
Cytotoxicity Mechanisms of Eight Major Herbicide Active Ingredients in Comparison to Their Commercial Formulations




Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Mammalian Cell Tissue Culture
2.3. Cytotoxicity Assays
2.4. ToxiLight Cell Membrane Damage Assay
2.5. Oxidative Stress
2.6. ToxTracker Assay System
2.7. Statistical Analysis
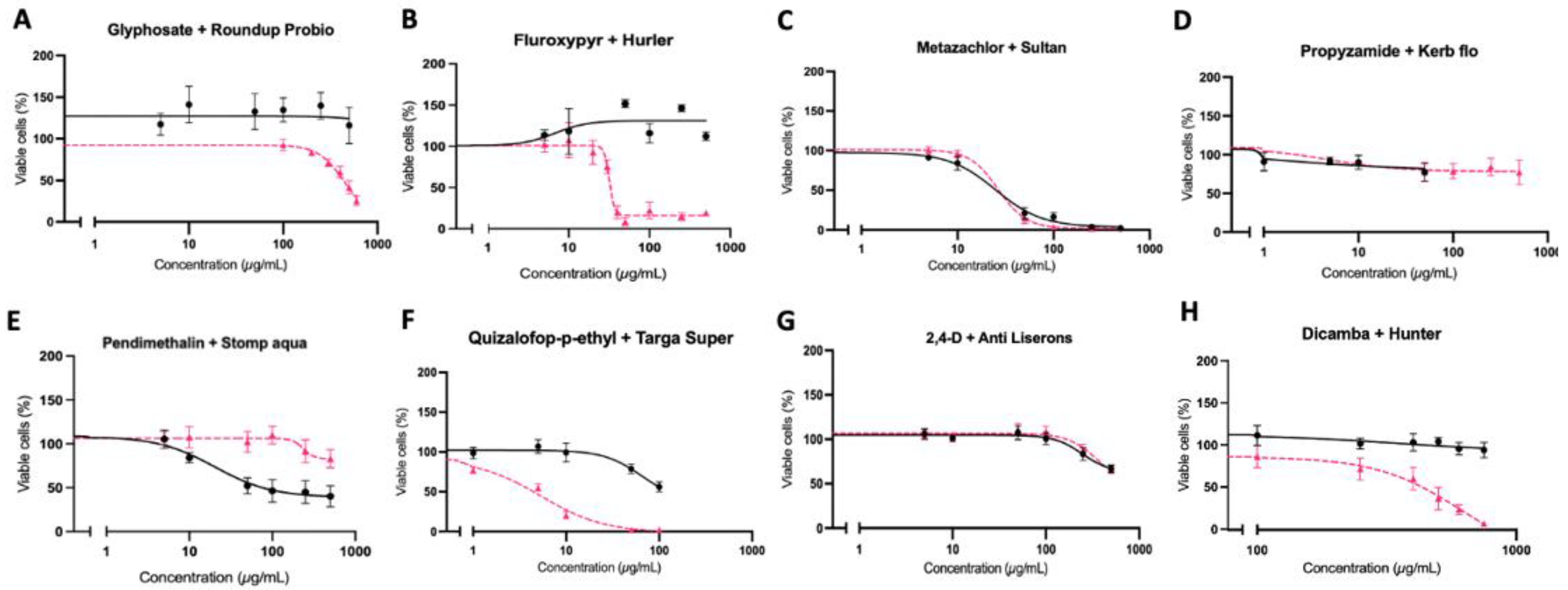
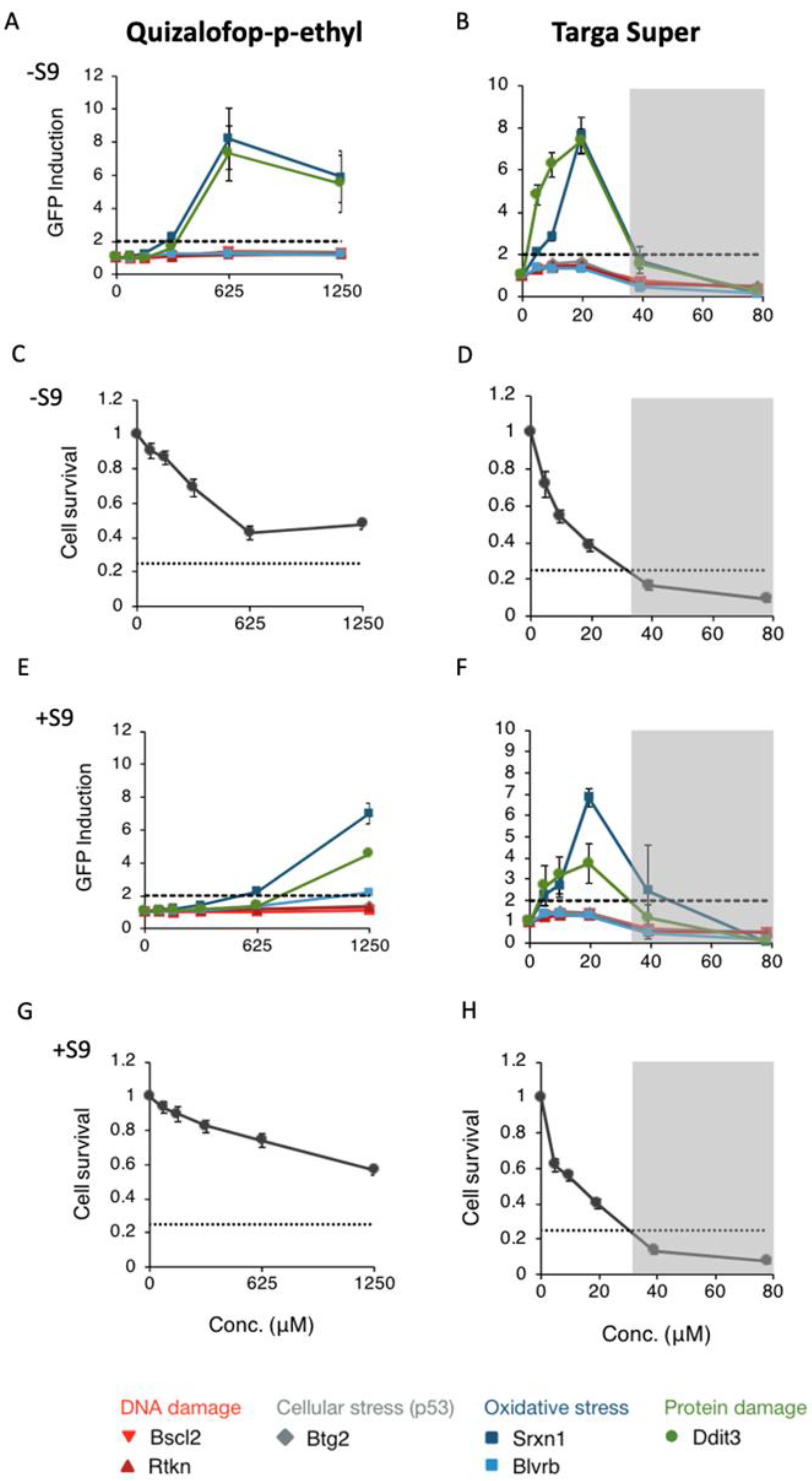
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Multigner, L.; Ndong, J.; Giusti, A.; Romana, M.; Delacroix-Maillard, H.; Cordier, S.; Jégou, B.; Thome, J.; Blanchet, P. Chlordecone Exposure and Risk of Prostate Cancer. J. Clin. Oncol. 2010, 28, 3457–3462. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; van Gent, D.; Incrocci, L.; van Weerden, W.; Nonnekens, J. Role of the DNA damage response in prostate cancer formation, progression and treatment. Prostate Cancer Prostatic Dis. 2019, 23, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Tanner, C.; Kamel, F.; Ross, G.; Hoppin, J.; Goldman, S.; Korell, M.; Marras, C.; Bhudhikanok, G.; Kasten, M.; Chade, A.; et al. Rotenone, Paraquat, and Parkinson’s Disease. Environ. Health Perspect. 2011, 119, 866–872. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Zhang, Y.; Liu, L.; Yan, H. Pesticide exposure and risk of Alzheimer’s disease: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 32222. [Google Scholar] [CrossRef] [Green Version]
- Potashnik, G.; Porath, A. Dibromochloropropane (DBCP): A 17-year Reassessment of Testicular Function and Reproductive Performance. J. Occup. Environ. Med. 1995, 37, 287–1292. [Google Scholar] [CrossRef]
- González-Alzaga, B.; Hernández, A.; Rodríguez-Barranco, M.; Gómez, I.; Aguilar-Garduño, C.; López-Flores, I.; Parrón, T.; Lacasaña, M. Pre- and postnatal exposures to pesticides and neurodevelopmental effects in children living in agricultural communities from South-Eastern Spain. Environ. Int. 2015, 85, 229–237. [Google Scholar] [CrossRef]
- Gunier, R.; Bradman, A.; Harley, K.; Kogut, K.; Eskenazi, B. Prenatal Residential Proximity to Agricultural Pesticide Use and IQ in 7-Year-Old Children. Environ. Health Perspect. 2017, 125, 057002. [Google Scholar] [CrossRef] [Green Version]
