
Current and Future Methodology for Quantitation and Site-Specific Mapping the Location of DNA Adducts



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methodology
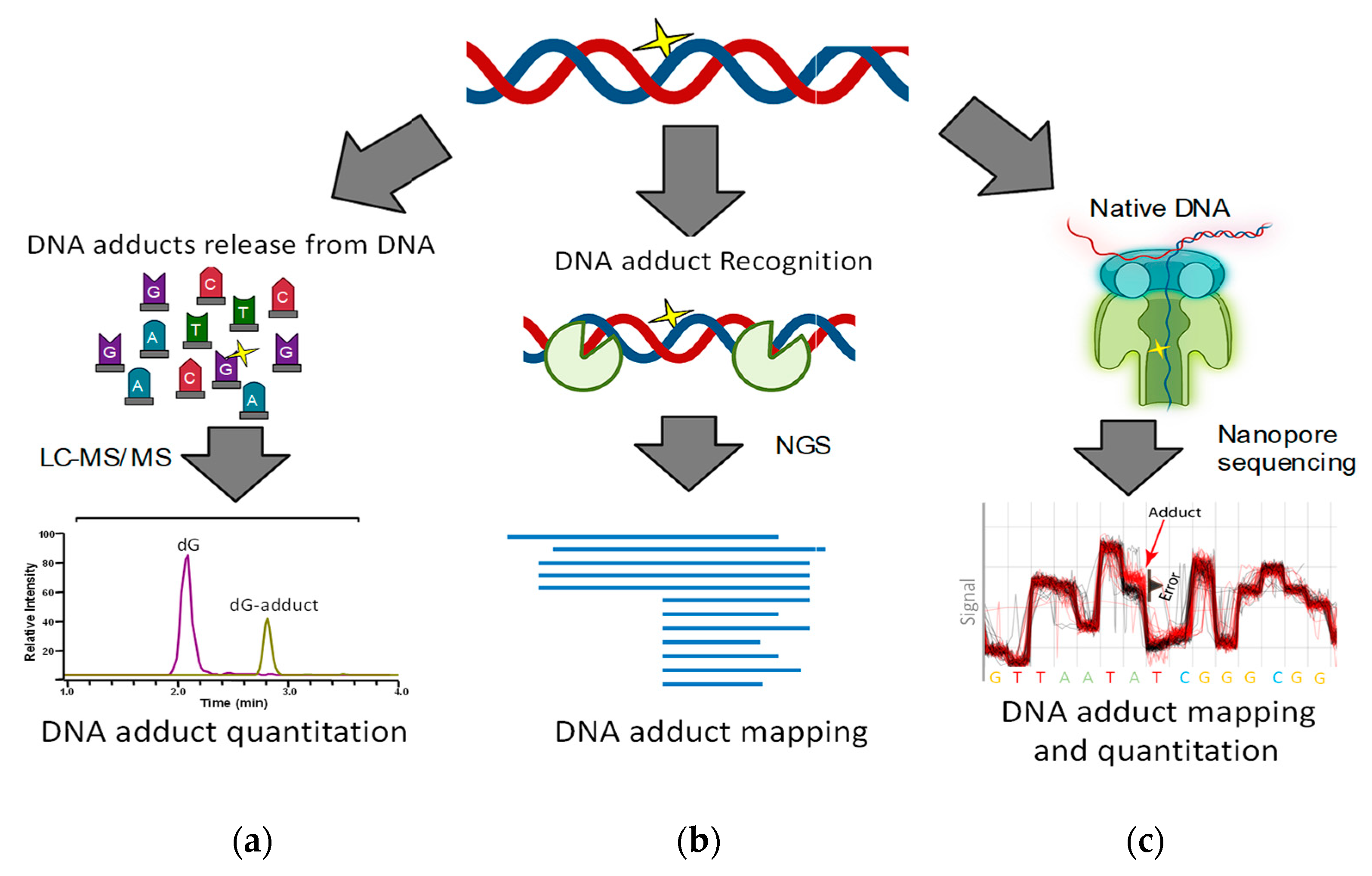
2.1. Mass Spectrometry-Based DNA Adduct Quantitation
2.2. Adductomics, Nontargeted and Qualitative Screening of DNA Adducts
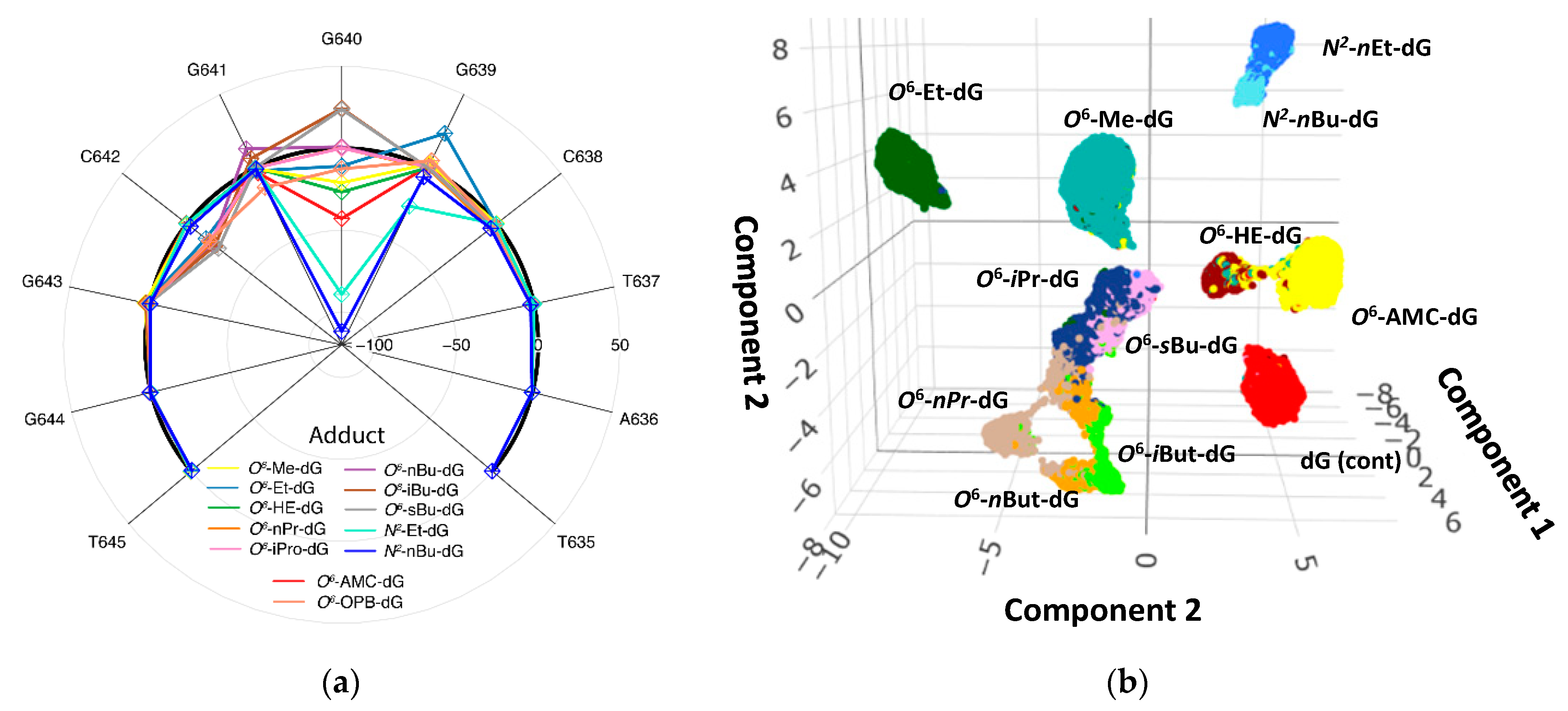
2.3. Amplification-Based Mapping of DNA Adducts
2.4. Single Molecule DNA Sequencing
2.4.1. Nanopore Technology
