | 1.75 ± 1.65 | 1.31 (0.57–2.47) | 398 (89.0) | 122 ± 125 | 85.8 (36.6–164) |
| Abnormal (<2.2 or >3.0) | 48 (10.5) | 1.39 ± 1.00 | 1.45 (0.49–1.98) | 48 (10.7) | 93.8 ± 70.5 | 91.9 (26.4–131) | |
| Missing | 1 (0.22) | 1 (0.22) | |||||
| p value | 0.16 | 0.20 | |||||
| Adjusted p value § | 0.17 | 0.22 | |||||
| Birth head circumference (cm) | <35 | 202 (44.3) | 1.73 ± 1.72 | 1.31 (0.44–2.51) | 196 (43.8) | 116 ± 124 | 79.7 (28.7–159) |
| ≥35 | 253 (55.5) | 1.71 ± 1.51 | 1.34 (0.63–2.37) | 250 (55.9) | 121 ± 119 | 89.7 (38.9–161) | |
| Missing | 1 (0.22) | 1 (0.22) | |||||
| p value | 0.32 | 0.15 | |||||
| Adjusted p value * | 0.26 | 0.17 | |||||
| Gestational age at birth (weeks) | Preterm birth (<37) | 24 (5.26) | 1.49 ± 1.83 | 0.89 (0.22–2.12) | 24 (5.37) | 93.8 ± 120 | 44.1 (7.36–451) |
| Term birth (≥37) | 417 (91.5) | 1.75 ± 1.60 | 1.38 (0.59–2.44) | 408 (91.3) | 122 ± 122 | 89.1 (37.4–163) | |
| Missing | 15 (5.86) | 15 (3.36) | |||||
| p value | 0.08 # | 0.05 | |||||
| Adjusted p value * | 0.16 | 0.11 | |||||
| Gender | Male | 241 (52.9) | 1.70 ± 1.68 | 1.24 (0.54–2.37) | 236 (52.8) | 119 ± 123 | 85.1 (37.1–159) |
| Female | 214 (46.9) | 1.73 ± 1.51 | 1.42 (0.56–2.42) | 210 (47.0) | 119 ± 118 | 88.8 (33.9–160) | |
| Missing | 1 (0.22) | 1 (0.22) | |||||
| p value | 0.89 | 0.85 | |||||
| Adjusted p value * | 0.58 | 0.62 | |||||
| AhR–TEQ (pg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Crude | Adjusted a | Adjusted b | |||||||
| n | β (95% CI) | p | n | β (95% CI) | p | n | β (95% CI) | p | |
| PCB105 (µg/L) | 465 | 1.65 (−3.56; 6.85) | 0.53 | 465 | 1.54 (–3.94; 7.01) | 0.58 | 258 | 6.00 (–4.24; 16.2) | 0.25 |
| PCB118 (µg/L) | 465 | 0.16 (–0.73; 1.04) | 0.73 | 465 | 0.13 (–0.80; 1.06) | 0.79 | 258 | 0.96 (–1.17; 3.09) | 0.38 |
| PCB156 (µg/L) | 465 | 2.30 (–0.55; 5.15) | 0.11 | 465 | 2.39 (–0.61; 5.39) | 0.12 | 258 | 6.59 (1.22; 12.0) | 0.02 |
| ΣDL–PCB (µg/L) | 465 | 0.20 (–0.41; 0.83) | 0.51 | 465 | 0.20 (–0.45; 0.86) | 0.55 | 258 | 1.04 (–0.41; 2.49) | 0.16 |
| ΣPCB (µg/L) | 465 | 0.01 (–0.03; 0.04) | 0.72 | 465 | 0.01 (–0.03; 0.04) | 0.77 | 258 | 0.09 (–0.02; 0.20) | 0.11 |
| ΣDL–PCB / ΣPCB | 465 | 5.47 (–2.61; 13.6) | 0.18 | 465 | 5.44 (–2.65; 13.5) | 0.19 | 258 | 9.93 (–2.42; 22.3) | 0.11 |
| HCB (µg/L) | 465 | 0.16 (–0.50; 0.81) | 0.64 | 465 | 0.14 (–0.56; 0.84) | 0.70 | 258 | 0.38 (–0.74; 1.50) | 0.50 |
| β–HCH (µg/L) | 464 | 0.64 (–2.19; 3.47) | 0.66 | 464 | 0.59 (–2.41; 3.59) | 0.70 | 258 | 2.77 (–3.16; 8.70) | 0.36 |
| p,p’–DDE (µg/L) | 465 | 0.00 (–0.03; 0.03) | 0.98 | 465 | –0.001 (–0.03; 0.03) | 0.94 | 258 | 0.09 (–0.06; 0.24) | 0.24 |
| Oxychlordane (µg/L) | 465 | 0.04 (–0.20; 0.28) | 0.74 | 465 | 0.03 (–0.21; 0.28) | 0.79 | 258 | 0.65 (–0.08; 1.37) | 0.08 # |
| Transnonachlor (µg/L) | 465 | 0.03 (–0.11; 0.16) | 0.72 | 465 | 0.02 (–0.12; 0.16) | 0.77 | 258 | 0.33 (–0.12; 0.78) | 0.15 |
| Mirex (µg/L) | 465 | 0.45 (–1.25; 2.15) | 0.60 | 465 | 0.42 (–1.34; 2.17) | 0.64 | 258 | 3.91 (–0.89; 8.71) | 0.11 |
| ΣOCP (µg/L) | 464 | 0.001 (–0.02; 0.02) | 0.92 | 464 | 0.00 (–0.02; 0.02) | 0.97 | 258 | 0.06 (–0.03; 0.14) | 0.20 |
| ΣLegacyPOP (µg/L) | 464 | 0.001 (–0.01; 0.01) | 0.85 | 463 | 0.001 (–0.01; 0.01) | 0.89 | 257 | 0.04 (–0.01; 0.09) | 0.15 |
| ΣLipPOPs (µg/L) | 464 | 0.001 (–0.01; 0.01) | 0.84 | 463 | 0.001 (–0.01; 0.01) | 0.66 | 257 | 0.04 (–0.01; 0.09) | 0.20 |