- Wigle, D.; Turner, M.; Krewski, D. A Systematic Review and Meta-analysis of Childhood Leukemia and Parental Occupational Pesticide Exposure. Environ. Health Perspect. 2009, 117, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Martenies, S.; Perry, M. Environmental and occupational pesticide exposure and human sperm parameters: A systematic review. Toxicology. 2013, 307, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Rana, I.; Shaffer, R.; Taioli, E.; Sheppard, L. Exposure to glyphosate-based herbicides and risk for non-Hodgkin lymphoma: A meta-analysis and supporting evidence. Mutat. Res. Rev. Mutat. 2019, 781, 186–206. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Antoniou, M. Ignoring Adjuvant Toxicity Falsifies the Safety Profile of Commercial Pesticides. Front. Public Health 2018, 5, 361. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Defarge, N.; Spiroux de Vendômois, J.; Séralini, G. Major Pesticides Are More Toxic to Human Cells Than Their Declared Active Principles. Biomed Res. In. 2014, 2014, 179691. [Google Scholar] [CrossRef] [Green Version]
- Guyton, K.; Loomis, D.; Grosse, Y.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H.; Straif, K. Carcinogenicity of tetrachlorvinphos, parathion, malathion, diazinon, and glyphosate. Lancet Oncol. 2015, 16, 490–491. [Google Scholar] [CrossRef]
- Spinaci, M.; Nerozzi, C.; Tamanini, C.; Bucci, D.; Galeati, G. Glyphosate and Its Formulation Roundup Impair Pig Oocyte Maturation. Sci. Rep. 2020, 10, 12007. [Google Scholar] [CrossRef]
- Mesnage, R.; Ferguson, S.; Brandsma, I.; Moelijker, N.; Zhang, G.; Mazzacuva, F.; Caldwell, A.; Halket, J.; Antoniou, M. The surfactant co-formulant POEA in the glyphosate-based herbicide RangerPro but not glyphosate alone causes necrosis in Caco-2 and hepg2 human cell lines and ER stress in the toxtracker assay. Food Chem. Toxicol. 2022, 168, 113380. [Google Scholar] [CrossRef]
- Nagy, K.; Tessema, R.A.; Budnik, L.T.; Ádám, B. Comparative Cyto- and Genotoxicity Assessment of Glyphosate and Glyphosate-Based Herbicides in Human Peripheral White Blood Cells. Environ. Res. 2019, 179, 108851. [Google Scholar] [CrossRef]
- Oláh, M.; Farkas, E.; Székács, I.; Horvath, R.; Székács, A. Cytotoxic Effects of Roundup Classic and Its Components on NE-4C and MC3T3-E1 Cell Lines Determined by Biochemical and Flow Cytometric Assays. Toxicol. Rep. 2022, 9, 914–926. [Google Scholar] [CrossRef]