Oxford Nanopore Technologies (ONT)
PacBio DNA Sequencer
2.4.2. Data Types and Analyses
ONT/ELIGOS
2.4.3. CRISPR/cas-9 Targeted Sequencing
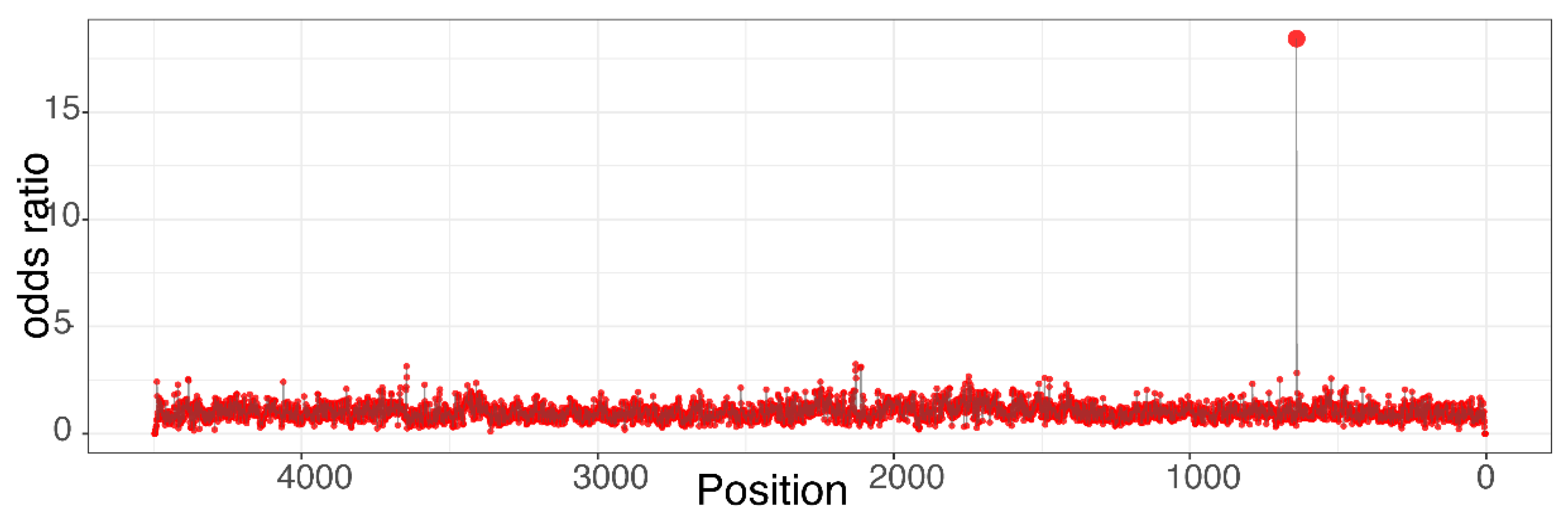
2.4.4. Limit of Detection
- one adduct at a given site/per 105 unmodified bases at position Y (e.g., alkyl-dG/105 dG at position Y);
- five adducts per 109 unmodified nucleotides (e.g., alkyl-dG/109 nnt);
- or approximately one adduct per 109 corresponding nucleotides (e.g., alkyl-dG/109 dG).