| PCA component | |||||||||
| PC–1 (lipPOP, ug/L) | 457 | 0.04 (–0.10; 0.17) | 0.60 | 253 | 0.25 (–0.16; 0.66) | 0.24 | 253 | 0.31 (–0.13; 0.76) | 0.17 |
| PC–2 (Se and Hg, ug/L) | 457 | –0.03 (–0.17; 0.11) | 0.63 | 253 | 0.07 (–0.13; 0.26) | 0.48 | 253 | 0.08 (–0.12; 0.27) | 0.45 |
| Birth Weight (BW) (gram) | Birth Length (BL) (cm) | Ponderal Index (PI) (g/cm3) | Head Circumference (HC) (cm) | Gestational Age at Birth (GA) (Week) | AhR–TEQ (pg/mL Serum) | AhR–TEQ (pg/g Serum Lipid) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | rs | p | n | rs | p | n | rs | p | n | rs | p | n | rs | p | n | rs | p | n | rs | p | |
| Age (years) | 456 | 0.05 | 0.25 | 456 | 0.06 | 0.20 | 456 | 0.02 | 0.66 | 455 | 0.08 | 0.11 | 441 | 0.03 | 0.49 | 456 | 0.04 | 0.40 | 447 | 0.02 | 0.76 |
| Pre–pregnancy BMI (kg/m2) | 442 | 0.15 | 0.002 | 442 | 0.11 | 0.03 | 442 | 0.11 | 0.02 | 441 | 0.12 | 0.01 | 428 | 0.04 | 0.38 | 442 | –0.11 | 0.02 | 433 | –0.11 | 0.02 |
| Plasma cotinine (ng/mL) | 444 | –0.25 | <0.0001 | 444 | –0.26 | 0.0001 | 444 | –0.03 | 0.50 | 443 | –0.24 | <0.0001 | 432 | –0.15 | 0.002 | 444 | 0.05 | 0.27 | 435 | 0.04 | 0.47 |
| Smoking history | 455 | –0.18 | <0.0001 | 455 | –0.24 | <0.0001 | 455 | 0.03 | 0.56 | 454 | –0.16 | <0.0001 | 440 | –0.12 | 0.01 | 455 | 0.04 | 0.44 | 446 | 0.05 | 0.26 |
| n–3/n–6 PUFA ratio | 455 | 0.04 | 0.38 | 455 | 0.03 | 0.58 | 455 | –0.01 | 0.84 | 454 | –0.04 | 0.43 | 440 | 0.09 | 0.05 | 455 | 0.06 | 0.20 | 446 | 0.10 | 0.04 |
| Alcohol intake | |||||||||||||||||||||
| before pregnancy | 422 | –0.03 | 0.57 | 422 | –0.02 | 0.64 | 422 | –0.04 | 0.46 | 421 | –0.03 | 0.53 | 407 | 0.03 | 0.49 | 422 | 0.003 | 0.95 | 413 | –0.02 | 0.72 |
| during pregnancy | 286 | –0.08 | 0.20 | 286 | –0.09 | 0.12 | 286 | –0.01 | 0.85 | 285 | –0.13 | 0.03 | 273 | –0.04 | 0.49 | 286 | –0.05 | 0.40 | 278 | –0.06 | 0.30 |
| Parity | 441 | 0.02 | 0.73 | 441 | –0.02 | 0.73 | 441 | 0.07 | 0.17 | 441 | 0.04 | 0.46 | 427 | –0.04 | 0.43 | 441 | –0.07 | 0.16 | 432 | –0.11 | 0.02 |
| Maternal education | 455 | 0.10 | 0.04 | 455 | 0.10 | 0.03 | 455 | 0.02 | 0.61 | 454 | 0.14 | 0.002 | 440 | 0.07 | 0.17 | 455 | 0.04 | 0.40 | 446 | 0.06 | 0.40 |
| Gestational age (GA) (week) | 457 | 0.51 | <0.0001 | 441 | 0.46 | <0.0001 | 441 | 0.19 | <0.0001 | 440 | 0.41 | <0.0001 | – | – | – | 441 | –0.05 | 0.29 | 432 | 0.001 | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, M.; Wielsøe, M.; Bonefeld-Jørgensen, E.C. Dioxin-like Activity in Pregnant Women and Indices of Fetal Growth: The ACCEPT Birth Cohort. Toxics 2022, 10, 26. https://doi.org/10.3390/toxics10010026
Long M, Wielsøe M, Bonefeld-Jørgensen EC. Dioxin-like Activity in Pregnant Women and Indices of Fetal Growth: The ACCEPT Birth Cohort. Toxics. 2022; 10(1):26. https://doi.org/10.3390/toxics10010026
Chicago/Turabian StyleLong, Manhai, Maria Wielsøe, and Eva Cecilie Bonefeld-Jørgensen. 2022. "Dioxin-like Activity in Pregnant Women and Indices of Fetal Growth: The ACCEPT Birth Cohort" Toxics 10, no. 1: 26. https://doi.org/10.3390/toxics10010026
APA StyleLong, M., Wielsøe, M., & Bonefeld-Jørgensen, E. C. (2022). Dioxin-like Activity in Pregnant Women and Indices of Fetal Growth: The ACCEPT Birth Cohort. Toxics, 10(1), 26. https://doi.org/10.3390/toxics10010026