- Mesnage, R.; Bernay, B.; Séralini, G. Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology 2013, 313, 122–128. [Google Scholar] [CrossRef]
- Spinaci, M.; Nerozzi, C.; Mislei, B.; Blanco-Prieto, O.; Mari, G.; Galeati, G.; Bucci, D. Impact of Glyphosate and Its Formulation Roundup® on Stallion Spermatozoa. Theriogenology 2022, 179, 197–203. [Google Scholar] [CrossRef]
- Clair, É.; Mesnage, R.; Travert, C.; Séralini, G. A Glyphosate-Based Herbicide Induces Necrosis and Apoptosis in Mature Rat Testicular Cells in Vitro, and Testosterone Decrease at Lower Levels. Toxicol. Vitro 2012, 26, 269–279. [Google Scholar] [CrossRef]
- Manservisi, F.; Lesseur, C.; Panzacchi, S.; Mandrioli, D.; Falcioni, L.; Bua, L.; Manservigi, M.; Spinaci, M.; Galeati, G.; Mantovani, A.; et al. The Ramazzini Institute 13-Week Pilot Study Glyphosate-Based Herbicides Administered at Human-Equivalent Dose to Sprague Dawley Rats: Effects on Development and Endocrine System. Environ. Health. 2019, 18, 15. [Google Scholar] [CrossRef] [Green Version]
- El-Shenawy, N.S. Oxidative Stress Responses of Rats Exposed to Roundup and Its Active Ingredient Glyphosate. Environ. Toxicol. Pharmacol. 2009, 28, 379–385. [Google Scholar] [CrossRef]
- Mesnage, R.; Ibragim, M.; Mandrioli, D.; Falcioni, L.; Tibaldi, E.; Belpoggi, F.; Brandsma, I.; Bourne, E.; Savage, E.; Mein, C.; et al. Comparative Toxicogenomics of Glyphosate and Roundup Herbicides by Mammalian Stem Cell-Based Genotoxicity Assays and Molecular Profiling in Sprague-Dawley Rats. Toxicol. Sci. 2021, 186, 83–101. [Google Scholar] [CrossRef]
- Mesnage, R.; Panzacchi, S.; Bourne, E.; Mein, C.A.; Perry, M.J.; Hu, J.; Chen, J.; Mandrioli, D.; Belpoggi, F.; Antoniou, M. Glyphosate and Its Formulations Roundup Bioflow and RangerPro Alter Bacterial and Fungal Community Composition in the Rat Caecum Microbiome. Front. Microbiol. 2022, 13, 888853. [Google Scholar] [CrossRef]
- Mesnage, R.; Renney, G.; Séralini, G.; Ward, M.; Antoniou, M. Multiomics reveal non-alcoholic fatty liver disease in rats following chronic exposure to an ultra-low dose of Roundup herbicide. Sci. Rep. 2017, 7, 39328. [Google Scholar] [CrossRef] [Green Version]