3. Discussion
3.1. DNA Adduct Levels
3.2. DNA Adductomics
3.3. DNA Adduct Mapping
3.4. Single Molecule DNA Sequencing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swenberg, J.A.; Lu, K.; Moeller, B.C.; Gao, L.; Upton, P.B.; Nakamura, J.; Starr, T.B. Endogenous versus Exogenous DNA Adducts: Their Role in Carcinogenesis, Epidemiology, and Risk Assessment. Toxicol. Sci. 2011, 120, S130–S145. [Google Scholar] [CrossRef] [PubMed]
- Reiner, B.; Zamenhof, S. Studies on the Chemically Reactive Groups of Deoxyribonucleic Acids. J. Biol. Chem. 1957, 228, 475–486. [Google Scholar] [CrossRef]
- Hwa Yun, B.; Guo, J.; Bellamri, M.; Turesky, R.J. DNA Adducts: Formation, Biological Effects, and New Biospecimens for Mass Spectrometric Measurements in Humans. Mass Spectrom. Rev. 2020, 39, 55–82. [Google Scholar] [CrossRef]
- Tarun, M.; Rusling, J.F. Measuring DNA Nucleobase Adducts Using Neutral Hydrolysis and Liquid Chromatography-Mass Spectrometry. Crit. Rev. Eukaryot. Gene Expr. 2005, 15, 295–316. [Google Scholar] [CrossRef]
- Boysen, G.; Hecht, S.S. Analysis of DNA and Protein Adducts of Benzo[a]Pyrene in Human Tissues Using Structure-Specific Methods. Mutat. Res. 2003, 543, 17–30. [Google Scholar] [CrossRef]
- Boysen, G.; Pachkowski, B.F.; Nakamura, J.; Swenberg, J.A. The Formation and Biological Significance of N7-Guanine Adducts. Mutat. Res. 2009, 678, 76–94. [Google Scholar] [CrossRef] [Green Version]
- Van Zeeland, A.A. Molecular Dosimetry of Chemical Mutagens. Relationship between DNA Adduct Formation and Genetic Changes Analyzed at the Molecular Level. Mutat. Res. 1996, 353, 123–150. [Google Scholar] [CrossRef]
- De Bont, R.; van Larebeke, N. Endogenous DNA Damage in Humans: A Review of Quantitative Data. Mutagenesis 2004, 19, 169–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rundle, A. Carcinogen-DNA Adducts as a Biomarker for Cancer Risk. Mutat. Res. 2006, 600, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, H. DNA Adducts in Human Carcinogenesis: Etiological Relevance and Structure-Activity Relationship. Mutat. Res. 1996, 340, 67–79. [Google Scholar] [CrossRef]
- Harris, C.C. Chemical and Physical Carcinogenesis: Advances and Perspectives for the 1990s. Cancer Res. 1991, 51, 5023s–5044s. [Google Scholar]
- Harvey, R.G. Historical Overview of Chemical Carcinogenesis. In Chemical Carcinogenesis; Penning, T.M., Ed.; Humana Press: Totowa, NJ, USA, 2011; Volume 6, pp. 1–26. ISBN 9781617379949. [Google Scholar]
- Hartwig, A.; Arand, M.; Epe, B.; Guth, S.; Jahnke, G.; Lampen, A.; Martus, H.-J.; Monien, B.; Rietjens, I.M.C.M.; Schmitz-Spanke, S.; et al. Mode of Action-Based Risk Assessment of Genotoxic Carcinogens. Arch. Toxicol. 2020, 94, 1787–1877. [Google Scholar] [CrossRef] [PubMed]
- La, D.K.; Swenberg, J.A. DNA Adducts: Biological Markers of Exposure and Potential Applications to Risk Assessment. Mutat. Res. 1996, 365, 129–146. [Google Scholar] [CrossRef]
- Phillips, D.H. DNA Adducts as Markers of Exposure and Risk. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2005, 577, 284–292. [Google Scholar] [CrossRef]
- Poirier, M.C. DNA Adducts as Exposure Biomarkers and Indicators of Cancer Risk. Environ. Health Perspect. 1997, 105 (Suppl. 4), 907–912. [Google Scholar]
- Pottenger, L.H.; Boysen, G.; Brown, K.; Cadet, J.; Fuchs, R.P.; Johnson, G.E.; Swenberg, J.A. Understanding the Importance of Low-Molecular Weight (Ethylene Oxide- and Propylene Oxide-Induced) DNA Adducts and Mutations in Risk Assessment: Insights from 15 Years of Research and Collaborative Discussions. Environ. Mol. Mutagenesis 2019, 60, 100–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balbo, S.; Turesky, R.J.; Villalta, P.W. DNA Adductomics. Chem. Res. Toxicol. 2014, 27, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, S.J.; Guo, J.; Wang, J.; Villalta, P.W.; Turesky, R.J. Methods and Challenges for Computational Data Analysis for DNA Adductomics. Chem. Res. Toxicol. 2019, 32, 2156–2168. [Google Scholar] [CrossRef]
- Guo, J.; Turesky, R.J. Emerging Technologies in Mass Spectrometry-Based DNA Adductomics. High-Throughput 2019, 8, 13. Available online: https://www.mdpi.com/2571-5135/8/2/13 (accessed on 13 December 2021). [CrossRef] [Green Version]
- Alhegaili, A.S.; Ji, Y.; Sylvius, N.; Blades, M.J.; Karbaschi, M.; Tempest, H.G.; Jones, G.D.D.; Cooke, M.S. Genome-Wide Adductomics Analysis Reveals Heterogeneity in the Induction and Loss of Cyclobutane Thymine Dimers across Both the Nuclear and Mitochondrial Genomes. Int. J. Mol. Sci. 2019, 20, 5112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, V.; Khan, A.; Gonzales, C.; Phillips, D.H.; Schoket, B.; Györffy, E.; Anna, L.; Kovács, K.; Møller, P.; Loft, S.; et al. Validation of Biomarkers for the Study of Environmental Carcinogens: A Review. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2008, 13, 505–534. [Google Scholar] [CrossRef] [PubMed]