- González, M.; Soloneski, S.; Reigosa, M.; Larramendy, M. Genotoxicity of the herbicide 2,4-dichlorophenoxyacetic and a commercial formulation, 2,4-dichlorophenoxyacetic acid dimethylamine salt. I. Evaluation of DNA damage and cytogenetic endpoints in Chinese Hamster ovary (CHO) cells. Toxicol. Vitro 2005, 19, 289–297. [Google Scholar] [CrossRef]
- Yin, X.; Li, S.; Zhang, L.; Zhu, G.; Zhuang, H. Evaluation of DNA damage in Chinese toad (Bufo bufo gargarizans) after in vivo exposure to sublethal concentrations of four herbicides using the comet assay. Ecotoxicology 2008, 17, 280–286. [Google Scholar] [CrossRef]
- Nikoloff, N.; Larramendy, M.; Soloneski, S. Assessment of DNA damage, cytotoxicity, and apoptosis in human hepatoma (HepG2) cells after flurochloridone herbicide exposure. Food Chem. Toxicol. 2014, 65, 233–241. [Google Scholar] [CrossRef]
- Wang, C.; Harwood, J.; Zhang, Q. Oxidative stress and DNA damage in common carp (Cyprinus carpio) exposed to the herbicide mesotrione. Chemosphere 2018, 193, 1080–1086. [Google Scholar] [CrossRef]
- Li, M.; Ma, X.; Saleem, M.; Wang, X.; Sun, L.; Yang, Y.; Zhang, Q. Biochemical response, histopathological change and DNA damage in earthworm (Eisenia fetida) exposed to sulfentrazone herbicide. Ecol. Indic. 2020, 115, 106465. [Google Scholar] [CrossRef]
- Mesnage, R.; Brandsma, I.; Moelijker, N.; Zhang, G.; Antoniou, M. Genotoxicity evaluation of 2,4-D, dicamba and glyphosate alone or in combination with cell reporter assays for DNA damage, oxidative stress and unfolded protein response. Food Chem. Toxicol. 2021, 157, 112601. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- DEFRA. Pesticide Usage Survey Report 295 Arable Crops in the United Kingdom 2020. 2021. Available online: https://pusstats.fera.co.uk/published-reports (accessed on 4 June 2022).
- Hendriks, G.; Derr, R.S.; Misovic, B.; Morolli, B.; Calléja, F.M.G.R.; Vrieling, H. The extended ToxTracker assay discriminates between induction of DNA damage, oxidative stress, and protein misfolding. Toxicol. Sci. 2016, 150, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Arici, M.; Abudayyak, M.; Boran, T.; Özhan, G. Does pendimethalin develop in pancreatic cancer induced inflammation? Chemosphere 2020, 252, 126644. [Google Scholar] [CrossRef]
- Saquib, Q.; Siddiqui, M.; Ansari, S.; Alwathnani, H.; Musarrat, J.; Al-Khedhairy, A. Cytotoxicity and genotoxicity of methomyl, carbaryl, metalaxyl, and pendimethalin in human umbilical vein endothelial cells. J. Appl. Toxicol. 2021, 41, 832–846. [Google Scholar] [CrossRef]
- Gasnier, C.; Dumont, C.; Benachour, N.; Clair, E.; Chagnon, M.; Séralini, G. Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology 2009, 262, 184–191. [Google Scholar] [CrossRef]
- Gasnier, C.; Laurant, C.; Decroix-Laporte, C.; Mesnage, R.; Clair, E.; Travert, C.; Séralini, G. Defined plant extracts can protect human cells against combined xenobiotic effects. J. Occup. Med. Toxicol. 2011, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Biserni, M.; Mesnage, R.; Ferro, R.; Wozniak, E.; Xenakis, T.; Mein, C.; Antoniou, M. Quizalofop-p-Ethyl Induces Adipogenesis in 3T3-L1 Adipocytes. Toxicol. Sci. 2019, 170, 452–461. [Google Scholar] [CrossRef] [Green Version]
- González, N.V.; Soloneski, S.; Larramendy, M.L. The Chlorophenoxy Herbicide Dicamba and Its Commercial Formulation Banvel® Induce Genotoxicity and Cytotoxicity in Chinese Hamster Ovary (CHO) Cells. Mutat. Res.-Genet. Toxicol. Environ. Mutagen. 2007, 634, 60–68. [Google Scholar] [CrossRef]
- Mckee, R.; Wong, Z.; Schmitt, S.; Beatty, P.; Swanson, M.; Schreiner, C.; Schardein, J. The Reproductive and Developmental Toxicity of High Flash Aromatic Naphtha. Toxicol. Ind. Health 1990, 6, 441–460. [Google Scholar] [CrossRef]
- Chang, T.; Jeong, W.; Woo, H.; Lee, S.; Park, S.; Rhee, S. Characterization of Mammalian Sulfiredoxin and Its Reactivation of Hyperoxidized Peroxiredoxin through Reduction of Cysteine Sulfinic Acid in the Active Site to Cysteine. J. Biol. Chem. 2004, 279, 50994–51001. [Google Scholar] [CrossRef] [Green Version]
- Socianu, S.; Bopp, S.; Govarts, E.; Gilles, L.; Buekers, J.; Kolossa-Gehring, M.; Backhaus, T.; Franco, A. Chemical Mixtures in the EU Population: Composition and Potential Risks. Int. J. Environ. Rese. Public Health. 2022, 19, 6121. [Google Scholar] [CrossRef]
- Mesnage, R.; Bowyer, R.; El Balkhi, S.; Saint-Marcoux, F.; Gardere, A.; Ducarmon, Q.; Geelen, A.; Zwittink, R.; Tsoukalas, D.; Sarandi, E.; et al. Impacts of dietary exposure to pesticides on faecal microbiome metabolism in adult twins. Environ. Health 2022, 21, 46. [Google Scholar] [CrossRef]
- Behrens, R.; Lueschen, W. Dicamba Volatility. Weed Sci. 1979, 27, 486–493. [Google Scholar] [CrossRef]