- Santella, R.M. Immunological Methods for Detection of Carcinogen-DNA Damage in Humans. Cancer Epidemiol. Biomark. Prev. 1999, 8, 733–739. [Google Scholar]
- Phillips, D.H.; Arlt, V.M. The 32p-Postlabeling Assay for DNA Adducts. Nat. Protoc. 2007, 2, 2772–2781. [Google Scholar] [CrossRef]
- Cummings, J.; French, R.C.; Smyth, J.F. Application of High-Performance Liquid Chromatography for Recognition of Covalent Nucleic Acid Modification with Anticancer Drugs. J. Chromatogr. 1993, 618, 251–276. [Google Scholar] [CrossRef]
- Koc, H.; Swenberg, J.A. Applications of Mass Spectrometry for Quantitation of DNA Adducts. J. Chromatogr. B 2002, 778, 323–343. [Google Scholar] [CrossRef]
- Farmer, P.B.; Singh, R. Use of DNA Adducts to Identify Human Health Risk from Exposure to Hazardous Environmental Pollutants: The Increasing Role of Mass Spectrometry in Assessing Biologically Effective Doses of Genotoxic Carcinogens. Mutat. Res. 2008, 659, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Dole, M.; Mack, L.L.; Hines, R.L.; Mobley, R.C.; Ferguson, L.D.; Alice, M.B. Molecular Beams of Macroions. J. Chem. Phys. 1968, 49, 2240–2249. [Google Scholar] [CrossRef]
- Fenn, J.B.; Mann, M.; Meng, C.K.; Wong, S.F.; Whitehouse, C.M. Electrospray Ionization-Principles and Practice. Mass Spectrom. Rev. 1990, 9, 37–70. [Google Scholar] [CrossRef]
- Guo, J.; Villalta, P.W.; Weight, C.J.; Bonala, R.; Johnson, F.; Rosenquist, T.A.; Turesky, R.J. Targeted and Untargeted Detection of DNA Adducts of Aromatic Amine Carcinogens in Human Bladder by Ultra-Performance Liquid Chromatography-High-Resolution Mass Spectrometry. Chem. Res. Toxicol. 2018, 31, 1382–1397. [Google Scholar] [CrossRef]
- Guo, J.; Turesky, R.J.; Tarifa, A.; DeCaprio, A.P.; Cooke, M.S.; Walmsley, S.J.; Villalta, P.W. Development of a DNA Adductome Mass Spectral Database. Chem. Res. Toxicol. 2020, 33, 852–854. [Google Scholar] [CrossRef]
- Behl, T.; Rachamalla, M.; Najda, A.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Al-harrasi, A.; Chigurupati, S.; Vargas-de-la-cruz, C.; et al. Applications of Adductomics in Chemically Induced Adverse Outcomes and Major Emphasis on DNA Adductomics: A Pathbreaking Tool in Biomedical Research. Int. J. Mol. Sci. 2021, 22, 141. [Google Scholar] [CrossRef] [PubMed]
- Olinski, R.; Rozalski, R.; Gackowski, D. Mass Spectrometry-Based Analysis of DNA Modifications: Potential Applications in Basic Research and Clinic. Methods Mol. Biol. 2021, 2198, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Stepanov, I.; Hecht, S.S. Recent Studies on DNA Adducts Resulting from Human Exposure to Tobacco Smoke. Toxics 2019, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Monien, B.H. Mass Spectrometric DNA Adduct Quantification by Multiple Reaction Monitoring and Its Future Use for the Molecular Epidemiology of Cancer. Adv. Exp. Med. Biol. 2019, 1140, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Tretyakova, N.; Goggin, M.; Sangaraju, D.; Janis, G. Quantitation of DNA Adducts by Stable Isotope Dilution Mass Spectrometry. Chem. Res. Toxicol. 2012, 25, 2007–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-J.; Cooke, M.S.; Hu, C.-W.; Chao, M.-R. Novel Approach to Integrated DNA Adductomics for the Assessment of in Vitro and in Vivo Environmental Exposures. Arch. Toxicol. 2018, 92, 2665–2680. [Google Scholar] [CrossRef]
- Chang, Y.J.; Cooke, M.S.; Chen, Y.R.; Yang, S.F.; Li, P.S.; Hu, C.W.; Chao, M.R. Is High Resolution a Strict Requirement for Mass Spectrometry-Based Cellular DNA Adductomics? Chemosphere 2021, 274, 129991. [Google Scholar] [CrossRef]
- Zhang, P.; Carlsten, C.; Chaleckis, R.; Hanhineva, K.; Huang, M.; Isobe, T.; Koistinen, V.M.; Meister, I.; Papazian, S.; Sdougkou, K.; et al. Defining the Scope of Exposome Studies and Research Needs from a Multidisciplinary Perspective. Environ. Sci. Technol. Lett. 2021, 8, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.J.; Carlson, E.S.; Stornetta, A.; Balskus, E.P.; Villalta, P.W.; Balbo, S. Extension of Diagnostic Fragmentation Filtering for Automated Discovery in DNA Adductomics. Anal. Chem. 2021, 93, 5754–5762. [Google Scholar] [CrossRef] [PubMed]
- Htun, H.; Johnston, B.H. Mapping Adducts of DNA Structural Probes Using Transcription and Primer Extension Approaches. Methods Enzymol. 1992, 212, 272–294. [Google Scholar] [CrossRef]