- Vardakas, P.; Veskoukis, A.; Rossiou, D.; Gournikis, C.; Kapetanopoulou, T.; Karzi, V.; Docea, A.; Tsatsakis, A.; Kouretas, D. A Mixture of Endocrine Disruptors and the Pesticide Roundup® Induce Oxidative Stress in Rabbit Liver When Administered under the Long-Term Low-Dose Regimen: Reinforcing the Notion of Real-Life Risk Simulation. Toxics. 2022, 10, 190. [Google Scholar] [CrossRef]
- Isling, L.; Boberg, J.; Jacobsen, P.; Mandrup, K.; Axelstad, M.; Christiansen, S.; Vinggaard, A.; Taxvig, C.; Kortenkamp, A.; Hass, U. Late-life effects on rat reproductive system after developmental exposure to mixtures of endocrine disrupters. Reproduction 2014, 147, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Gaudriault, P.; Mazaud-Guittot, S.; Lavoué, V.; Coiffec, I.; Lesné, L.; Dejucq-Rainsford, N.; Scholze, M.; Kortenkamp, A.; Jégou, B. Endocrine Disruption in Human Fetal Testis Explants by Individual and Combined Exposures to Selected Pharmaceuticals, Pesticides, and Environmental Pollutants. Environ. Health Perspect. 2017, 125, 087004. [Google Scholar] [CrossRef]
- Wan, M.; Co, V.; El-Nezami, H. Endocrine disrupting chemicals and breast cancer: A systematic review of epidemiological studies. Crit. Rev. Food. Sci. Nutr. 2021, 62, 6549–6576. [Google Scholar] [CrossRef]
- Wang, L.; Deng, Q.; Hu, H.; Liu, M.; Gong, Z.; Zhang, S.; Xu-Monette, Z.; Lu, Z.; Young, K.; Ma, X.; et al. Glyphosate induces benign monoclonal gammopathy and promotes multiple myeloma progression in mice. J. Hematol. Oncol. 2019, 12, 70. [Google Scholar] [CrossRef]
- Bradberry, S.M.; Proudfoot, A.T.; Vale, J.A. Poisoning due to chlorophenoxy herbicides. Toxicol Rev. 2004, 23, 65–73. [Google Scholar] [CrossRef]
- Sanmarco, L.M.; Chao, C.C.; Wang, Y.C.; Kenison, J.E.; Li, Z.; Rone, J.M.; Rejano-Gordillo, C.M.; Polonio, C.M.; Gutierrez-Vazquez, C.; Piester, G.; et al. Identification of environmental factors that promote intestinal inflammation. Nature 2022. [Google Scholar] [CrossRef]
- Gupta, P.; Verma, S. Evaluation of genotoxicity induced by herbicide pendimethalin in freshwater fish Clarias batrachus (linn.) and possible role of oxidative stress in induced DNA damage. Drug Chem. Toxicol. 2020, 45, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, W.; Zha, J.; Wang, Z. Dicamba affects sex steroid hormone level and mRNA expression of related genes in adult rare minnow Gobiocypris rarus at environmentally relevant concentrations. Environ. Toxicol. 2015, 30, 693–703. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Ingredient | Formulation | Area Spray in (Hectares) | Active Ingredients Applied (kg) | Declared Co-Formulants |
|---|---|---|---|---|
| Glyphosate | Roundup Probio | 2,812,366 | 2,556,968 | Alkylpolyglycoside, nitroryl, water and minor formulating ingredients |
| Pendimethalin | Stomp Aqua | 309,809 | 295,480 | methylenediphenyl diisocyanate, Isocyanic acid, polymethylenepolyphenylene ester (P-MDI), 4,4’-methylenediphenyl diisocyanate; diphenylmethane-4,4’-diisocyanate, Magnesium sulphate |
| Propyzamide | Kerb flo | 228,341 | 105,406 | Propylene glycol, 2-Naphthalenesulfonic acid, 6-hydroxy-,polymer with formaldehyde and methylphenol, sodium salt |
| Metazachlor | Sultan 50C | 51,656 | 30,958 | - |
| Fluroxypyr | Hurler | 675,157 | 83,041 | BUTANOL-norm, Hydrocarbons, C9, aromatics, Organic sulfonate–proprietary, Solvent naphtha (petroleum), light aromatic |
| Dicamba | Hunter | 76,083 | 39,591 | - |
| Quizalofop-p-ethyl | Targa Super | N/A | N/A | Calcium dodecylbenzene sulphonate, Ethoxylated lauryl alcohol C12, Solvent naphtha (petroleum), super heavy aromatic (<1% naphthalene), solvent naphtha (petroleum), heavy aromatic (<1% naphthalene) |
| 2,4-Dichlorophenoxyacetic acid (2,4-D) | Anti-liserons | N/A | N/A | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferguson, S.; Mesnage, R.; Antoniou, M.N. Cytotoxicity Mechanisms of Eight Major Herbicide Active Ingredients in Comparison to Their Commercial Formulations. Toxics 2022, 10, 711. https://doi.org/10.3390/toxics10110711
Ferguson S, Mesnage R, Antoniou MN. Cytotoxicity Mechanisms of Eight Major Herbicide Active Ingredients in Comparison to Their Commercial Formulations. Toxics. 2022; 10(11):711. https://doi.org/10.3390/toxics10110711
Chicago/Turabian StyleFerguson, Scarlett, Robin Mesnage, and Michael N. Antoniou. 2022. "Cytotoxicity Mechanisms of Eight Major Herbicide Active Ingredients in Comparison to Their Commercial Formulations" Toxics 10, no. 11: 711. https://doi.org/10.3390/toxics10110711
APA StyleFerguson, S., Mesnage, R., & Antoniou, M. N. (2022). Cytotoxicity Mechanisms of Eight Major Herbicide Active Ingredients in Comparison to Their Commercial Formulations. Toxics, 10(11), 711. https://doi.org/10.3390/toxics10110711