- Sinden, R.R.; Ussery, D.W. Analysis of DNA Structure In Vivo Using Psoralen Photobinding: Measurement of Supercoiling, Topological Domains, and DNA-Protein Interactions. Methods Enzymol. 1992, 212, 319–335. [Google Scholar] [CrossRef]
- Cao, B.; Wu, X.; Zhou, J.; Wu, H.; Liu, L.; Zhang, Q.; Demott, M.S.; Gu, C.; Wang, L.; You, D.; et al. Nick-Seq for Single-Nucleotide Resolution Genomic Maps of DNA Modifications and Damage. Nucleic Acids Res. 2020, 48, 6715–6725. [Google Scholar] [CrossRef] [PubMed]
- Denissenko, M.F.; Venkatachalam, S.; Ma, Y.H.; Wani, A.A. Site-Specific Induction and Repair of Benzo[a]Pyrene Diol Epoxide DNA Damage in Human H-Ras Protooncogene as Revealed by Restriction Cleavage Inhibition. Mutat. Res. 1996, 363, 27–42. [Google Scholar] [CrossRef]
- Denissenko, M.F.; Pao, A.; Tang, M.-s.; Pfeifer, G.P. Preferential Formation of Benzo[a]Pyrene Adducts at Lung Cancer Mutational Hotspots in P53. Science 1996, 274, 430–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denissenko, M.F.; Chen, J.X.; Tang, M.S.; Pfeifer, G.P. Cytosine Methylation Determines Hot Spots of DNA Damage in the Human P53 Gene. Proc. Natl. Acad. Sci. USA 1997, 94, 3893–3898. [Google Scholar] [CrossRef] [Green Version]
- Mao, P.; Smerdon, M.J.; Roberts, S.A.; Wyrick, J.J. Chromosomal Landscape of UV Damage Formation and Repair at Single-Nucleotide Resolution. Proc. Natl. Acad. Sci. USA 2016, 113, 9057–9062. [Google Scholar] [CrossRef] [Green Version]
- Mao, P.; Brown, A.J.; Malc, E.P.; Mieczkowski, P.A.; Smerdon, M.J.; Roberts, S.A.; Wyrick, J.J. Genome-Wide Maps of Alkylation Damage, Repair, and Mutagenesis in Yeast Reveal Mechanisms of Mutational Heterogeneity. Genome Res. 2017, 27, 1674–1684. [Google Scholar] [CrossRef]
- Hu, J.; Li, W.; Adebali, O.; Yang, Y.; Oztas, O.; Selby, C.P.; Sancar, A. Genome-Wide Mapping of Nucleotide Excision Repair with XR-Seq. Nat. Protoc. 2019, 14, 248–282. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hu, J.; Adebali, O.; Adar, S.; Yang, Y.; Chiou, Y.-Y.; Sancar, A. Human Genome-Wide Repair Map of DNA Damage Caused by the Cigarette Smoke Carcinogen Benzo[a]Pyrene. Proc. Natl. Acad. Sci. USA 2017, 114, 6752–6757. [Google Scholar] [CrossRef] [Green Version]
- Govindan, R.; Szczesna, A.; Ahn, M.; Schneider, C.; Gonzalez Mella, P.F.; Barlesi, F.; Han, B.; Ganea, D.E.; Von Pawel, J.; Vladimirov, V.; et al. Phase III Trial of Ipilimumab Combined with Paclitaxel and Carboplatin in Advanced Squamous Non-Small-Cell Lung Cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2017, 35, 3449–3457. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, J.; Selby, C.P.; Li, W.; Yimit, A.; Jiang, Y.; Sancar, A. Single-Nucleotide Resolution Analysis of Nucleotide Excision Repair of Ribosomal DNA in Humans and Mice. J. Biol. Chem. 2019, 294, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Xiong, X.; Song, J.; He, C.; Yi, C. Base-Resolution Analysis of Cisplatin-DNA Adducts at the Genome Scale. Angew. Chem. Int. Ed. Engl. 2016, 55, 14246–14249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; McKeague, M.; Sturla, S.J. Nucleotide-Resolution Genome-Wide Mapping of Oxidative DNA Damage by Click-Code-Seq. J. Am. Chem. Soc. 2018, 140, 9783–9787. [Google Scholar] [CrossRef] [PubMed]
- Schutsky, E.K.; Denizio, J.E.; Hu, P.; Liu, M.Y.; Nabel, C.S.; Fabyanic, E.B.; Hwang, Y.; Bushman, F.D.; Wu, H.; Kohli, R.M. Nondestructive, Base-Resolution Sequencing of 5-Hydroxymethylcytosine Using a DNA Deaminase. Nat. Biotechnol. 2018, 36, 1083–1090. [Google Scholar] [CrossRef]
- Vitelli, V.; Galbiati, A.; Iannelli, F.; Pessina, F.; Sharma, S.; d’Adda di Fagagna, F. Recent Advancements in DNA Damage-Transcription Crosstalk and High-Resolution Mapping of DNA Breaks. Annu. Rev. Genom. Hum. Genet. 2017, 18, 87–113. [Google Scholar] [CrossRef] [PubMed]
- Amente, S.; Scala, G.; Majello, B.; Azmoun, S.; Tempest, H.G.; Premi, S.; Cooke, M.S. Genome-Wide Mapping of Genomic DNA Damage: Methods and Implications. Cell. Mol. Life Sci. 2021, 78, 6745–6762. [Google Scholar] [CrossRef] [PubMed]
- Poetsch, A.R. The Genomics of Oxidative DNA Damage, Repair, and Resulting Mutagenesis. Comput. Struct. Biotechnol. J. 2020, 18, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Nookaew, I.; Jenjaroenpun, P.; Du, H.; Wang, P.; Wu, J.; Wongsurawat, T.; Moon, S.H.; Huang, E.; Wang, Y.; Boysen, G. Detection and Discrimination of DNA Adducts Differing in Size, Regiochemistry, and Functional Group by Nanopore Sequencing. Chem. Res. Toxicol. 2020, 33, 2944–2952. [Google Scholar] [CrossRef]
- Perera, R.T.; Fleming, A.M.; Johnson, R.P.; Burrows, C.J.; White, H.S. Detection of Benzo[a]Pyrene-Guanine Adducts in Single-Stranded DNA Using the α-Hemolysin Nanopore. Nanotechnology 2015, 26, 074002. [Google Scholar] [CrossRef] [Green Version]
- An, N.; Fleming, A.M.; White, H.S.; Burrows, C.J. Crown Ether-Electrolyte Interactions Permit Nanopore Detection of Individual DNA Abasic Sites in Single Molecules. Proc. Natl. Acad. Sci. USA 2012, 109, 11504–11509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, N.; Fleming, A.M.; Rosecrans, N.C.; Liao, Y.; Burrows, C.J. Synthesis of Site-Specific Crown Ether Adducts to DNA Abasic Sites: 8-Oxo-7,8-Dihydro-2’-Deoxyguanosine and 2’-Deoxycytidine. Methods Mol. Biol. 2019, 1973, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.J.; Martínez Cuesta, S.; van Delft, P.; Balasubramanian, S. Sequencing Abasic Sites in DNA at Single-Nucleotide Resolution. Nat. Chem. 2019, 11, 629–637. [Google Scholar] [CrossRef]
- Tan, C.S.; Fleming, A.M.; Ren, H.; Burrows, C.J.; White, H.S. γ-Hemolysin Nanopore Is Sensitive to Guanine-to-Inosine Substitutions in Double-Stranded DNA at the Single-Molecule Level. J. Am. Chem. Soc. 2018, 140, 14224–14234. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Fleming, A.M.; Ding, Y.; Ren, H.; White, H.S.; Burrows, C.J. Nanopore Analysis of the 5-Guanidinohydantoin to Iminoallantoin Isomerization in Duplex DNA. J. Org. Chem. 2018, 83, 3973–3978. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.M.; Ding, Y.; Burrows, C.J. Oxidative DNA Damage Is Epigenetic by Regulating Gene Transcription via Base Excision Repair. Proc. Natl. Acad. Sci. USA 2017, 114, 2604–2609. [Google Scholar] [CrossRef]
- Ding, Y.; Fleming, A.M.; Burrows, C.J. Sequencing the Mouse Genome for the Oxidatively Modified Base 8-Oxo-7,8-Dihydroguanine by OG-Seq. J. Am. Chem. Soc. 2017, 139, 2569–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, A.; Holmes, N.; Rakyan, V.; Loose, M. BulkVis: A Graphical Viewer for Oxford Nanopore Bulk FAST5 Files. Bioinformatics 2019, 35, 2193–2198. [Google Scholar] [CrossRef]
- Tyson, J.R.; O’Neil, N.J.; Jain, M.; Olsen, H.E.; Hieter, P.; Snutch, T.P. MinION-Based Long-Read Sequencing and Assembly Extends the Caenorhabditis Elegans Reference Genome. Genome Res. 2018, 28, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Seki, M. Recent Advances in the Detection of Base Modifications Using the Nanopore Sequencer. J. Hum. Genet. 2020, 65, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Rand, A.C.; Jain, M.; Eizenga, J.M.; Musselman-Brown, A.; Olsen, H.E.; Akeson, M.; Paten, B. Mapping DNA Methylation with High-Throughput Nanopore Sequencing. Nat. Methods 2017, 14, 411–413. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.T.; Workman, R.E.; Zuzarte, P.C.; David, M.; Dursi, L.J.; Timp, W. Detecting DNA Cytosine Methylation Using Nanopore Sequencing. Nat. Methods 2017, 14, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fang, L.; Yu, G.; Wang, D.; Xiao, C.-L.; Wang, K. Detection of DNA Base Modifications by Deep Recurrent Neural Network on Oxford Nanopore Sequencing Data. Nat. Commun. 2019, 10, 2449. [Google Scholar] [CrossRef] [Green Version]
- Ni, P.; Huang, N.; Zhang, Z.; Wang, D.-P.; Liang, F.; Miao, Y.; Xiao, C.-L.; Luo, F.; Wang, J. DeepSignal: Detecting DNA Methylation State from Nanopore Sequencing Reads Using Deep-Learning. Bioinformatics 2019, 35, 4586–4595. [Google Scholar] [CrossRef] [PubMed]
- Flusberg, B.A.; Webster, D.R.; Lee, J.H.; Travers, K.J.; Olivares, E.C.; Clark, T.A.; Korlach, J.; Turner, S.W. Direct Detection of DNA Methylation during Single-Molecule, Real-Time Sequencing. Nat. Methods 2010, 7, 461–465. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Chen, C.; DeMott, M.S.; Cheng, Q.; Clark, T.A.; Xiong, X.; Zheng, X.; Butty, V.; Levine, S.S.; Yuan, G.; et al. Genomic Mapping of Phosphorothioates Reveals Partial Modification of Short Consensus Sequences. Nat. Commun. 2014, 5, 3951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlgren, N.A.; Chen, Y.; Needham, D.M.; Parada, A.E.; Sachdeva, R.; Trinh, V.; Chen, T.; Fuhrman, J.A. Genome and Epigenome of a Novel Marine Thaumarchaeota Strain Suggest Viral Infection, Phosphorothioation DNA Modification and Multiple Restriction Systems. Environ. Microbiol. 2017, 19, 2434–2452. [Google Scholar] [CrossRef]
- Jenjaroenpun, P.; Wongsurawat, T.; Pereira, R.; Patumcharoenpol, P.; Ussery, D.W.; Nielsen, J.; Nookaew, I. Complete Genomic and Transcriptional Landscape Analysis Using Third-Generation Sequencing: A Case Study of Saccharomyces Cerevisiae CEN.PK113-7D. Nucleic Acids Res. 2018, 46, e38. [Google Scholar] [CrossRef] [Green Version]
- Wongsurawat, T.; Jenjaroenpun, P.; Taylor, M.K.; Lee, J.; Tolardo, A.L.; Parvathareddy, J.; Kandel, S.; Wadley, T.D.; Kaewnapan, B.; Athipanyasilp, N.; et al. Rapid Sequencing of Multiple RNA Viruses in Their Native Form. Front. Microbiol. 2019, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Holt, K.E. Performance of Neural Network Basecalling Tools for Oxford Nanopore Sequencing. Genome Biol. 2019, 20, 129. [Google Scholar] [CrossRef] [Green Version]
- Gilpatrick, T.; Lee, I.; Graham, J.E.; Raimondeau, E.; Bowen, R.; Heron, A.; Downs, B.; Sukumar, S.; Sedlazeck, F.J.; Timp, W. Targeted Nanopore Sequencing with Cas9-Guided Adapter Ligation. Nat. Biotechnol. 2020, 38, 433–438. [Google Scholar] [CrossRef]
- Veres, A.; Gosis, B.S.; Ding, Q.; Collins, R.; Ragavendran, A.; Brand, H.; Erdin, S.; Cowan, C.A.; Talkowski, M.E.; Musunuru, K. Low Incidence of Off-Target Mutations in Individual CRISPR-Cas9 and TALEN Targeted Human Stem Cell Clones Detected by Whole-Genome Sequencing. Cell Stem Cell 2014, 15, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Wongsurawat, T.; Jenjaroenpun, P.; de Loose, A.; Alkam, D.; Ussery, D.W.; Nookaew, I.; Leung, Y.K.; Ho, S.M.; Day, J.D.; Rodriguez, A. A Novel Cas9-Targeted Long-Read Assay for Simultaneous Detection of IDH1/2 Mutations and Clinically Relevant MGMT Methylation in Fresh Biopsies of Diffuse Glioma. Acta Neuropathol. Commun. 2020, 8, 87. [Google Scholar] [CrossRef]
- Villalta, P.W.; Hochalter, J.B.; Hecht, S.S. Ultrasensitive High-Resolution Mass Spectrometric Analysis of a DNA Adduct of the Carcinogen Benzo[a]Pyrene in Human Lung. Anal. Chem. 2017, 89, 12735–12742. [Google Scholar] [CrossRef]
- Walker, V.E.; Fennel, T.R.; Upton, P.B.; Skopek, T.R.; Prevost, V.; Shuker, D.E.G.; Swenberg, J.A. Molecular Dosimetry of Ethylene Oxide: Formation and Persistence of 7-(2-Hydroxyethyl)Guanine in DNA Following Repeated Exposures of Rats and Mice. Cancer Res. 1992, 52, 4328–4334. [Google Scholar]
- Walker, V.E.; Fennell, T.R.; Boucheron, J.A.; Fedtke, N.; Ciroussel, F.; Swenberg, J.A. Macromolecular Adducts of Ethylene Oxide: A Literature Review and a Time-Course Study on the Formation of 7-(2-Hydroxyethyl)Guanine Following Exposures of Rats by Inhalation. Mutat. Res. 1990, 233, 151–164. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Persmark, M. Mechanism of Formation of Ethenoguanine Adducts from 2-Haloacetaldehydes: 13C-Labeling Patterns with 2-Bromoacetaldehyde. Chem. Res. Toxicol. 1994, 7, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.; Barbin, A.; Bartsch, H. Etheno DNA-Base Adducts from Endogenous Reactive Species. Mutat. Res.-Fundam. Mol. Mech. Mutagenesis 1999, 424, 59–69. [Google Scholar] [CrossRef]
- Collins, A.R.; Cadet, J.; Moller, L.; Poulsen, H.E.; Viña, J. Are We Sure We Know How to Measure 8-Oxo-7,8-Dihydroguanine in DNA from Human Cells? Arch. Biochem. Biophys. 2004, 423, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.; Gedik, C.; Vaughan, N.; Wood, S.; White, A.; Dubois, J.; Rees, J.F.; Loft, S.; Møller, P.; Poulsen, H.; et al. Measurement of DNA Oxidation in Human Cells by Chromatographic and Enzymic Methods. Free Radic. Biol. Med. 2003, 34, 1089–1099. [Google Scholar] [CrossRef]
- Lunec, J. ESCODD: European Standards Committee on Oxidative DNA Damage. Free Radic. Res. 1998, 29, 601–608. [Google Scholar] [CrossRef]
- Collins, A. Comparison of Different Methods of Measuring 8-Oxoguanine as a Marker of Oxidative DNA Damage. Free Radic. Res. 2000, 32, 333–341. [Google Scholar] [CrossRef]
- ESCODD. Comparative Analysis of Baseline 8-Oxo-7,8-Dihydroguanine in Mammalian Cell DNA, by Different Methods in Different Laboratories: An Approach to Consensus. Carcinogenesis 2002, 23, 2129–2133. [Google Scholar] [CrossRef]
- Boysen, G.; Collins, L.B.; Liao, S.; Luke, A.M.; Pachkowski, B.F.; Watters, J.L.; Swenberg, J.A. Analysis of 8-Oxo-7,8-Dihydro-2’-Deoxyguanosine by Ultra High Pressure Liquid Chromatography-Heat Assisted Electrospray Ionization-Tandem Mass Spectrometry. J. Chromatography. B Anal. Technol. Biomed. Life Sci. 2010, 878, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Watters, J.J.L.; Satia, J.J.A.; da Costa, K.-A.; Boysen, G.; Collins, L.B.; Morrow, J.D.; Milne, G.L.; Swenberg, J.A.; Costa, K. Comparison of Three Oxidative Stress Biomarkers in a Sample of Healthy Adults. Biomarkers 2009, 14, 587–595. [Google Scholar] [CrossRef]
- Ma, B.; Zarth, A.T.; Carlson, E.S.; Villalta, P.W.; Upadhyaya, P.; Stepanov, I.; Hecht, S.S. Identification of More than 100 Structurally Unique DNA-Phosphate Adducts Formed during Rat Lung Carcinogenesis by the Tobacco-Specific Nitrosamine 4-(Methylnitrosamino)-1-(3-Pyridyl)-1-Butanone. Carcinogenesis 2018, 39, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanaly, R.A.; Matsui, S.; Hanaoka, T.; Matsuda, T. Application of the Adductome Approach to Assess Intertissue DNA Damage Variations in Human Lung and Esophagus. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2007, 625, 83–93. [Google Scholar] [CrossRef]
- Bessette, E.E.; Goodenough, A.K.; Langouët, S.; Yasa, I.; Kozekov, I.D.; Spivack, S.D.; Turesky, R.J. Screening for DNA Adducts by Data-Dependent Constant Neutral Loss-Triple Stage Mass Spectrometry with a Linear Quadrupole Ion Trap Mass Spectrometer. Anal. Chem. 2009, 81, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Bessette, E.E.; Spivack, S.D.; Goodenough, A.K.; Wang, T.; Pinto, S.; Kadlubar, F.F.; Turesky, R.J. Identification of Carcinogen DNA Adducts in Human Saliva by Linear Quadrupole Ion Trap/Multistage Tandem Mass Spectrometry. Chem. Res. Toxicol. 2010, 23, 1234–1244. [Google Scholar] [CrossRef] [Green Version]
- Iwashita, Y.; Ohnishi, I.; Matsushita, Y.; Ohtsuka, S.; Yamashita, T.; Inaba, K.; Fukazawa, A.; Ochiai, H.; Matsumoto, K.; Kurono, N.; et al. Geospatial Assessments of DNA Adducts in the Human Stomach: A Model of Field Cancerization. Cancers 2021, 13, 3728. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, I.; Iwashita, Y.; Matsushita, Y.; Ohtsuka, S.; Yamashita, T.; Inaba, K.; Fukazawa, A.; Ochiai, H.; Matsumoto, K.; Kurono, N.; et al. Mass Spectrometric Profiling of DNA Adducts in the Human Stomach Associated with Damage from Environmental Factors. Genes Environ. Off. J. Jpn. Environ. Mutagen Soc. 2021, 43, 12. [Google Scholar] [CrossRef]
- Tang, M.S.; Zheng, J.B.; Denissenko, M.F.; Pfeifer, G.P.; Zheng, Y. Use of UvrABC Nuclease to Quantify Benzo[a]Pyrene Diol Epoxide-DNA Adduct Formation at Methylated versus Unmethylated CpG Sites in the P53 Gene. Carcinogenesis 1999, 20, 1085–1089. [Google Scholar] [CrossRef]
- Dittrich, K.A.; Krugh, T.R. Mapping of (+/−)-Anti-Benzo[a]Pyrene Diol Epoxide Adducts to Human c-Ha-Ras1 Protooncogene. Chem. Res. Toxicol. 1991, 4, 277–281. [Google Scholar] [CrossRef]
- Briedé, J.J.; Godschalk, R.W.L.; Emans, M.T.G.; de Kok, T.M.C.M.; van Agen, E.; van Maanen, J.; van Schooten, F.-J.; Kleinjans, J.C.S. In Vitro and in Vivo Studies on Oxygen Free Radical and DNA Adduct Formation in Rat Lung and Liver during Benzo[a]Pyrene Metabolism. Free. Radic. Res. 2004, 38, 995–1002. [Google Scholar] [CrossRef]
- Matter, B.; Wang, G.; Jones, R.; Tretyakova, N. Formation of Diastereomeric Benzo[a]Pyrene Diol Epoxide-Guanine Adducts in P53 Gene-Derived DNA Sequences. Chem. Res. Toxicol. 2004, 17, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Bush, J.; Maulbetsch, W.; Lepoitevin, M.; Wiener, B.; Mihovilovic Skanata, M.; Moon, W.; Pruitt, C.; Stein, D. The Nanopore Mass Spectrometer. Rev. Sci. Instrum. 2017, 88. [Google Scholar] [CrossRef]
- Maulbetsch, W.; Wiener, B.; Poole, W.; Bush, J.; Stein, D. Preserving the Sequence of a Biopolymer’s Monomers as They Enter an Electrospray Mass Spectrometer. Phys. Rev. Appl. 2016, 6, 054006. [Google Scholar] [CrossRef] [Green Version]
- Brodbelt, J.S. Photodissociation Mass Spectrometry: New Tools for Characterization of Biological Molecules. Chem. Soc. Rev. 2014, 43, 2757–2783. [Google Scholar] [CrossRef]
- Sun, X.; Song, L.; Yang, W.; Zhang, L.; Liu, M.; Li, X.; Tian, G.; Wang, W. Nanopore Sequencing and Its Clinical Applications. Methods Mol. Biol. 2020, 2204, 13–32. [Google Scholar] [CrossRef]
- Lin, B.; Hui, J.; Mao, H. Nanopore Technology and Its Applications in Gene Sequencing. Biosensors 2021, 11, 214. [Google Scholar] [CrossRef]
- Robertson, J.W.F.; Ghimire, M.L.; Reiner, J.E. Nanopore Sensing: A Physical-Chemical Approach. Biochim. Biophys. Acta-Biomembr. 2021, 1863, 183644. [Google Scholar] [CrossRef]
- Totsuka, Y.; Watanabe, M.; Lin, Y. New Horizons of DNA Adductome for Exploring Environmental Causes of Cancer. Cancer Sci. 2020, 112, 7–15. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boysen, G.; Nookaew, I. Current and Future Methodology for Quantitation and Site-Specific Mapping the Location of DNA Adducts. Toxics 2022, 10, 45. https://doi.org/10.3390/toxics10020045
Boysen G, Nookaew I. Current and Future Methodology for Quantitation and Site-Specific Mapping the Location of DNA Adducts. Toxics. 2022; 10(2):45. https://doi.org/10.3390/toxics10020045
Chicago/Turabian StyleBoysen, Gunnar, and Intawat Nookaew. 2022. "Current and Future Methodology for Quantitation and Site-Specific Mapping the Location of DNA Adducts" Toxics 10, no. 2: 45. https://doi.org/10.3390/toxics10020045
APA StyleBoysen, G., & Nookaew, I. (2022). Current and Future Methodology for Quantitation and Site-Specific Mapping the Location of DNA Adducts. Toxics, 10(2), 45. https://doi.org/10.3390/toxics10020045